연자육 추출물의 멜라닌 합성 저해효과
이준영, 임경란, 정택규, 윤경섭*
The Inhibitory Effects of Nelumbo nucifera Gaertner Extract on Mel- anogenesis
Jun Young Lee, Kyung Ran Im, Taek Kyu Jung, and Kyung-Sup Yoon*
접수: 2013년 1월 14일 / 게재승인: 2013년 4월 25일
© 2013 The Korean Society for Biotechnology and Bioengineering
Abstract: In order to develop new skin whitening agents, we prepared the CH2Cl2 layer (NGC) and BuOH layer (NGB) of 75% EtOH extract of the Nelumbinis nucifera Gaertner. We measured their tyrosinase inhibitory activity in vitro and mel- anin synthesis inhibitory activity in B16-F1 melanoma cells.
They did not show inhibitory activity against mushroom tyro- sinase but showed melanin synthesis inhibitory activity in a dose-dependent manner. In a melanin synthesis inhibition assay, NGC and NGB suppressed melanin production up to 52% and 46% at a concentration of 100µg/mL, respectively.
To elucidate the mechanism of the inhibitory effects of NGC and NGB on melanogenesis, we measured the expression of melanogenesis-related proteins by western blot assay. As a result, NGC suppressed the expression of tyrosinase, tyrosi- nase related protein 1 (TRP-1), tyrosinase related protein 2 (TRP-2), phosphorylated cAMP responsive element binding (p-CREB) protein, and microphthalmia associated transcrip- tion factor (MITF). And NGB inhibited the protein expres- sion of tyrosinase and MITF, but had no significant effect on TRP-1, TRP-2, and p-CREB expression. Moreover, NGB increased the expression of phosphorylated extracellular sig- nal-regulated kinase (p-ERK). In addition, we examined the inhibitory effect on the glycosylation of tyrosinase. As a result, NGC and NGB inhibited the activity of α-glucosidase in vitro and the glycosylation of tyrosinase in B16-F1 mela-
noma cells. From these results, we concluded that NGC and NGB could be used as active ingredients for skin whitening.
Keywords: Nelumbo nucifera Gaertner, Melanogenesis, Tyro- sinase, MITF, Glycosylation
1. 서론
멜라닌은 태양광선 중 유해한 자외선으로부터 생체를 보호 하는 중요한 방어수단이나 멜라닌이 과도하게 합성되거나 노화 등에 의해 피부의 생리기능이 떨어지게 되면 멜라닌이 피부 표면에 침착되어 기미, 주근깨 등 다양한 색소 침착을 유발하게 된다 [1]. 멜라닌은 표피 기저층에 존재하는 멜라 닌세포의 멜라노좀에서 합성되어 멜라닌세포의 수지상 돌 기를 통해 피부 각질층으로 이동된다 [2].
멜라닌의 합성은 자외선, cytokine, growth factor 및 호르몬 등에 의해 조절되며 많은 인자들이 관여하는 매우 복잡한 과 정으로 몇 가지 세포내 신호전달 기전을 통하여 합성되는데, 주요 경로로서 cyclic adenosine monophosphate (cAMP) / pro- tein kinase A (PKA) 경로가 있다. 이는 피부가 UV에 노출되 었을 때 멜라닌세포의 cAMP 신호를 증폭시켜 하류 신호전 달 물질인 PKA의 활성화를 유도하며, 세포내 cAMP respon- se element binding (CREB) protein을 활성화시킴으로써 mi- crophthalmia-associated transcription factor (MITF)의 발현을 증가시킨다 [3,4]. MITF는 멜라닌 합성 과정에서 중요한 전 사 조절 인자로 tyrosinase와 tyrosinase-related proteins (TRPs;
TRP-1, TRP-2)의 합성을 증가시켜 멜라닌 합성을 촉진시키 는 것으로 알려져 있다 [5-7]. 따라서 MITF는 멜라닌 합성을
(주)사임당화장품 기술연구소
Saimdang Cosmetics Co., Ltd., R&D Center, Chungbuk 363-886, Korea
Tel: +82-43-218-4203, Fax: +82-43-218-4204 e-mail: [email protected]
연구논문
조절하는 skin-lightening agents를 개발하기 위한 주요 타깃 으로서 여겨지고 있다 [8-11]. 멜라닌 합성을 조절하는 또 다 른 경로로서 extracellular signal-regulated kinase (ERK) path- way에 대한 연구들이 보고되었다. 이는 ERK pathway를 활성 화시키면 MITF의 인산화를 유도하고 MITF가 ubiquitination (MITF-ⓟ-Ser73)으로 분해되어 결국 멜라닌 합성을 억제한 다는 것이다 [12-15]. 그 외에도 nitric oxide에 의한 cyclic gu- anosine monophosphate (cGMP) 경로, protein kinase C (PKC) 의 경로 등이 알려져 있다.
