Thermus thermophilus HJ6 유래 N-말단 결실 DNA Polymerase의 정제 및 특성
전숭종1,2,3*·서민호1
1동의대학교 생명공학과, 2동의대학교 바이오물질제어학과, 3동의대학교 블루바이오 RIC
Purification and Characterization of the N-terminally Truncated DNA Polymerase from Thermus ther- mophilus HJ6. Jeon, Sung-Jong1,2,3* and Min-Ho Seo1. 1Department of Biotechnology & Bioengineering, Dong-Eui University, Busan 614-714, Korea, 2Department of Biomaterial Control, Dong-Eui University, Busan 614-714, Korea, 3Blue-Bio Industry RIC, Dong-Eui University, Busan 614-714, Korea − The gene encoding N- terminally truncated Tod polymerase (∆Tod polymerase) from Thermus thermophilus HJ6 was expressed in Escherichia coli under the control of the lambda pR and pL tandem promoters on the expression vector pJLA503. The N-terminal domain (250 amino acids) of Tod polymerase was removed without significant effect on enzyme activity and stability, while no 5'→3' exonuclease activity was detected. The ∆Tod poly- merase was verified to possess very efficient reverse transcriptase (RT) activity in the presence of MgCl2. The cDNA can also be amplified in the polymerase chain reaction (PCR) with this mutant enzyme. The ∆Tod poly- merase was exhibited higher activity than the Taq polymerase in a one-step RT-PCR.
Key words: DNA polymerase, RT-PCR, Thermus thermophilus, PCR, exonuclease
서 론
RNA 분자의 분석과 검출에 대한 표준 방법에는 in situ hybridization, northern, dot, slot blot analysis, S1 nuclease analysis 및 RNase protection assay 등이 있다. 그러나 이들 방법은 많은 양의 RNA를 필요로 하거나 분석 방법상에 어 려움이 있기 때문에 활용 면에서 제한을 받을 수 있다. 이들 문제의 해결방안으로 DNA를 증폭하는 PCR 기술은 avian myeloblastosis virus reverse transcriptase(RTase) 또는 moloney murine leukemia virus(M-MLV) RTase과 같은 역 전사효소의 등장과 더불어 RNA를 주형으로 DNA를 증폭하 는 Reverse Transcription-Polymerase Chain Reaction (RT- PCR) 기술로 발전하였다[4, 7]. RT-PCR 기술은 유전자 발 현의 검출[7], 증폭된 RNA 서열 분석[13], 유전병과 감염원 의 진단[5] 등에 활용되고 있다.
역전사 반응에서는 안정한 2차 RNA 구조에서의 cDNA 합성과 primer 신장의 특이성을 향상시키기 위해 고온에서 의 RT 반응이 요구되었고 이런 이유로 내열성 역전사 효소 의 필요성이 인식 되었다[1]. PCR의 출현 후 Taq poly- merase의 역전사 활성을 이용하여 mRNA로부터 한번에 PCR까지 수행하는 방법이 보고되었다[12]. 그러나 증폭에는 다량(1~5 µg)의 RNA가 필요하기 때문에 실용적이지 않았
다. 그 후 Tth polymerase과 같은 강한 역전사 활성이 있는 효소가 발견되어 one-step RT-PCR이 실용화되었다[9]. Tth polymerase는 금속이온 Mn2+가 존재하면 강한 역전사활성 을 나타내기 때문에 MnCl2존재 하에서 역전사 반응을 실 시하고, EGTA로 킬레이트한 후 MgCl2를 첨가해 PCR을 실 시하는 방법이 이용되고 있다[9]. 최근, 우리들은 새로운 고 온균 Thermus thermophilus HJ6 균주를 분리하고 이 균주 의 DNA polymerase(Tod polymerase) 유전자의 클로닝 및 대장균에서의 발현에 대해 보고한 바 있다[10]. Tod poly- merase 유전자는 5'→3' exonuclease domain과 5'→3' poly- merase domain을 포함하는 834개의 아미노산으로 구성된다.
DNA polymerase는 dNTP가 없는 상태에서 선상 DNA 단 편과 함께 존재하면 단편을 분해하는 exonuclease 활성을 나 타내기 때문에 PCR 반응에 있어서는 불리한 반응이다.
본 연구에서는 Tod polymerase에서 5'→3' exonuclease 활성을 나타내는 N-말단 영역의 250개 아미노산을 결실시 킨 ∆Tod polymerase를 대장균에서 발현정제하여 그 특성을 분석하고 RT-PCR에의 적용을 시도하였다.
