Evaluation of the Genetic Diversity of Biovar 3 Strains of Pseudomonas syringae pv. actinidiae Isolated in Korea
Young Sun Lee1, Gyoung Hee Kim2, Young Jin Koh2 and Jae Sung Jung1*
1Department of Biology, Sunchon National University, Suncheon 57922, Korea
2Department of Plant Medicine, Sunchon National University, Suncheon 57922, Korea
Received October 18, 2019 /Revised November 20, 2019 /Accepted November 21, 2019
Pseudomonas syringae pv. actinidiae, the causal agent of a bacterial canker disease in kiwifruit, is sub- divided into five genetically distinct populations, namely biovars 1, 2, 3, 5, and 6. Of these, strains belonging to biovar 3 are responsible for a pandemic bacterial canker of kiwifruits since 2008. This study aimed to characterize the structure of the biovar 3 population and investigate the origin of bio- var 3 strains isolated in Korea. The genetic variability of fifteen biovar 3 strains, thirteen Korean and two Chinese, were evaluated through random amplified polymorphic DNA (RAPD)-PCR. The RAPD results revealed the presence of eight lineages, designated as subgroups I-VIII, across the biovar 3 strains used in this study. As the strains in subgroups II and III from China were not found in the Korean examples, we concluded that six genetically different biovar 3 subgroups (I, IV, V, VI, VII, and VIII) are present in Korea. In PCR analysis using primers specific to the strains of New Zealand and Europe, Korean strains in subgroups V and VI amplified the relevant DNA bands, suggesting that these were introduced from these two origins, respectively. PCR primers specific to subgroup VIII were developed to monitor the spread of the first biovar 3 strain in Korea, and investigations revealed that this strain was not found in Korea after its first occurrence.
Key words : Bacterial canker, biovar 3, genetic diversity, kiwifruit, Pseudomonas syringae pv. actinidiae
*Corresponding author
Tel : +82-61-750-3616, Fax : +82-61-750-3208 E-mail : [email protected]
This is an Open-Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Life Science 2020 Vol. 30. No. 1. 1~9 DOI : https://doi.org/10.5352/JLS.2020.30.1.1
Introduction
Pseudomonas syringae pv. actinidiae (PSA) is the causal agent of bacterial canker of kiwifruits (Actinidia deliciosa and A. chi- nensis). Presently, PSA is classified into five biovars based on biochemical, pathogenic, and genotypic characteristics.
The strains of biovar 1 were initially isolated from Japan in 1984 [36] and Italy prior to 2008 [34]; these strains produce the phytotoxin phaseolotoxin. Biovar 2 represents the strains isolated from Korea [23], and produces coronatine instead of phaseolotoxin [16]. Biovar 3 was first isolated in 2008, in central Italy, from kiwifruit orchards showing a sudden and severe outbreak of bacterial canker [11]. The strains belong- ing to biovar 3 were subsequently observed in most kiwi- fruit-producing countries in Europe [1, 2, 4, 9, 18, 39] and outside Europe, in Chile [8] and New Zealand [10] since
2010. Biovar 3 strains do not produce phaseolotoxin or coro- natine [27]. However, they are thought to be more aggressive than the other biovars owing to their rapid spreading ability, and were responsible for the current pandemics, which caused severe economic losses in kiwifruit-producing coun- tries worldwide. The biovar 5 strains were found in limited local areas of Japan. They formed a district clade, which was closely related to biovar 2 in a phylogenetic tree based on multilocus sequence analysis (MLSA) using concatenated se- quences of seven housekeeping genes [32]. They did not have the coronatine biosynthesis genes found in biovar 2 or the phaseolotoxin biosynthetic genes detected in biovar 1 [14]. Recently, biovar 6 was reported from Japan. This pathogen was different from the other biovars in biochemi- cal characteristics, and formed a distinct cluster upon multi- locus sequence analysis with the four existing biovars. They have unique characteristics that produce both paseolotoxin and coronatine, simultaneously [31]. Although the strains belonging to this biovar were less aggressive and caused on- ly mild symptoms such as leaf spotting, biovar 4 was ini- tially thought to be a new biovar of PSA [40]. However, based on the genotypic, phenotyphic, and phylogenetic dif- ferences, biovar 4 was proposed as a new pathovar of P.
syringae and renamed as Pseudomonas syringae pv. actinidifo-
liorum [7, 12]. Therefore, currently five biovars (biovars 1, 2, 3, 5 and 6) remain in PSA. Among them, four biovars (biovars 1, 3, 5 and 6) are found in Japan [31, 32], while two biovars (biovar 2 and 3) are reported in Korea [22, 26].
However, all PSA strains found within China are members of biovar 3 [17, 29].