멜라닌은 크게 두 가지 타입이 만들어지는데, 노란색에서 붉은색의 색소를 보이는 pheomelanin과 검은색에서 갈색을 띠는 eumelanin이다 [16]. 멜라닌 합성의 주 효소인 tyrosinase 는 tyrosine을 3,4-dihydroxyphenyl-alanine (DOPA)으로 변환 시키고, 이것이 DOPA quinone으로 산화되는 것을 촉진하며, 5,6-dihydroxyindole (DHI)을 indole-5,6-quinone으로 산화하 는 것을 촉진한다. 반면 TRPs는 그 다음 단계인 eumelano- genesis에 관여한다 [7,17].
Tyrosinase는 type I membrane glycoprotein으로 [18], aspara- gine의 side chain에 있는 NH2기에 당을 붙이는 N-linked gly- cosylation 과정과 serine이나 threonine의 side chain에 있는 OH 기에 당을 붙이는 O-linked glycosylation 과정 [19] 중 N-linked glycosylation 과정에 의해 만들어진다. 단백질의 N-linked gly- cosylation은 endoplasmic reticulum (ER)에서 단백질 합성 번 역과 함께 일어난다 [20]. Glycan unit의 절반인 14개짜리의 당 (Glc3Man9GlcNAc2)은 ER막의 바깥인 세포질 쪽에서 만들 어지며 이후 나머지 절반은 ER내에서 만들어져 단백질로 전 이된다 [21]. ER에서 이런 합성 과정은 oligosaccharide의 말단 끝에 존재하는 mannose에서 3개의 glucose를 α-glucosidase I 과 II가 끊어줌으로써 시작된다 [22]. 그러므로 α-glucosidase 를 저해하게 되면 tyrosinase의 glycosylation이 저해되어 구조 는 변형되고 불활성 형태로 멜라노좀으로 이동하게 되어 결 과적으로 melanogenesis가 억제된다 [23-25]. 이와 같은 tyro- sinase glycosylation 저해제에 대한 보고들로는 Imokawa와 Mishima [26]이 B16 melanoma 세포배양에서 glucosamine과 tunicamycin 같은 물질들은 tyrosinase의 glycosylation 반응을 저해하여 멜라닌 합성이 완전히 억제될 수 있음을 보고하였 고, Franchi 등 [27]은 calcium D-pantetheine-S-sulphonate가 tyrosinase의 glycosylation을 저해하여 tyrosinase의 안정도를 저해하는 역할을 한다고 보고하였다. 그리고 Petrescu 등 [23]에 의하면, ER processing enzyme인 α-glucosidase I, II의 저해제인 N-butyl-deoxynojirimycin (NB-DNJ)이 B16 melano- ma 세포의 활성을 억제하며 비처리군에 비해 2%의 멜라닌 만이 합성되었다고 보고하여 tyrosinase glycosylation 저해제 에 대한 연구들이 이루어지고 있음을 알 수 있다.
이처럼 최근의 미백 물질에 대한 탐색 방법은 기존의 tyrosi- nase 활성을 억제하는 물질에 대한 연구뿐만 아니라 멜라닌 합성 기전에 관련된 단백질 발현을 조절하는 물질, N-glycosy- lation 억제제와 같이 멜라닌 합성 기전을 방해하여 미백효과 를 나타내는 물질 등 다양한 측면에서 진행되고 있다 [28-30].
연자육은 수련과 (Nymphaeaceae)에 속하는 다년생 수생식 물인 연꽃 (Nelumbo nucifera Gaertner)의 잘 익은 종자를 가 을에 수확하여 과피를 제거하여 말린 것으로 연밥 혹은 연자 로 불리며, 예부터 한방 및 민간에서 약재 혹은 식용으로 널 리 사용되어져 왔다.
연자육은 한방에서 주로 지혈, 어혈제거, 해혈, 토혈, 혈뇨, 혈변을 치료하는데 사용되고 있으며 [31], 기능성에 대한 연 구로는 지사, 이뇨, 해열, 항균, 항당뇨 및 항산화효과 등이 보 고되어 있다 [32-42]. 최근에는 식물 호르몬인 dihydrophaseic acid의 분리 동정, 돌연변이원성에 대한 효과, 우울증의 치료 에 대한 효과, 세포독성 보호효과, 심장질환 치료효과 및 항 경련효과 [43-48] 등이 보고되었으나 미백효과에 대해서는 거의 보고된 바가 없다.
현재까지 연자육에 대한 성분연구로는 주로 alkaloid 성분 에 대한 연구가 집중적으로 행해졌으며 nuciferine, N-nornu- ciferine, O-nornuciferine, roemerine 등의 aporphine계 alkaloid [49]와 liensinine, isoliensinine, neferine 등의 phenolic alkaloid [50] 및 (+)-1(R)-coclaurine, (-)-1(S)-norcoclaurine 등 benzyli- soquinoline alkaloid [51] 성분들이 보고되고 있다. 그밖에 (+)- catechin, (+)-gallocatechin, quercetin, kaempferol 등의 flavo- noid 화합물 [52] 및 tryptophan, asparagin, tyrosine 등 amino acid 성분들이 보고되어 있다.
본 연구에서는 연자육을 화장품의 새로운 미백소재로 이 용하기 위해 in vitro 상의 tyrosinase 활성 억제효과와 B16-F1 melanoma cell에서의 멜라닌 합성 억제효과를 측정하여 미 백효과를 평가하였고, 그 작용기전을 확인하기 위해 멜라닌 합성 과정에 관여하는 단백질의 발현과 N-glycosylation 저해 효과 등을 측정하였다.