재료 및 방법 균주 및 플라스미드
본 연구에 사용된 Thermus thermophilus HJ6 균주는 일 본의 아리마(Arima) 온천수에서 분리한 것을 사용하였고[10], ATCC medium 697(8 g polypeptone, 4 g yeast extract, 4 g/L NaCl, pH 7.5)을 사용하여 80oC에서 배양 하였다. 대장
*Corresponding author
Tel: 82-2-51-890-2278, Fax: 82-2-51-890-2632
E-mail: [email protected]
균 DH5α와 BL21(DE3) codon plus(Novagen, Inc., San Diego, CA, USA)는 각각 클로닝 및 유전자 발현을 위한 균 주로 사용하였다. 플라스미드 pJLA503은 일본 오사카 대학 의 Kanaya연구실에서 분양 받은 것으로 단백질 발현을 위 해 사용하였다.
∆Tod polymerase의 생산 및 정제
Tod polymerase 유전자에서 N 말단의 250 아미노산에 해 당하는 유전자를 결실하기 위하여 Tod polymerase 유전자 가 삽입된 플라스미드 pGEMTOD를 주형으로 Primer 1(5'- GGCTCTCCCATATGCGCACCGACC-3', 밑줄은 Nde I 부 위)과 Primer 2(5'-TGGAATTCCTAACCCTTGGCGGAA- 3', 밑줄은 Eco RI 부위)를 이용하여 PCR을 수행하였다. 증 폭된 1.75 kb의 DNA 단편은 Nde I과 Eco RI으로 절단한 후 같은 효소로 절단한 pJLA503 벡터에 연결하고 E. coli DH5α에 형질전환 하였다. 클로닝 된 plasmid를 pJTODexo 로 명명하고 E. coli BL21(DE3) codon plus에 다시 형질전 환한 후 2×YTA 배지(tryptone 16 g/L, yeast extract 10g/
L, NaCl 5 g/L, ampicillin 100 µg/mL)에서 30oC로 20시 간 전배양 하였다. 전배양액의 1%를 새로운 2×YTA 배지에 접종한 후, 흡광도(A600)가 0.6이 되었을 때 42oC로 온도를 올려 5시간 동안 유도배양 하였다. 배양된 균체는 원심분리 를 통하여 집균하고 buffer A(50 mM Tris-HCl, pH 8.0, 1 mM dithiothreitol)에 녹인 다음 초음파로 파쇄 하였다. 초음 파 파쇄 후 세포추출물은 80oC에서 20분간 열처리한 후 원 심분리 하여 상등액을 회수하였다. 상등액은 ÄKTAprime(GE Healthcare)을 이용하여 buffer A로 평형화시킨 HiTrap Heparin column(GE Healthcare)에 로딩한 후 0~1.0 M의 NaCl로 농도구배를 주어 결합된 단백질을 용출하였다. 용출 된 단백질의 정제도는 SDS-PAGE(10% acrylamide gel)를 통하여 확인하였고, 전기영동 밴드는 coomassie 염색으로 확 인하였다.
Exonuclease 활성 측정
5' 말단이 표식된 DNA 기질을 만들기 위하여 PCR 반응 으로 2.5 kb의 Tod polymerase 유전자를 증폭하고 [γ-
32P]ATP와 T4 polynucleotide kinase를 이용하여 5' 말단을 인산화하였다. 3' 말단 표식을 위해서는 pET-21a 플라스미 드를 제한효소 Not I으로 절단하고 [α-32P]dCTP와 Klenow fragment를 이용하여 돌출말단을 수복하였다. 방사능 표식된 DNA 기질은 50 mM Tris-HCl(pH 8.5), 3 mM MgCl2, 1 mM DTT, 0.001% BSA 및 정제된 ∆Tod polymerase의 반 응 혼합액(50 µL)과 함께 75oC에서 반응하였다. 반응은 5%
trichloroacetic acid 1 mL을 첨가하여 정지시키고 원심 후 상등액을 회수하여 방사능 활성을 측정하였다[11].