In Korea, the bacterial canker of kiwifruit was first found in 1988 from green-fleshed A. deliciosa cv. ‘Hayward’, which was cultivated in Jeju [23], and has subsequently occurred from yellow- and red-fleshed kiwifruits of A. chinensis since 2006 [24]. The pathogen isolated from the canker was identi- fied as PSA biovar 2 [26]. PSA biovar 3, responsible for the current pandemic disease in all kiwifruit-producing coun- tries, was isolated for the first time from an orchard in Korea in 2011 [25].
In this study, we aimed to determine the population struc- ture of biovar 3, which was propagated rapidly between 2014 and 2016 in Korea.
Materials and Methods
Bacterial isolation
PSA biovar 3 strains were obtained as follows: PSA strains were isolated from 46 kiwifruit orchards infected with the disease in all kiwifruit cultivation areas of Korea during 2014 to 2016. Fragments of diseased plant tissue were cut with a blade and suspended in sterile distilled water. The suspen- sions were diluted and plated on peptone sucrose agar me- dium (PS agar; 20 g peptone, 20 g sucrose, 15 g agar per 1 liter). The plates were incubated at 28℃ for 48 hr. Bacterial colonies presenting similar morphological characteristics with PSA were selected and streaked on PS agar for single colony isolation. The preliminary identification of the iso- lates was performed using standard bacteriological methods proposed by Takikawa et al. [36].
Identification of biovar 3 strains of P. syringae pv.
actinidiae
For the identification and confirmation of PSA, PCR was performed using the primer set, P. syringae pv. actinidiae F/R (CAGAGGCGCTAACGAGGAAA/ CGAGCATACATCAAC AGGTCA). This primer pair was designed based on hopZ3 of PSA and amplified a DNA fragment of 311 bp from PSA strains isolated from all over the world, regardless of their biovars [3]. Biovar 3 strains were screened using Tac F/R primer pair (CGGGCTAGACAGTACGCTGT/CAGGCCCT
TCTACCGCTAC), which specifically amplified a 545 bp fragment among the strains identified as PSA [22]. A total of 53 biovar 3 strains were recovered from symptomatic samples of kiwifruit plants.
Genomic DNA extraction
Genomic DNA was isolated from bacteria using an AccuPrep genomic DNA extraction kit (Bioneer, Korea) ac- cording to the manufacturer’s instruction. The DNA concen- tration was measured with NanoDrop 2000 spectropho- tometer (Thermo Fisher Scientific, USA).
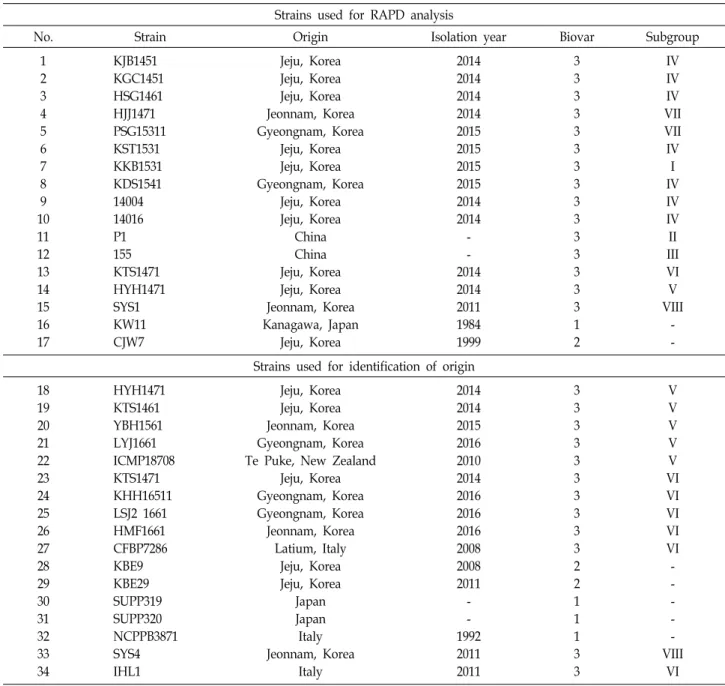
Bacterial strains used for RAPD analysis
Among 53 biovar 3 strains isolated in Korea, 13 repre- sentative strains were selected by the preliminary RAPD as- says with the criteria of band pattern similarity. Additionally, two biovar 3 strains obtained from China and one strain each of biovar 1 and 2 were included in the collection. The details about the bacterial strains such as name, year, and location of isolation are listed in Table 1.
RAPD-PCR
The DNA amplification reactions were carried out in a final volume of 50 μl, using a PCR Thermal Cycler Dice (Takara Bio Inc., Japan). The reaction mixture contained 5 μl of 10X reaction buffer (100 mM Tris-HCl, pH 8.0, 500 mM KCl, 25 mM MgCl2), 200 μM of each dNTPs, 20 pmol of random primer (Operon Biotechnologies, USA), 2.0 U of Taq DNA polymerase (Bioneer, Korea), and 20 ng of template DNA. A total of thirty 10-mer primers were chosen to obtain RAPD patterns for 17 PSA strains. The amplification con- ditions were as follows: Initial denaturation at 94℃ for 5 min followed by 40 cycles of denaturation at 94℃ for 30 sec, annealing at 37℃ for 1 min, and extension at 72℃ for 1 min. A final extension at 72℃ was performed for 7 min.