2. 재료 및 방법
2.1. 실험재료, 시약 및 기기
본 실험에 사용한 연자육은 국내산으로 경기도 파주의 (주) 지유본초로부터 구입하여 사용하였다. 연자육의 추출과정 및 분획에 사용된 용매들은 시약급을 사용하였다. 세포배양 에 필요한 배지 및 시약은 Hyclone (USA), Sigma (USA)로부 터 구입하여 사용하였다. 흡광도 측정은 BIO-TEK Instruments (USA)의 EL800 microplate reader를 사용하였다. Western blot 을 위해 Bio-rad (USA)의 western blot kit 및 semi-dry transfer system을 사용하였고 image analysis system (Bio-rad, USA) 으로 결과를 확인하였다. 항체는 Santa Cruz Biotech (USA), Cell Signaling Technology (USA), Abcam Biochemicals (UK) 로부터 구입하였다.
2.2. 연자육 추출물 및 분획물의 제조
건조 연자육 100 g에 75% 에탄올수용액 500 mL을 가하여 60~90oC에서 4 h 동안 가열추출하고 여과하였다. 이 추출 과 정을 2회 반복하고 여과한 다음 추출여액을 회전식 증발 건
조기를 이용하여 40oC로 감압 농축하여 연자육 조추출물 (19.35 g)을 얻었다. 이것을 20% 메탄올수용액으로 현탁시킨 후 500 mL의 CH2Cl2를 가하여 분획한 뒤 얻어진 상층을 농 축하여 CH2Cl2 분획물 (NGC) 0.21 g을 얻었으며, 하층은 n- BuOH (500 mL)를 가하여 분획한 뒤 얻어진 상층을 농축하 여 n-BuOH 분획물 (NGB) 1.80 g을 얻었다.
2.3. Tyrosinase 저해활성 측정
Tyrosinase에 대한 활성은 각 농도별 시료 0.9 mL, 0.1 M 인산 완충액 (pH 6.8) 1.0 mL, 1.5 mM L-tyrosine 용액 1.0 mL을 넣 은 후, 37oC에서 10 min 간 반응시킨 뒤 mushroom tyrosinase (1,500 units/mL) 0.1 mL를 첨가하여 37oC에서 10 min 간 반 응시킨 후 UV-Vis spectrophotometer를 사용하여 475 nm에 서 흡광도를 측정하였다. Tyrosinase 저해활성은 다음 식에 따라 시료용액의 첨가군과 무첨가군의 흡광도 감소율로 나 타내었다.
2.4. 세포주 및 세포배양
세포주는 마우스 흑색종 세포주인 B16-F1 melanoma cell을 ATCC (American Type Culture Collection)에서 분양받아 사 용하였으며, 세포배양에 사용된 배지 (Dulbecco's modified Eagle's medium; DMEM)는 10% fetal bovine serum (HyClone Lab., USA), 1% antibiotic antimycotic (100 U/mL penicillin and 50µg/mL streptomycin, Life Tech Inc., USA)을 혼합한 배지를 사용하여 37oC, 5% CO2 incubator에서 배양하였다.
실험과정의 세포는 70~80%의 단층배양에서 실시하였다.
2.5. 세포 생존력 시험
본 실험에서 B16-F1 melanoma cell에 대한 시료의 처리농도 를 결정하기 위해 MTT [3-(4,5-dimethythiazol-2-yl)-2,5-di- phenytetrazolium bromide, Sigma] assay를 Mosmann의 방법 [53]을 변형하여 실시하였다. 이 분석법은 노란색의 수용성 기질인 MTT를 진청색의 비수용성 formazan으로 변환시키 는 살아있는 세포의 mitochondria dehydrogenase의 능력을 이 용한 방법이다. 생성된 formazan의 양은 살아있는 세포 수에 비례한다.
B16-F1 melanoma cell을 5×103 cells/well의 농도로 96-well plate에 분주하여 37oC, 5% CO2 조건하에서 24 h 동안 배양 하였다. 배양 후 배양액을 제거하고 시료를 농도별로 배지에 희석하여 교체한 후, 최종 200 nM α-melanocyte stimulating hormone (α-MSH)가 되도록 첨가하여 48 h 동안 더 배양하였 다. 배양 후 MTT assay를 통하여 세포 생존율을 확인하였다.
대조군은 시료를 처리하지 않은 배양액으로 설정한 후 흡 광도를 측정하였다. 세포의 생존능력은 다음의 식에 따라 계 산하였다.
2.6. 멜라닌 함량 측정
멜라닌 함량 측정은 Oka 등의 방법 [54]을 변형하여 사용하 였다. B16-F1 melanoma cell을 6-well plate에 1×105 cells/well 이 되게 준비한 한 후, 24 h 동안 37oC, CO2 incubator에서 배 양하였다. 배양액을 제거하고 시료를 농도별로 배지에 희석 하여 교체한 후, 최종 200 nM α-MSH가 되도록 첨가하여 48 h 동안 더 배양하였다. 대조군은 α-MSH만 첨가한 것으로 하 였으며 양성 대조군으로는 알부틴을 사용하였다. 배양 후 배 양액을 제거하고 PBS로 세척한 후, 10% DMSO가 함유된 1 N NaOH를 첨가한 후 50oC 항온조에서 세포 내 멜라닌을 용 해시켰다. 이 액을 ELISA reader를 이용하여 490 nm에서 흡 광도를 측정하였으며, 총 단백질량으로 보정하였다.