Two-step RT-PCR 반응
2단계 역전사 반응은 PrimeScriptTM RT-PCR kit(Takara, Japan)를 사용하여 제조사에서 제시한 방법을 약간 변형하여 수행하였다. 20 mM Tris-HCl(pH 8.8), 0.1 mM dNTP(dATP, dCTP, dGTP, dTTP), 2 mM MgCl2 또는 MnCl2, oligo dT primer, 5 unit ∆Tod polymerase, plasmid pSPTet3 유래 RNA template의 다양한 양(5~250 ng)을 포함하는 혼합액 (20 µL)을 65oC에서 5분간 반응한 후, 60oC에서 30분간 역 전사 반응을 수행하였다. 반응 후 20 mM Tris-HCl (pH 8.8), 0.2 mM dNTP(dATP, dCTP, dGTP, dTTP), 2 mM MgCl2, 0.001% BSA, 0.1% Triton X-100, 50 pM F-1 primer (5'-CTGCTCGCTTCGCTACTTGGA-3'), 50 pM R-1 primer (5'-CGGCACCTGTCCTACGAGTTG-3')를 포함하는 PCR 혼합액(80 µL)을 첨가하고 DNA Thermal cycler(Takara, Japan)를 사용하여 PCR 반응을 수행하였다. PCR 조건은 95oC, 5분간의 변성단계를 거쳐 94oC에서 30초, 60oC에서 30초, 72oC에서 1분간을 30회 반복 수행하고 72oC에서 5분 간 마지막 신장반응을 수행하였다. PCR 반응액(5 µL)은 1.5% agarose 전기영동을 통하여 분석하였다. ∆Tod poly- merase의 DNA polymerase 활성은 최 등[3]의 방법에 따라 실시하였고 효소활성의 1 unit은 75oC에서 10분간 1 pmol 의 DNA를 합성하는 것으로 정의하였다.
One-step RT-PCR 반응
역전사 반응과 PCR 반응을 단일 튜브에서 수행하는 one- step RT-PCR 반응은 다음과 같은 방법으로 실시하였다. 20 mM Tris-HCl(pH 8.8), 0.25 mM dNTP(dATP, dCTP, dGTP, dTTP), 2 mM MgCl2, 0.001% BSA, 0.1% TritonX-100, 50 pM 16s-F primer(5'-AGAGTTTGATCATGGCTCAG-3'), 50 pM 16s-R primer(5'-AAGGAGGTGATCCAGCC-3'), 10 unit
∆Tod polymerase, E. coli DH5α 유래 total RNA template 의 다양한 양을 포함하는 혼합액(100 µL)을 65oC에서 5분 간 반응한 후, 60oC에서 30분간 역전사 반응을 수행하였다.
여기서 16s-F와 16s-R primer는 대장균의 16s rDNA를 증 폭할 수 있는 공통 염기서열을 바탕으로 설계하였다.
Thermal cycler(Takara, Japan)를 사용하여 95oC, 2분간의 변성단계를 거쳐 94oC에서 30초, 65oC에서 30초, 72oC에서 1분간을 30회 반복 수행하고 72oC에서 5분간 마지막 신장 반응을 수행하였다. Taq polymerase(Takara, Japan)를 사용 한 RT-PCR 반응은 제조사에서 제공된 buffer, 50 pM 16s- F primer, 50 pM 16s-R primer 및 total RNA를 혼합하고 위 와 동일한 방법으로 반응하였다. RT-PCR 반응액(5 µL)은 1.5% agarose 전기영동을 통하여 분석하였다. 증폭된 DNA 단 편은 densitometry(Gel-Pro Analyzer 3.1, MediaCybernetics, MD, USA)를 사용하여 정량 분석하였다.