The amplification products obtained from PCR were electro- phoresed on 2% agarose gel along with a 100 bp standard DNA ladder (Bioneer, Korea). The amplicons were vi- sualized using a Gel-Doc system (Bio-Rad, USA) after stain- ing with ethidium bromide. The RAPD experiments were duplicated to confirm the banding pattern and consistent bands were scored for data analysis.
Data analysis
RAPD data files were analyzed using Gelcompar II soft- ware version 4.6 (Applied Maths, Belgium). Bands with the
Table 1. Pseudomonas syringae pv. actinidiae strains used in this study
Strains used for RAPD analysis
No. Strain Origin Isolation year Biovar Subgroup
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17
KJB1451 KGC1451 HSG1461 HJJ1471 PSG15311 KST1531 KKB1531 KDS1541 14004 14016 P1 155 KTS1471 HYH1471 SYS1 KW11 CJW7
Jeju, Korea Jeju, Korea Jeju, Korea Jeonnam, Korea Gyeongnam, Korea
Jeju, Korea Jeju, Korea Gyeongnam, Korea
Jeju, Korea Jeju, Korea
China China Jeju, Korea Jeju, Korea Jeonnam, Korea Kanagawa, Japan
Jeju, Korea
2014 2014 2014 2014 2015 2015 2015 2015 2014 2014 - - 2014 2014 2011 1984 1999
3 3 3 3 3 3 3 3 3 3 3 3 3 3 3 1 2
IV IV IV VII VII IV I IV IV IV II III VI V VIII
- - Strains used for identification of origin
18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34
HYH1471 KTS1461 YBH1561 LYJ1661 ICMP18708 KTS1471 KHH16511 LSJ2 1661 HMF1661 CFBP7286 KBE9 KBE29 SUPP319 SUPP320 NCPPB3871 SYS4 IHL1
Jeju, Korea Jeju, Korea Jeonnam, Korea Gyeongnam, Korea Te Puke, New Zealand
Jeju, Korea Gyeongnam, Korea Gyeongnam, Korea Jeonnam, Korea
Latium, Italy Jeju, Korea Jeju, Korea
Japan Japan Italy Jeonnam, Korea
Italy
2014 2014 2015 2016 2010 2014 2016 2016 2016 2008 2008 2011 - - 1992 2011 2011
3 3 3 3 3 3 3 3 3 3 2 2 1 1 1 3 3
V V V V V VI VI VI VI VI - - - - - VIII
VI
same mobility among individual PSA strains were consid- ered as identical fragments, regardless of the staining intensity. The RAPD data obtained using the 30 primers were combined and similarity values of DNA profiles were computed on the basis of band positions using the Dice sim- ilarity index [35]. The similarity values were then used to construct a dendrogram by the unweighted pair group method using arithmetic averages (UPGMA).
PCR for identifying geographic origin
The PCR primer sets that can identify European and New Zealand strains were developed by Balestra et al. [3]. The
primer pair, Europe F (TGGTGATCGTCTGGATGTGT) and Europe R (ATTATGCTCCTGGCTCATGG), produced a 733 bp product from the DNA of biovar 3 strains of PSA isolated in Europe. The primer pair, China F (GGAGTTCCAGCAAC TGACG) and China R (CGCTCAAGATCCTTTTCCAT), am- plified a 609 bp fragment from some strains isolated in China and all biovar 3 strains from New Zealand. Amplifications were carried out at the following conditions: Initial denatu- ration at 94℃ for 5 min followed by 40 cycles of denaturation at 94℃ for 30 sec, annealing at 60℃ for 30 sec, and extension at 72℃ for 30 sec. A final extension at 72℃ was performed for 7 min.
Fig. 1. Example of RAPD polymorphisms of Pseudomonas syringae pv. actinidiae strains using random 10-mer primers.
Lane M, 100-bp DNA ladder (Bion- eer); lanes 1 to 17 represent the strain numbers in Table 1. The primer name used for RAPD is indicated below the each gel photograph.