2.7. 단백질 발현 조사 (Western Blot 분석)
B16-F1 melanoma cell을 10% FBS를 첨가한 DMEM에 1×
106 cells의 밀도로 100 mm dish에 분주하여 37oC, 5% CO2 조 건하에서 24 h 동안 배양하였다. 배양액을 제거하고 시료를 농도별로 배지에 희석하여 교체한 후, 최종 200 nM α-MSH 가 되도록 첨가하여 48 h 동안 더 배양하였다. 배양한 세포를 PBS로 세척하여 1.5 mL microtube에 옮기고 세포침전물을 Pro-prep protein extraction solution (Intron Biotechnology, Korea)을 첨가하여 세포를 파괴한 후 4oC에서 15,000 rpm으 로 20 min 동안 원심분리하고 상층액을 취하여 단백질을 분 리하였다. 분리한 단백질은 BCA 방법에 따라 정량한 후 12% SDS-ployacrylamide gel을 사용하여 전기영동한 다음, PVDF membrane에 transfer시켰다. Membrane을 5% skim milk solution으로 1 h 동안 blocking한 뒤 primary antibody로 상온 에서 3~4 h 동안 반응시키고, horseradish peroxidase가 결합된 secondary antibody로 처리하였다. Chemiluminescence kit을 이용하여 발색된 밴드의 강도를 확인하였다. β-actin은 각각 의 시료에 동양의 단백질이 들어있는지 확인하기 위한 대조 군으로 사용하였다.
2.8. α-Glucosidase 저해활성 측정
α-Glucosidase가 ρ-nitrophenyl (ρNP) glycoside의 glycoside 부분을 기질로 인식하여, ρNP와 glycoside를 효소반응으로 끊어주고 여기에서 끊어져 나온 ρNP의 양을 405 nm에서 흡 광도를 측정하여 이것으로 α-glucosidase activity를 간접적으 로 측정하였다 [55]. 즉, 0.1 M phosphate buffer (pH 6.8)에 시 료와 α-glucosidase (1 unit/mL)를 넣고 37oC에서 5 min 간 반 응시켰다. 여기에 2 mM 4-Nitrophenyl α-D-glucopyranoside 를 가해 37oC에서 10 min 간 효소반응을 시킨 후 1 M Na2CO3 를 넣어 반응을 정지시키고 생성된 ρNP의 양을 405 nm에서 흡광도를 측정하여 결정하였다.
2.9. N-Glycosylation 억제효과 측정
B16-F1 melanoma cell을 10% FBS를 첨가한 DMEM에 1×106 cells의 밀도로 100 mm dish에 분주하여 37oC, 5% CO2 조건 하에서 24 h 동안 배양하였다. 배양액을 제거하고 시료를 농
저해율 (%) = 시료 첨가군의 흡광도 ×100
무첨가군의 흡광도
세포 생존율 (%) =시료 첨가군의 흡광도−대조군의 흡광도×100
대조군의 흡광도
도별로 배지에 희석하여 교체한 후, 최종 200 nM α-MSH가 되도록 첨가하여 48 h 동안 더 배양하였다. Western blot 실험 의 단백질 분리법과 같은 방법으로 단백질을 분리하고 정량 한 후 100 µg의 단백질을 denaturing buffer와 섞어 100oC에서 10 min 동안 반응시킨 후 Endo H 효소 (New England Bio- Labs, USA)와 반응 버퍼를 첨가하여 37oC에서 1 h 동안 반응 시켰다. 반응물은 western blot을 수행하여 fully glycosylated protein과 naked protein을 image analysis system으로 분석하 였다.
2.10. 통계처리
모든 실험 결과는 평균±표준편차로 표기하였고 통계적 유의 성은 SPSS를 이용한 Student's t-test를 시행하여 p-value를 구 하였으며, p<0.05인 경우 *, p<0.01인 경우 **로 유의성이 있 다고 표시하였다.
3. 결과 및 고찰 3.1. 세포독성
NGB와 NGC의 실험에 사용할 농도범위를 결정하기 위해 MTT assay를 시행하였다. 세포 수준의 연구에 많이 이용되고 있는 MTT assay는 cell proliferation과 viability의 in vitro 분 석에 매우 유용하게 사용되고 있다 [53]. NGB와 NGC에 의 한 B16-F1 melanoma cell의 생존율을 확인한 결과 (Fig. 1), NGB와 NGC 모두 100 µg/mL 이하의 농도로 처리시 세포생 존율이 80% 이상으로 나타났지만 200 µg/mL 이상의 농도로 처리시 둘 다 세포독성으로 인해 60% 이하로 급격히 생존율
이 저하되었다. 따라서 100 µg/mL 농도 이하에서 멜라닌 합 성 저해 및 단백질 발현 실험을 진행하였다.