결과 및 고찰
∆Tod polymerase의 생산
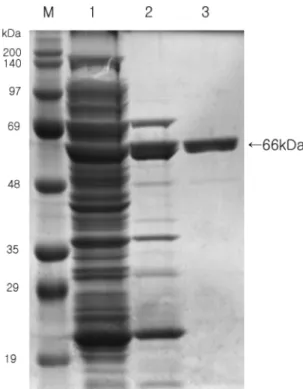
Tod polymerase 유전자는 Taq DNA polymerase와 유사 하게 2가지 domain으로 구성된다. 첫 번째는 5'→3' exo- nuclease domain이고 두 번째는 5'→3' polymerase domain 이다. DNA polymerase의 5'→3' exonuclease 활성은 PCR 반응에서 프라이머를 분해하는 등의 저해 요인으로 작용한 다. Taq DNA polymerase의 경우 N-말단의 289 아미노산에 해당하는 유전자를 결실시켜 중합 활성과 안정성에는 큰 영 향을 미치지 않고 5'→3' exonuclease을 성공적으로 제거하 였다[8]. 또한 Tfi DNA polymerase에서도 N-말단의 283 아 미노산을 결실시켜 동일한 효과를 나타낸다고 보고된 바 있 다[2]. 본 연구에서는 Tod polymerase 유전자에서 5'→3' exonuclease 활성에 관여하는 부분(개시코돈에서 750 bp)이 결실된 Tod 유전자를 발현 벡터 pJLA503에 클로닝하고 plasmid pJTODexo로 명명하였다. 효소 생산을 위한 숙주로 는 E. coli BL21(DE3) codon plus 균주를 사용하였다. Tod polymerase 유전자에는 대장균에서 잘 사용되지 않는 rare codon을 다수 가지고 있기 때문에 rare tRNA 합성효소가 보 강된 codon plus 균주를 사용하였다. Plasmid pJTODexo로 E. coli BL21(DE3) codon plus를 형질전환하고 30oC에서 배양하여 세포성장이 600 nm에서 흡광도가 0.5~0.6에 도달 하였을 때 온도를 42oC로 올려 단백질의 생산을 유도하였 다. 배양한 세포를 초음파로 파쇄하고 80oC에서 20분간 열 처리한 후, DNA-binding protein에 친화력을 가지는 HiTrap Heparin column(GE Healthcare)을 이용하여 한 단계로 순수 정제하였다. 정제된 단백질을 SDS-PAGE로 분석한 결과, 약 66 kDa 부위에서 ∆Tod polymerase의 밴드가 확인되었다 (Fig. 1, lane 3). 이 밴드의 분자량은 N 말단을 제거한 아미 노산 서열에서 계산된 분자량(65,645 Da)과도 잘 일치하였다.
∆Tod polymerase의 exonuclease 활성
DNA polymerase는 dNTP가 없는 상태에서 선상 DNA 단편과 함께 존재하면 DNA 단편을 분해하는 exonuclease 활성을 나타낸다. N-말단 250개의 아미노산을 결실한 ∆Tod polymerase의 경우 5'→3' exonuclease 활성을 나타내지 않 았다(Fig. 2). 반면, 야생형 Tod polymerase[8]의 경우 30분 만에 기질 DNA로부터 87%의 인산기를 제거하는 exo- nuclease 활성을 나타내었다(Fig. 2). 이 결과는 ∆Tod poly- merase에서 5'→3' exonuclease 활성이 완전히 제거 되었다 는 것을 의미한다. 한편, 야생형과 결실 변이형 Tod poly- merase는 모두 3'→5' exonuclease 활성에 대해서 거의 활성 을 보이지 않았다(data not shown). 이 결과는 Thermus filiformis 유래 DNA polymerase에서 보고한 것과도 일치하 였다[2].
∆Tod polymerase의 RT-PCR 조건
내열성 DNA polymerase의 역전사 반응에 대한 활성은 cDNA 합성에 대해 이온의 종류와 이온강도에 영향을 받는 것으로 조사된 바 있다[6]. 본 실험에서는 ∆Tod polymerase 의 역전사 활성을 조사하기 위하여 각기 다른 양(5~250 ng)
Fig. 1. Purification of ∆Tod polymerase. Lane M, molecular mass marker; lane 1, crude extract induced cells; lane 2, superna- tant of crude extract after heat treatment at 80
oC for 20 min; lane 3, Heparin column peak fractions. The gel was stained with coo- massie brilliant blue.
Fig. 2. Comparison of the 5' →3' exonuclease activity of ∆Tod
polymerase with that of Tod polymerase. The 5' →3' exonu-
clease activity of ∆Tod polymerase (
○) and Tod polymerase (
●)
was assayed in the absence of dNTPs. AR-RA, radioactivity of
supernatant solution after reaction; BR-RA, radioactivity of the 5'
end-labeled DNA substrate before reaction.