Development of specifis primers for SYS1 RAPD band unique to SYS1, the first biovar 3 strain iso- lated in Korea, was recovered from agarose gel and purified by AccuPrep Gel Purification Kit (Bioneer, Korea). The puri- fied fragment was ligated into the pGEM T-Easy vector sys- tem (Promega, USA) and transformed into competent Escher- ichia coli JM109. The inserted DNA fragment was sequenced by SolGent Co. (Korea). The primers specific for SYS1 was designed with the Primer 3 software based on internal se- quences of the inserted fragment. The nucleotide sequences of primers designed are as follows: PKN-F2, 5’-AGCGAGTC GCTGAATTGCGAAC-3’ and PKN-R4, 5’-ATGCTCGATAC ACCACACAATG-3’. This primer pair was then tested for its ability to produce an amplicon specific to SYS1. PCR con- ditions for amplification with the PKN-F2/R4 primers were identical to those with the Europe F/R primers except that the annealing temperature was 65℃.
Results and Discussion
RAPD is one of the efficient methods for the differentiation of population of PSA. The genetic diversity of biovar 3 strains of PSA isolated in Korea was assessed by RAPD. In a first trial, sixty eight 10-mer primers with arbitrary se- quences were tested on five PSA strains in order to select suitable primers for RAPD. Thirty primers produced reliable fingerprints for the bacterial strains. The selected 30 primers were used to analyze genomic DNAs of 17 representative
strains of PSA, including 13 biovar 3 strains isolated in Korea, two biovar 3 from China, one biovar 1 and one biovar 2 strains. Examples of RAPD-PCR fingerprints obtained us- ing random primers are shown in Fig 1. The approximate range of band size was 250-2,500 bp. A total of 236 bands were obtained from 30 primers, and the number of poly- morphic amplicons in RAPD was 153, accounting for 64.8%.
Genetic diversity analysis
Based on the presence or absence of 153 RAPD bands, cluster analysis was performed to generate a dendrogram.
The UPGMA dendrogram (Fig. 2) showed that fifteen biovar 3 strains exhibited variability within the biovar and were clearly distinct from the reference strains of biovar 1 and biovar 2 at a similarity level of 0.46. The strains belonging to biovar 3 were separated into eight lineages at a Dice co- efficient of 0.93 and were named biovar 3 subgroup I to VIII (Table 1).
The strains of P1 and 155 belonging to subgroup II and III, respectively, were obtained in China. The results of re- cent studies revealed that biovar 3 strains from China have broad genetic variability and are divided into several sub- groups that are related to the geographic source location [17]. In preliminary experiments, we did not find any strain with the same RAPD profiles as these subgroups among 53 biovar 3 strains isolated in Korea. The strains belonging to the remaining six subgroups were isolated in Korea. The re- sult indicates that six genetically different subpopulation of
Fig. 2. Dendrogram generated by GelCompar II software showing the relationship between Pseudomonas syringae pv. acti- nidiae strains. The analysis of the bands was performed using the Dice coeffici- ent and unweighted pair group meth- od with arithmetic averages (UPGMA).
A
B
Fig. 3. PCR amplification with specific primers of New Zealand (A) and European strains (B). Lane M, 100-bp DNA lad- der (Bioneer); lanes 1 to 10 represent the strain numbers 18 to 27 in Table 1.
PSA biovar 3 are present in Korea.
Origin of subgroup V and VI strains
Although it was difficult to determine the sources of all subgroups identified as Korean biovar 3 strains, a few sub- groups could be traced to their origins. To determine if any of the biovar 3 subgroups isolated in Korea include strains belonging to New Zealand or European populations, PCR amplifications were carried out against 53 biovar 3 strains using two specific primer sets. As shown in Fig. 3, a 609 bp fragment was amplified using China F/R primers in DNAs from five strains including HYH1471, subgroup V strain, and strain ICMP18708 isolated from New Zealand, while the Europe F/R primer set produced a 733 bp frag- ment with DNAs of subgroup VI strain, KTS1471, and four strains including the European reference strain CFBP7286.
These results indicate that the subgroup V and VI strains might originate in New Zealand and Europe, respectively.
Origin of subgroup VIII strains
The subgroup VIII strain SYS1 is the first Korean PSA strain belonging to biovar 3, which was isolated in 2011 on A. chinensis cultivars from an orchard located in Goheung, Jeonnam province in Korea [25]. The strain SYS1 was not detected with primers designed for the identification of European or New Zealand strains. Therefore, we needed to develop a new primer pair specific to SYS1 to monitor the spread of this biovar 3 strain in Korea. Of the RAPD primers tested, OPA-15 primer produced distinct polymorphic band
of approximately 1,250 bp from the strain SYS1. This frag- ment was cloned and the nucleotide sequences were determined. Based on the sequence information, SYS1 specif- ic primers were developed. The designed primer pair named as PKN-F2 and PKN-R4 amplified the expected 771 bp of DNA from SYS1 and SYS4 isolated from the same orchard.
The amplicon was not produced in strains of biovar 1, biovar 2 and biovar 3 isolated from Italy and New Zealand (Fig.