3.2. 멜라닌 합성 억제효과
우선 in vitro 상에서 mushroom tyrosinase를 이용하여 tyrosi- nase 활성에 미치는 영향을 측정한 결과, NGB와 NGC는 mu- shroom tyrosinase에 대한 저해 활성을 나타내지 않았다 (data not shown). 다음으로 멜라닌 합성에 미치는 영향을 확인하 기 위해 B16-F1 melanoma cell에 NGB와 NGC를 각각 0, 20, 50, 100µg/mL의 농도로 처리하고 48 h이 지난 후 멜라닌 함 량을 측정한 결과, 농도 의존적으로 멜라닌 합성이 저해됨을 확인하였다. Fig. 2에서처럼 최종 농도인 100 µg/mL에서 NGB 는 46%, NGC는 53%의 멜라닌 합성 억제효과를 나타내었다.
이는 100 µg/mL의 농도에서 45%의 멜라닌 합성 억제효과를 나타낸 양성대조군으로 사용한 알부틴과 유사한 정도의 효 과였다. 이러한 결과로 보아 NGB와 NGC는 tyrosinase의 활 성을 직접적으로 저해하지 않고 멜라닌 합성 신호전달 경로 에 작용할 것으로 추정된다. 따라서 NGB와 NGC에 의한 멜 라닌 합성 관련 단백질의 발현 양상을 분석하였다.
3.3. Tyrosinase/TRP-1/TRP-2 단백질 발현 저해효과 본 실험에서는 NGB와 NGC의 멜라닌 합성 저해효과가 tyro- sinase, TRP-1 및 TRP-2와 같은 멜라닌 합성 효소의 발현 억 제와 관련된 영향인지 확인하기 위하여 western blot assay를 수행하였다. 즉, 100 µg/mL 농도의 NGB와 NGC를 B16-F1 melanoma cell에 처리한 후 멜라닌 합성에 관여하는 tyrosi- nase, TRP-1, TRP-2의 단백질 발현량을 측정하였다. 이때 세
Fig. 1. The effect of BuOH layer of 75% EtOH extract of the Ne- lumbinis nucifera Gaertner (NGB) and CH2Cl2 layer of 75% EtOH extract of the Nelumbinis nucifera Gaertner (NGC) on cell viabi- lity in B16-F1 melanoma cells by MTT assay. The cells were trea- ted with various concentration of NGB and NGC for 48 h. The cell viability was measured by the MTT method. The results were ex- pressed as the mean±S.D. from the three independent experiments.
*p<0.05 and **p<0.01 vs. control group treated with α-MSH.
Fig. 2. Inhibitory effect of BuOH layer of 75% EtOH extract of the Nelumbinis nucifera Gaertner (NGB) and CH2Cl2 layer of 75%
EtOH extract of the Nelumbinis nucifera Gaertner (NGC) on mela- nin synthesis in B16-F1 melanoma cells. The cells were treated with NGB and NGC at indicated concentration for 48 h. The ab- sorbance was measured at 490 nm and corresponding total protein content was used to normalize absorbance. The results were expre- ssed as the mean±S.D. from the three independent experiments.
**p<0.01 vs. control group treated with α-MSH.
포의 여러 조건에서도 그 발현 정도의 차이가 거의 없는 housekeeping gene인 β-actin을 대조군으로 사용하였다. 그 결 과 Fig. 3 (a)에서와 같이 NGB와 NGC를 처리한 군에서의 ty- rosinase 발현이 처리하지 않은 군보다 감소하였다. 즉, NGB 를 처리한 군에서는 35%, NGC를 처리한 군에서는 63%의 tyrosinase 발현 저해효과를 확인하였다. 같은 농도의 알부틴 을 처리한 군의 tyrosinase 발현 저해율은 2% 정도로, tyrosi- nase 단백질의 발현에는 거의 영향을 미치지 않는 것으로 확 인되었다. 또한 Fig. 3의 (b), (c)에서와 같이 TRP-1과 TRP-2 의 경우, NGB를 처리한 군에서는 저해효과가 나타나지 않 았으며 NGC를 처리한 군에서는 TRP-1과 TRP-2의 발현도 저해되었는데, TRP-1은 72%, TRP-2는 15%가 저해됨을 확 인하였다. 알부틴을 처리한 경우, TRP-1의 발현은 약 20% 저 해하였으며, TRP-2에는 영향을 미치지 않는 것으로 확인되 었다. 이상의 결과, NGB는 멜라닌 합성에 직접적으로 관여 하는 효소인 tyrosinase의 발현을 저해함으로써, NGC는 tyro- sinase, TRP-1 및 TRP-2의 발현을 모두 저해함으로써 멜라닌 합성을 저해하는 것임을 확인할 수 있었다. Nakamura 등 [56]
은 연꽃봉오리와 연잎의 methanol 추출물이 강력한 melano- genesis 저해효과를 나타내었으며 이는 연꽃봉오리와 연잎 의 methanol 추출물로부터 분리한 alkaloid 류 중 nuciferine과 N-methylasimilobine이 tyrosinase, TRP-1 및 TRP-2의 mRNA 발현을 저해함에 따른 것으로 보고한 바 있다. 따라서 NGB 및 NGC의 멜라닌 합성에 관여하는 단백질 즉, tyrosinase, TRP-1 및 TRP-2의 발현 저해는 연자육에 함유된 nuciferine 과 같은 alkaloid 성분들에 의한 것으로 추정된다.