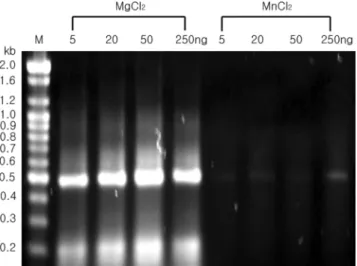
의 RNA 주형과 Mg2+및 Mn2+이온을 반응액에 첨가하여 역전사 반응을 수행하고, 생성된 cDNA를 주형으로 PCR 반 응을 수행하였다. 그 결과 Mg2+의 존재 하에서는 5, 20, 50, 250 ng을 주형 RNA로 사용한 반응에서 모두 cDNA 단편 이 강하게 증폭되었지만, Mn2+존재 하에서는 모든 반응에 서 매우 약한 cDNA 단편의 증폭이 확인되었다(Fig. 3). 그 러나 본 효소와 98.5%의 상동성을 보이는 Thermus
thermophilus HB8유래의 Tth DNA polymerase[9]는 역전 사 반응에서 MnCl2이 존재할 경우 효율적인 역전사 반응을 수행하여 본 연구의 결과와는 대조적인 모습을 나타내었다.
이것은 Tod polymerase의 N-말단을 제거한 것에 따른 단백 질의 구조적인 변화에 기인할지도 모른다. 이와 같이 기존 의 Tth DNA polymerase는 역전사 반응에서 사용한 MnCl2
를 킬레이트한 후 PCR 반응을 수행해야 하지만, ∆Tod polymerase는 Mg2+존재 하에서 역전사 반응과 PCR 반응 을 함께 수행할 수 있기 때문에 one-step RT-PCR 반응에 대 해 매우 실용적일 것으로 생각된다.
∆Tod polymerase의 one-step RT-PCR에 대한 적용 One-step RT-PCR에서는 역전사 반응과 PCR 반응을 같은 tube 내에서 연속해서 실시하기 때문에 조작이 간편하고 sample간에 오염의 위험이 낮다. 한편, 역전사 반응과 PCR 을 각각 최적인 조건으로 실시할 수 없기 때문에, 실험에 따 라 Two-Step RT-PCR 보다 반응 효율이 낮을 수도 있다. 본 실험에서는 Mg2+존재 하에서 ∆Tod polymerase가 one-step RT-PCR 반응에 적합한지를 시험하였다. 실험 대상으로 2차 구조를 취하기 쉬운 rRNA를 대상으로 one-step RT-PCR을 실시한 결과, Fig. 4에서와 같이 기존의 Taq polymerase를 사용한 반응에서는 50, 100, 250 ng의 RNA 농도에서만 cDNA가 증폭된 반면, ∆Tod polymerase의 one-step RT- PCR 반응에서는 5, 20, 50, 100, 250 ng의 모든 RNA 농도 에서 cDNA가 증폭하여, 미량의 RNA에서도 높은 효율의 RT-PCR 결과를 나타내었다. 대장균의 total RNA를 주형으 로 사용한 one-step RT-PCR에서는 plasmid를 주형으로 사 용한 two-step RT-PCR 보다 주형의 양이 상대적으로 적기
Fig. 3. Analysis of RT-PCR products for optimal RT-PCR con-
dition. The cDNA synthesis was performed in a 20 µL reaction mixture containing 2 mM MgCl
2or MnCl
2. The amounts of pSPTet3 cRNA used for cDNA synthesis are indicated at the top.
PCR was performed as described in the Materials and methods section. A 5 µL sample of each mixture was subjected to electro- phoresis on 1.0% agarose gel and stained with ethidium bromide.
Lane M, 100 bp DNA ladder markers.
Fig. 4. Reverse transcription and PCR amplification by either ∆Tod pol or Taq pol. The cDNA synthesis and amplification with
primer pair 16s-F/16s-R and total RNA of E. coli DH5α were performed as described in materials and methods section. ∆Tod polymerase
or Taq polymerase were used for both cDNA synthesis and PCR amplification from 5, 20, 50, 100, 250 ng of RNA. A 5 µL sample of each
mixture was subjected to electrophoresis on 1.0% agarose gel and stained with ethidium bromide. Lane M, λ Hind III markers.
때문에 DNA 증폭율이 감소하는 결과를 나타냈다(Fig. 3, 4).
One-step RT-PCR은 2차 구조를 취하기 쉬운 rRNA를 sample로 사용하는 경우와 RNA 바이러스 검출 등의 검사 시 오염의 방지가 최우선 되는 경우에 매우 유용하기 때문 에 ∆Tod polymerase는 이와 같은 시료의 RT-PCR 반응에 적용할 수 있을 것으로 생각된다.