4). None of the biovar 3 strains examined in this study pro-
Fig. 4. Agarose gel electrophoresis of PCR products amplified using the PKN-F2/4 primer pair and genomic DNA of Pseudomonas syringae pv. actinidiae strains. Lane M, 100- bp DNA ladder (Bioneer); lanes 1-3, biovar 2 strains (CJW7, KBE9 and KBE29); lanes 4-7, biovar 1 strains (KW11, SUPP319, SUPP320 and NCPPB3871); lanes 8-9, subgroup VIII strains of biovar 3 (SYS1 and SYS4); lanes 10-11, subgroup VI strains of biovar 3 (IHL1 and CFBP 7286); lane 12, subgroup V strain of biovar 3 (ICMP18709).
duced amplicon of the expected size in PCR with PKN-F2/
R4 primer pair, except SYS1 and SYS4 (data not shown). The orchard where SYS1 was isolated was completely shut down by the plant quarantine authority on September 11, 2014[19].
Thereafter, the strains belonging to subgroup VIII have not been found in Korea. Epidemiological studies indicated that SYS1 strain was introduced into Korea through the seedlings from China in 2006[20].
Dissemination of biovar 3 strains
Biovar 3 was detected from one among 32 (3.1%) orchards in 2013 in Korea where the bacterial canker was investigated and confirmed, but 96.9% of PSA strains isolated were bio- var 2. However, biovar 3 spread rapidly in 2014, and 44.0%
(33/75) of the orchards were infected with this biovar. In 2015, biovar 3 was detected in 55.0% (72/131) of the infected orchards and became a dominant PSA biovar in Korea [20].
The number of kiwifruit orchards infected with biovar 3 in- creased to 63.9% in 2016 [21]. However, biovar 3 strains iso- lated from diseased plants did not amplify a 771 bp DNA fragment with the PKN-F2/R4 primer pair, which was a mo- lecular marker of subgroup VIII strains. The biovar 3 strains detected in Korea after 2014 were thought to have an origin different from the initially isolated subgroup VIII.
The bacterial canker, which spread in Jeju in 2014, showed symptoms at a flowering stage, unlike the typical symptoms of canker appearing at branches and trunk in early spring.
Since the symptoms first appeared from the fruit-bearing branches after artificial pollination and gradually progressed toward trunk, the disease was thought to be caused by pol-
lens contaminated with PSA [20]. The possibility of trans- mission of bacterial canker by infected pollens has been re- ported earlier [37, 38].
The biovar 3 strains have genetic variability that is related to geographic origin. MLSA conducted on housekeeping genes and core genome sequencing data indicated that bio- var 3 isolates from the outbreaks are monophyletic and be- longed to the same genetic lineage [6, 28]. However, the se- quencing of PSA genomes isolated from Europe, New Zealand and Chile revealed that PSA strains of the three continents are very similar but genetically distinct from each other owing to diversity in genomic islands, integrative con- jugative elements (ICEs), resulting from recent horizontal transfer from the other pathovars of P. syringae to PSA. The ICEs found in strains from New Zealand, Italy and Chile were referred to as Pac_ICE1, Pac_ICE2, and Pac_ICE3, re- spectively [5].
The population structure of biovar 3 strain is reported to vary in other countries also. Biovar 3 strains isolated from Japan were divided into two subgroups depending on the presence or absence of ICE. One of the subgroups of Japanese biovar 3 strains had a Pac_ICE1 like New Zealand strains, while the other subgroup did not have any ICEs [33].
The results of repetitive sequence PCR with BOX primer of 22 biovar 3 strains isolated in 2013-2014 from Portugal revealed that the fingerprints of five strains were different from the rest of the strains, which were similar to Italian strains. They showed genetic characteristics distinct from the New Zealand strains, indicating that these five strains repre- sent a new population of biovar 3. Therefore, biovar 3 strains isolated in Portugal were divided into two subpopulations [15].
The genetic diversity of the PSA strains isolated from the same geographical area in Italy but at different times in 2010 and 2014 was examined by RAPD-PCR, rep-PCR and MLSA.
All PSA strains belonged to biovar 3. However, the strains isolated in 2014 were shown greater genetic diversity than the strains of 2010. The strains from northern Italy exhibited a high level of variability than the strains from central Italy.
The regional diversity of population was explained on the basis of different weather conditions, which might have af- fected the bacterial population structure [30].
Three of the six subgroups found in Korea could be traced their origins, but remaining three were unknown. It was dif- ficult to estimate whether the three unidentified subgroups were evolved from a common ancestor introduced into
Korea or imported from other countries. However, as re- ported by Firrao et al. [13] in the European and New Zealand strains, minor variations within subgroup IV (Fig.
2) may be attributed to the genome rearrangement asso- ciated with mobile DNA elements following their first in- troduction in Korea.
The strains belonging to subgroup II and III, obtained from China, were not isolated in Korea as mentioned above.
However, a large number of pollens were imported into Korea from China, and hence there is a possibility that some subgroups of unknown origin were introduced from China.