3.4. cAMP / PKA 경로에 관련된 신호전달 단백질 발현에 미치는 효과
세포내 멜라닌 합성 신호전달 경로 중 주요 경로인 cAMP/
PKA 경로는 α-MSH에 의해 유도된 cAMP 신호가 하류 신호 전달 물질인 PKA의 활성화를 유도하며, 세포내 CREB를 활 성화시킴으로써 MITF의 발현을 증가시키고 MITF는 tyrosi- nase와 TRPs의 합성을 증가시켜 멜라닌 합성을 촉진시키는
것으로 알려져 있다 [5-7]. 따라서 본 연구에서는 NGB와 NGC의 멜라닌 합성 억제효과가 이러한 신호전달 과정과 연 관이 있는지 확인하기 위하여 α-MSH에 의해 멜라닌 합성이 Fig. 3. The effect of BuOH layer of 75% EtOH extract of the Nelumbinis nucifera Gaertner (NGB) and CH2Cl2 layer of 75% EtOH extract of the Nelumbinis nucifera Gaertner (NGC) on the melanogenic enzymes ((a) tyrosinase, (b) TRP-1, (c) TRP-2) expression in B16-F1 melanoma cells. The cells were treated with NGB, NGC, and arbutin at a concentration of 100µg/mL for 48 h. The results were expressed as the mean±S.D. from the three independent experiments. *p<0.05 and **p<0.01 vs. control group treated with α-MSH.
Fig. 4. The effect of BuOH layer of 75% EtOH extract of the Nelumbinis nucifera Gaertner (NGB) and CH2Cl2 layer of 75%
EtOH extract of the Nelumbinis nucifera Gaertner (NGC) on the CREB phosphorylation (a) and MITF expression (b) in B16-F1 melanoma cells. The cells were treated with NGB, NGC, and ar- butin at a concentration of 100µg/mL for 48 h. The results were expressed as the mean±S.D. from the three independent experi- ments. **p<0.01 vs. control group treated with α-MSH.
활성화된 세포에 NGB와 NGC를 처리 후 이들 신호전달 과 정에 관여하는 주요 단백질인 p-CREB과 MITF의 발현량을 western blot assay를 통하여 측정하였다. Fig. 4에서와 같이 p-CREB의 경우 NGC를 처리한 군에서만 46%의 저해효과를 나타내었고, MITF의 경우 NGB와 NGC 처리군 모두에서 저 해효과를 확인할 수 있었다. 즉, NGB를 처리한 군은 37%, NGC를 처리한 군은 62%의 MITF 발현 저해율을 나타내었다.
이러한 결과는 NGB와 NGC가 미백에 관련된 전사인자의 발 현에도 효과가 있음을 나타내며, 특히 NGC의 경우 cAMP/
PKA 경로에 영향을 미침으로써 전사인자인 MITF 발현과 상위 신호전달 인자인 CREB의 활성화를 조절함으로써 tyro- sinase 및 TRPs (TRP-1, TRP-2)의 발현을 저해하고 결과적으 로 멜라닌 합성을 억제하는 것으로 사료된다.
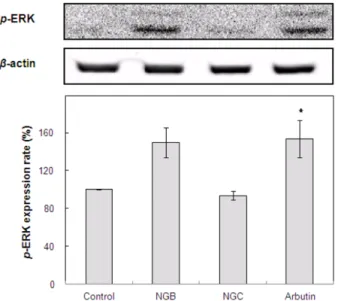
3.5. ERK pathway activation에 미치는 효과
주로 세포의 증식, 분화에 관여하는 것으로 알려진 MAPK 경 로에 속하는 ERK 신호는 MITF의 인산화를 유도하여 MITF 의 ubiquitination이 이루어져 proteosomal degradation을 일으 키게 됨으로써 멜라닌 합성을 감소시키는 것으로 알려져 있 다 [13,57]. NGB와 NGC가 이러한 ERK pathway 활성에 영 향을 미치는지 확인하고자 B16-F1 melanoma cell에 NGB와 NGC를 처리한 후 p-ERK의 발현을 측정하였으며 그 결과는 Fig. 5와 같다. NGB를 처리한 경우 ERK가 활성화되어 p-ERK 가 증가되었으나, NGC를 처리한 경우에는 ERK의 활성화에 영향을 미치지 않는 것으로 확인되었다. 이러한 결과로 보아 NGB의 멜라닌 합성 저해효과는 ERK pathway 활성화를 통
해 MITF의 분해를 촉진함으로써 MITF 발현이 억제되었고, 그 결과 tyrosinase 발현 역시 억제되었으며 최종적으로 멜라 닌 합성이 억제된 것으로 사료된다.