요 약
고온균 Thermus thermophilus HJ6 유래의 N-말단 결실 Tod polymerase(∆Tod polymerase)는 온도 감수성 프로모터 (lambda pR and pL)를 포함하는 pJLA503 벡터를 이용하여 대장균에서 발현하였다. N-말단 250개 아미노산이 제거된
∆Tod polymerase는 5'→3' exonuclease 활성은 없어지고 DNA 중합반응의 활성은 그대로 유지되었다. ∆Tod poly- merase는 MgCl2의 존재 하에서 매우 효율적으로 역전사 반 응과 PCR 반응을 수행하였다. 또한 ∆Tod polymerase는 one-step RT-PCR 반응에서 Taq polymerase 보다 높은 cDNA 증폭 효율을 나타내었다.
감사의 글
이 논문은 2009학년도 동의대학교 교내연구비에 의해 연 구 되었음(과제번호 2009AA193).
R EFERENCES
1. Chien, A., D. B. Edgar, and J. M. Trela. 1976. Deoxyri- bonucleic acid polymerase from the extreme thermophile Thermus aquaticus. J. Bacteriol. 127: 1550-1557.
2. Choi, J. J., H. K. Kim, and S. T. Kwon. 2001. Purification and characterization of the 5'→3' exonuclease domain- deleted Thermus filiformis DNA polymerase expressed in Escherichia coli. Biotech. Lett. 23: 1647-1652.
3. Choi, J. J., S. E. Jung, H. K. Kim, and S. T. Kwon. 1999.
Purification and properties of Thermus filiformis DNA polymerase expressed in Escherichia coli. Biotechnol. Appl.
Biochem. 30: 19-25.
4. Frohman, M. A., M. K. Dush, and G. R. Martin. 1988. Rapid production of full-length cDNAs from rare transcripts:
amplification using a single gene-specific oligonucleotide primer. Proc. Natl. Acad. Sc.i USA. 85: 8998-9002.
5. Gibbs, R. A., P. N. Nguyen, L. J. McBride, S. M. Koepf, and C. T. Caskey. 1989. Identification of mutations leading to the Lesch-Nyhan syndrome by automated direct DNA sequenc- ing of in vitro amplified cDNA. Proc Natl Acad Sci USA.
86: 1919-1923.
6. Grebennikova, T. V., A. I. Glukhov, L. G. Chistiakova, V. I.
Kiselev, and E. S. Severin. 1995. Use of thermostable DNA polymerase from Thermus thermophilus KTP in a combined reverse transcription and amplification reaction of detecting interleukin 2alpha RNA and determining expression of the multidrug resistance gene (MDR-1). Mol. Biol. (Mosk). 29:
930-941.
7. Kawasaki, E. S., S. S. Clark, M. Y. Coyne, S. D. Smith, R.
Champlin, O. N. Witte, and F. P. McCormick. 1988. Diagnosis of chronic myeloid and acute lymphocytic leukemias by detection of leukemia-specific mRNA sequences amplified in vitro. Proc. Natl. Acad. Sc.i USA. 85: 5698-5702.
8. Lawyer, F. C., S. Stoffel, R. K. Saiki, K. Myambo, R.
Drummond, and D. H. Gelfand. 1989. Isolation, characteri- zation, and expression in Escherichia coli of the DNA polymerase gene from Thermus aquaticus. J. Bio.l Chem.
264: 6427-6437.
9. Myers, T. M. and D. H. Gelfand. 1991. Reverse transcrip- tion and DNA amplification by a Thermus thermophilus DNA polymerase. Biochemistry. 30: 7661-7666.
10. Seo, M. H., B. K. Kim, P. H. Kwak, H. W. Kim, Y. H. Kim, S. W. Nam, and S. J. Jeon. 2009. Gene Cloning and Expression of thermostable DNA polymerase from Thermus thermophilus HJ6. Kor. J. Microbiol. Biotechnol. 37: 17-23.
11. Sorokine, I., K. Ben-Mahez, M. Nakayama, and M. Kohiyama 1991. Exonuclease activities associated with DNA poly- merase α and β of the archaebacterium Halobacterium halobium. Eur. J. Biochem. 197: 781-784.
12. Tse, W. T., and B. G. Forget. 1990. Reverse transcription and direct amplification of cellular RNA transcripts by Taq polymerase. Gene. 88: 293-296.
13. Veres, G., R. A. Gibbs, S. E. Scherer, and C. T. Caskey.
1987. The molecular basis of the sparse fur mouse mutation.
Science. 237: 415-417.
(Received March 4, 2010/Accepted March 31, 2010)