Several studies revealed that PSA biovar 3 strains isolated from different countries originated from the independent in- troduction of a variant strain from China [5, 28, 29].
However, some strains might have been introduced into Korea from Chile through the same route.
PSA biovar 3 was added to the list of plant pathogens to be quarantined on December 18, 2014 in Korea. The diver- sity observed in populations of biovar 3 strains was presum- ably due to the pollens imported before the beginning of quarantine inspection. However, the possibility of trans- mission by imported seedling or grafting scion cannot be excluded.
In conclusion, the PSA biovar 3 populations in Korea ex- hibit genetic variability and are divided into six genetically different subgroups. It is noteworthy that the biovar 3 strains found in Korea have high genetic diversity. In order to fur- ther clarify the population structure of biovar 3 in Korea, it is necessary to assess the diversity of the strains using genome sequencing.
The Conflict of Interest Statement
The authors declare that they have no conflicts of interest with the contents of this article.
References
1. Abelleira, A., Lopez, M. M., Penalver, J., Aguin, O., Mansilla, J. P., Picoaga, A. and Garcia, M. J. 2011. First report of bacte- rial canker of kiwifruit caused by Pseudomonas syringae pv.
actinidiae in Spain. Plant Dis. 95, 1583.
2. Balestra, G. M., Renzi, M. and Mazzaglia, A. 2010. First re- port of bacterial canker of Actinidia deliciosa caused by Pseudomonas syringae pv. actinidiae in Portugal. New Dis. Rep.
22, 10.
3. Balestra, G. M., Taratufolo, M. C., Vinatzer, B. A. and Mazzaglia, A. 2013. A multiplex PCR assay for detection
of Pseudomonas syringae pv. actinidiae and differentiation of populations with different geographic origin. Plant Dis. 97, 472-478.
4. Bastas, K. and Karakaya, A. 2012. First report of bacterial canker of kiwifruit caused by Pseudomonas syringae pv. acti- nidiae in Turkey. Plant Dis. 96, 452.
5. Butler, M. I., Stockwell, P. A., Black, M. A., Day, R. C., Lamont, I. L. and Poulter, R. T. M. 2013. Pseudomonas sy- ringae pv. actinidiae from recent outbreaks of kiwifruit bacte- rial canker belong to different clones that originated in China. PLoS One 8, e57464.
6. Chapman, J. R., Taylor, R. K., Wrir, B. S., Romberg, M. K., Vanneste, J. L., Luck, J. and Alexander, B. J. R. 2012.
Phylogenetic relationships among global populations of Pseudomonas syringae pv. actinidiae. Phytopathology 102, 1034- 1044.
7. Cunty, A., Poliakoff, F., Rivoal, C., Cesbron, S., Fischer-Le Saux, M., Lemaire, C., Jacques, M. A., Manceau, C. and Vanneste, J. L. 2014. Characterization of Pseudomonas sy- ringae pv. actinidiae (Psa) isolated from France and assign- ment of Psa biovar 4 to a de novo pathovar: Pseudomonas syringae pv. actinidifoliorum pv. nov. Plant Pathol. 64, 582-596.
8. EPPO. 2011. First report of Pseudomonas syringae pv. actini- diae in Chile. EPPO Rep. Serv. 3, 2011/055.
9. EPPO. 2013. First report of Pseudomonas syringae pv. actini- diae in Germany. EPPO Rep. Serv. 9, 2013/185.
10. Everett, K. R., Taylor, R. K., Romberg, M. K., Rees-George, J., Fullerton, R. A., Vanneste, J. L. and Manning, M. A. 2011.
First report of Pseudomonas syringae pv. actinidiae causing ki- wifruit bacterial canker in New Zealand. Australas. Plant Dis.
Notes 6, 67-71.
11. Ferrante, P. and Scortichini, M. 2009. Identification of Pseudomonas syringae pv. actinidiae as causal agent of bacte- rial canker of yellow kiwifruit (Actinidia chinensis Planchon) in central Italy. J. Phytopathol. 157, 768-770.
12. Ferrrante, P. and Scortichini, M. 2015. Redefining the global population of Pseudomonas syringae pv. actinidiae based on pathogenic, molecular and phenotypic characteristics. Plant Pathol. 64, 51-62.
13. Firrao, G., Torelli, E., Polano, C., Ferrante, P., Ferrini, F., Martini, M., Marcelletti, S., Scortichini, M. and Ermacora, P. 2018. Genomic structural variations affecting virulence during clonal expansion of Pseudomonas syringae pv. actini- diae biovar 3 in Europe. Front. Microbiol. 9, 656.
14. Fujikawa, T. and Sawada, H. 2016. Genome analysis of the kiwifruit canker pathogen of Pseudomonas syringae pv. actini- diae biovar 5. Sci. Rep. 6, 21399.