3.6. α-Glucosidase 억제효과
본 실험에서는 NGB와 NGC의 멜라닌 합성 저해효과가 tyro- sinase의 glycosylation 과정을 저해하는 것과 관련이 있는지 확인하기 위하여 in vitro 상에서 α-glucosidase 활성 억제효과 를 측정하였다. α-glucosidase에 ρNP glycoside를 넣고 효소 반응을 시킨 결과는 Table 1과 같다. 즉, α-glucosidase를 50%
억제하는 농도인 IC50이 NGB는 1.1±0.1 µg/mL, NGC는 14.6
±1.0 µg/mL로 468.6±16.1 µg/mL의 IC50치를 나타낸 positive control인 acarbose에 비해 월등히 높은 저해 효과를 나타내었 다. Choi 등 [58]은 최근 α-glucosidase 저해제인 deoxynojiri- mycin (DNJ)이 tyrosinase의 glycosylation을 차단하여 tyrosi- nase의 멜라노좀으로의 이동을 억제하였다고 보고하였으며, Mishima와 Imokawa [59]도 glucosamine이나 tunicamycin과 같은 α-glucosidase 저해제의 tyrosinase glycosylation 억제효 과에 대해 보고한 바 있다. 이와 같이 본 실험에서도 NGB와 NGC가 α-glucosidase 활성을 저해하였으며, 이로써 tyrosi- nase의 glycosylation을 억제할 가능성이 있는 것으로 판단되 어 이후 세포 수준에서의 N-glycosylation 저해효과를 측정하 였다.
3.7. N-Glycosylation 저해효과
Tyrosinase는 N-linked complex oligosaccharide가 붙어 있는 당단백질로서 Endo H 라는 효소에 의해 가수분해 되지 않는 기질특이성을 가진다. 어떠한 요인에 의해 당단백질을 만드 는 glycosylation 과정에 문제가 생겨 tyrosinase의 당 부분이 완전한 complex oligosaccharide를 형성하지 못하면 Endo H 에 의해서 가수분해되어 약 51 kD으로 작아지게 된다 [58, 60]. 이러한 특징을 이용하면 NGB와 NGC가 실질적으로 세 포 내에 작용하여 tyrosinase의 glycosylation을 억제하였는지 확인할 수 있다. 본 실험에서는 100 µg/mL 농도의 NGB와 NGC를 B16-F1 melanoma cell에 48 h 처리 후 단백질을 모아 Endo H로 1 h 처리하고 western blot assay를 실시하여 NGB 와 NGC가 tyrosinase의 glycosylation을 저해하였는지 관찰하 였다. 이 때, 양성대조군으로는 NB-DNJ를 사용하였다. Fig.
6에서 보는 바와 같이 tyrosinase의 경우 시료를 처리하지 않 은 대조군 (lane 1)은 64 kD에 진한 tyrosinase 밴드가 보이는
Fig. 5. The effect of BuOH layer of 75% EtOH extract of the Nelumbinis nucifera Gaertner (NGB) and CH2Cl2 layer of 75%
EtOH extract of the Nelumbinis nucifera Gaertner (NGC) on the ERK activation in B16-F1 melanoma cells. The cells were treated with NGB, NGC, and arbutin at a concentration of 100µg/mL for 48 h. The results were expressed as the mean±S.D. from the three independent experiments. *p<0.05 vs. control group treated with α-MSH.
Table 1. α-Glucosidase inhibitory effect of NGB and NGC
Materials IC50 (µg/mL)
NGB 1.1±0.1**
NGC 14.6±1.0**
Acarbose 468.6±16.1
aNB-DNJ 569.8±8.5**
The results are shown as the mean
±S.E. of triplicate measurements performed on separate experiments. **p<0.01, compared with positive control (arcarbose).
a
NB-DNJ: N-butyl-deoxynojirimycin.
반면, 양성대조군인 NB-DNJ 처리군 (lane 4)에서는 64 kD의 밴드가 없어지고 51 kD을 포함한 64 kD보다 작은 분자량의 밴드들을 확인할 수 있었다. 이는 tyrosinase의 glycosylation 이 저해되어 Endo H에 의해 가수분해된 결과로, NGB, NGC 처리군 (lane 2, lane 3)의 경우에도 64 kD의 밴드가 줄어들고 51 kD의 밴드가 진해진 것을 확인할 수 있었다. TRP-1의 경 우 양성대조군인 NB-DNJ 처리군 (lane 4)은 64 kD의 TRP-1 밴드가 사라지고 그 아래쪽의 51 kD를 포함한 작은 밴드들이 생성되었는데 이 역시 Endo H에 의해 가수분해 된 결과이다.
그러나 대조군과 NGB, NGC를 처리한 군에서는 이러한 효과 를 확인할 수 없었다. TRP-2의 경우에는 NB-DNJ, NGB, NGC 모두 영향을 미치지 않는 것으로 확인되었다. 이상의 실험 결 과, NGB와 NGC는 B16-F1 melanoma cell에서 α-glucosidase 를 억제하여 tyrosinase의 glycosylation을 저해하는 것으로 사 료되며 이로 인해 tyrosinase 발현 저해 및 멜라닌 합성 억제 효과에 영향을 미친 것으로 생각되어진다.