15. Garcia, E., Moura, L., Abelleira, A., Aguin, O., Ares, A. and Mansilla, P. 2018. Characterization of Pseudomonas syringae pv. actinidiae biovar 3 on kiwifruit in north-west Portugal.
J. Appl. Microbiol. 125, 1147-1161.
16. Han, H. S., Koh, Y. J., Hur, J. S. and Jung, J. S. 2003. Identifi- cation and characterization of coronatine-producing Pseudo- monas syringae pv. actinidiae. J. Microbiol. Biotechnol. 13, 110- 118.
17. He, R., Liu, P., Jia, B., Xue, S., Wang, X., Hu, J., Shoffe, Y.
A., Gallioli, L., Mazzaglia, A., Balestra, G. M. and Zhu, L.
2019. Genetic diversity of Pseudomonas syringae pv. actinidiae strains from different geographic regions in China. Phytopa- thology 109, 347-357.
18. Holeva, M. C., Glynos, P. E. and Karafla, C. D. 2015. First report of bacterial canker of kiwifruit caused by Pseudomonas syringae pv. actinidiae in Greece. Plant Dis. 99, 723.
19. Kim, G. H., Choi, E. D., Lee, Y. S., Jung, J. S. and Koh, Y.
J. 2016a. Spread of bacterial canker of kiwifruit by secondary infection of Pseudomonas syringae pv. actinidiae biovar 3 in Gyeongnam in 2016. Res. Plant Dis. 22, 276-283.
20. Kim, G. H., Kim, K. H., Son, K. I., Choi, E. D., Lee, Y. S., Jung, J. S. and Koh, Y. J. 2016b. Outbreak and spread of bacterial canker of kiwifruit caused by Pseudomonas syringae pv. actinidiae biovar 3 in Korea. Plant Pathol. J. 32, 542-551.
21. Kim, G. H., Jung, J. S. and Koh, Y. J. 2017. Occurrence and epidemics of bacterial canker of kiwifruit in Korea. Plant Pathol. J. 33, 351-361.
22. Koh, H. S., Kim, G. H., Lee, Y. S., Koh, Y. J. and Jung, J.
S. 2014. Molecular characteristics of Pseudomonas syringae pv.
actinidiae strains isolated in Korea and a multiplex PCR as- say for haplotype differentiation. Plant Pathol. J. 30, 96-101.
23. Koh, Y. J., Cha, B. J., Chung, H. J. and Lee, D. H. 1994.
Outbreak and spread of bacterial canker in kiwifruit. Kor.
J. Plant Pathol. 10, 68-72.
24. Koh, Y. J., Kim, G. H., Jung, J. S., Lee, Y. S. and Hur, J.
S. 2010. Outbreak of bacterial canker on Hort16A (Actinidia chinensis Planchon) caused by Pseudomonas syringae pv. acti- nidiae in Korea. N. Z. J. Crop Hortic. Sci. 38, 275-282.
25. Koh, Y. J., Kim, G. H., Koh, H. S., Lee, Y. S., Kim, S. C.
and Jung, J. S. 2012. Occurrence of a new type of Pseudomonas syringae pv. actinidiae strain of bacterial canker on kiwifruit in Korea. Plant Pathol. J. 28, 423-427.
26. Lee, Y. S., Kim, J., Kim, G. H., Choi, E. D., Koh, Y. J. and Jung, J. S. 2017. Biovars of Pseudomonas syringae pv. actinidiae strains, the causal agent of bacterial canker of kiwifruit, iso- lated in Korea. Res. Plant Dis. 23, 35-41.
27. Marcelletti, S., Ferrante, P., Petriccione, M., Firrao, G. and Scortichini, M. 2011. Pseudomonas syringae pv. actinidiae draft genomes comparison reveal strain-specific features involved in adaptation and virulence to Actinidia species. PLoS One 6, e27297.
28. Mazzaglia, A., Studholme, D. J., Taratufolo, M. C., Cai, R., Almeida, N. F., Goodman, T., Guttman, D. S., Vinatzer, B.
A. and Balestra, G. M. 2012. Pseudomonas syringae pv. actini- diae (PSA) isolates from recent bacterial canker of kiwifruit outbreaks belong to the same genetic lineage. PLoS One 7, e36518.
29. McCann, H. C., Li, L., Li, D., Pan, H., Zhong, C., Rikkerink, E. H. A., Templeton, M. D., Straub, C., Colombi, E., Rainey, P. B. and Huang, H. 2017. Origin and evolution of the kiwi- fruit canker pandemic. Genome Biol. Evol. 9, 932-944.
30. Prencipe, S., Gullino, M. L. and Spadaro, D. 2018. Pseudomo- nas syringae pv. actinidiae isolated from Actinidia chinensis var. deliciosa in northern Italy: genetic diversity and viru- lence. Eur. J. Plant Pathol. 150, 191-204.
31. Sawada, H., Kondo, K. and Nakaune, R. 2016. Novel biovar (biovar 6) of Pseudomonas syringae pv. actinidiae causing bac- terial canker of kiwifruit (Actinidia deliciosa) in Japan. Jpn.
J. Phytopathol. 82, 101-115.
32. Sawada, H., Miyoshi, T. and Ide, Y. 2014. Novel MLSA group (Psa5) of Pseudomonas syringae pv. actinidiae causing bacterial canker of kiwifruit (Actinidia chinensis) in Japan.
Jpn. J. Phytopathol. 80, 171-184.
33. Sawada, H., Shimizu, S., Miyoshi, T., Shinozaki, T., Kusu- moto, S., Noguchi, M., Naridomi, T., Kikuhara, K., Kansako, M., Fujikawa, T. and Nakaune, R. 2015. Characterization of biovar 3 strains of Pseudomonas syringae pv. actinidiae iso- lated in Japan. Jpn. J. Phytopathol. 81, 111-126.
34. Scortichini, M. 1994. Occurrence of Pseudomonas syringae pv.
actinidiae on kiwifruit in Italy. Plant Pathol. 43, 1035-1038.
35. Sneath, P. H. A. and Sokal, R. R. 1973. Numerical taxonomy.
The principles and practice of numerical classification.
Freeman, San Francisco, USA.
36. Takikawa, Y., Serizawa, S., Ichikawa, T., Tsuyumu, S. and Goto, M. 1989. Pseudomonas syringae pv. actinidiae pv. nov.:
the causal bacterium of kiwifruit canker in Japan. Ann. Phy- topathol. Soc. Japan 55, 437-444.
37. Tontou, R., Giovanardi, D. and Stefani, E. 2014. Pollen as a possible pathway for the dissemination of Pseudomonas sy- ringae pv. actinidiae and bacterial canker of kiwifruit. Phyto- pathol. Mediterr. 53, 333-339.
38. Vanneste, J. L., Giovanardi, D., Yu, J., Cornish, D. A., Kay, C., Spinelli, F. and Stefani, E. 2011. Detection of Pseudomonas syringae pv. actinidiae in kiwifruit pollen samples. N. Z. Plant Prot. 64, 246-251.
39. Vanneste, J. L., Poliakoff, F., Audusseau, C., Cornish, D. A., Paillard, S., Rivoal, C. and Yu, J. 2011. First report of Pseudo- monas syringae pv. actinidiae, the causal agent of bacterial canker of kiwifruit in France. Plant Dis. 95, 1311-1312.
40. Vanneste, J. L., Yu, J., Cornish, D. A., Tanner, D. J., Windner, R., Chapman, J. R., Taylor, R. K., Mackay, J. F. and Dowlut, S. 2013. Identification, virulence, and distribution of two bio- vars of Pseudomonas syringae pv. actinidiae in New Zealand.
Plant Dis. 97, 708-719.
초록:RAPD 지문을 통한 우리나라에서 분리된 Pseudomonas syringae pv. actinidiae biovar 3 균주의 유전적 다양성 평가
이영선1․김경희2․고영진2․정재성1*
(1순천대학교 생물학과, 2순천대학교 식물의학과)
키위 세균성 궤양병의 원인균인 Pseudomonas syringae pv. actinidiae는 유전적으로 구별되는 다섯 개의 biovar (1, 2, 3, 5, 6)로 나뉘어 진다. 그 중 biovar 3에 속하는 균주가 2008년 이래 전세계적으로 대유행을 일으키고 있다.
본 연구의 목표는 우리나라에서 분리된 biovar 3균주들의 집단 구조를 밝히고 그들의 기원을 추적하는데 있다.
13개의 우리나라 대표 균주와 2개의 중국 균주를 포함하는 15개 biovar 3 균주의 유전적 다양성을 RAPD-PCR로 평가하였다. RAPD 결과 연구에 사용된 균주들은 8개로 나누어져 이들을 subgroup I - VIII로 명명하였다.
Subgroups II와 III은 중국 균주로 우리나라에서 발견되지 않았다. 따라서 우리나라 biovar 3 균주는 유전적으로 구별되는 6개의 subgroup (I, IV, V, VI, VII, VIII)으로 구성되어 있음을 알 수 있었다. New Zealand와 Europe균 주에 특이적인 각각의 프라이머를 사용하여 PCR을 수행했을 때 subgroups V와 VI에 속하는 균주가 예상했던 DNA 절편을 증폭시켜 이들이 각각 두 지역에서 유입된 균주임을 시사하였다. 우리나라에서 처음 발견된 biovar 3 균주인 subgroup VIII에 특이적인 프라이머를 개발하여 조사한 결과 이 subgroup 균주는 처음 출현한 이후 더 이상 발견되지 않았다.