4. 결론
본 연구에서는 연자육을 화장품의 새로운 미백소재로 이용 하고자 연자육 에탄올추출물의 CH2Cl2 분획물인 NGC와 n- BuOH 분획물인 NGB를 얻어 이들의 미백활성을 측정하고, 그 작용기전에 대해 확인하였다. NGC와 NGB는 B16-F1 melanoma cell을 이용한 멜라닌 합성 억제효과 측정 결과, 미 백소재로 알려져 있는 알부틴과 유사한 정도의 미백 활성을 가지는 것으로 확인되었다. 그러나 in vitro 상의 mushroom
tyrosinase의 활성에는 영향을 미치지 않는 것으로 나타나 tyrosinase의 활성을 직접적으로 저해하지 않고 멜라닌 합성 신호전달 경로에 작용할 것으로 추정되어 NGC와 NGB에 의 한 멜라닌 합성 관련 단백질의 발현 양상을 분석하였다. 먼저 멜라닌 합성 경로에 작용하는 효소들 즉, tyrosinase와 TRP- 1, TRP-2의 발현 정도를 측정한 결과 NGC는 tyrosinase, TRP- 1, TRP-2의 발현을 모두 억제하였으며, NGB는 tyrosinase의 발현만을 억제하였다. 다음으로 NGC와 NGB의 멜라닌 합성 억제효과가 cAMP / PKA 경로에 작용한 결과인지를 확인하 고자 p-CREB와 MITF의 발현 정도를 측정한 결과 NGC는 p- CREB과 MITF의 발현을 모두 억제하였으며, NGB는 MITF 의 발현만을 억제하는 것으로 확인되었다. 멜라닌 합성 기전 중 ERK pathway 활성화에 미치는 영향을 측정한 결과, NGB 가 p-ERK의 발현을 상당히 증가시키는 것을 확인할 수 있었 다. 또한 tyrosinase glycosylation 저해효과를 확인하고자 먼 저 in vitro 상에서 α-glucosidase 저해효과를 측정한 결과 NGC와 NGB 모두 상당한 정도의 저해 활성을 나타내었다.
따라서 세포수준에서 Endo H 처리를 통해 tyrosinase의 gly- cosylation을 저해하는지 여부를 확인한 결과 NGB와 NGC 모 두 tyrosinase의 glycosylation을 저해하는 것으로 확인되었다.
결과를 종합해보면 NGC는 멜라닌 합성의 신호전달 경로 중 cAMP / PKA 경로를 저해함으로써 전사인자인 p-CREB와 MITF의 발현을 저해하고, 그 결과 tyrosinase와 TRP-1, TRP- 2의 발현을 저해함으로써 멜라닌 합성을 억제하는 기전과 tyrosinase의 N-glycosylation 과정을 저해함으로써 tyrosinase 활성을 억제하는 기전이 복합적으로 작용하여 멜라닌 합성 이 저해되는 것으로 판단된다. 또한 NGB는 ERK pathway의 활성화를 통해 MITF의 분해를 촉진시키고 이로 인해 tyrosi- nase의 발현을 감소시키는 기전과 역시 tyrosinase의 N-glyco- sylation 과정을 저해함으로써 tyrosinase 활성을 억제하는 2가 지 기전이 복합적으로 작용함으로써 멜라닌 합성을 저해하는 것으로 사료된다. 향후 활성 물질에 대한 분석과 인체실험을 통한 안정성 및 미백효능의 검증이 필요하나 연자육은 화장품 의 새로운 미백 소재로서의 활용가치가 큰 것으로 판단된다.
감사
본 연구는 2011년 중소기업기술혁신개발사업 (과제번호 SA 112778)의 연구비 일부에 의하여 이루어진 것으로 지원에 감 사드립니다.
REFERENCES
1. Hill, H. Z., W. Li, P. Xin, and D. L. Michell (1998) Melanin: A two edged sword? Pigment Cell Res. 10: 158-161.
2. Seiberg, M., L. Babiarz, and C. B. Lin (2003) IL-41 the PAR-2 pathway is differentially expressed in skin of color. Pigment Cell Res. 16: 591-591.
Fig. 6. The effect of BuOH layer of 75% EtOH extract of the Nelumbinis nucifera Gaertner (NGB) and CH2Cl2 layer of 75%
EtOH extract of the Nelumbinis nucifera Gaertner (NGC) on the N-glycosylation processing of tyrosinase, TRP-1, and TRP-2 in B16-F1 melanoma cells. Lane 1, Control; lane 2, NGB 100µg/
mL; lane 3, NGC 100µg/mL; lane 4, NB-DNJ 100 µg/mL treated with α-MSH.
3. Busca, R. and R. Ballotti (2000) Cyclic AMP a key messenger in the regulation of skin pigmentation. Pigment Cell Res. 13: 60-69.
4. Sassone, C. P. (1998) Coupling gene expression to cAMP signal- ling: Role of CREB and CREM. Int. J. Biochem. Cell B. 30: 27- 38.
5. Bertolotto, C., R. Busca, P. Abbe, K. Bille, E. Aberdam, J. P. Or- tonne, and R. Ballotti (1998) Different cis-acting elements are in- volved in the regulation of TRP1 and TRP2 promoter activities by cyclic AMP: pivotal role of M boxes (GTCATGTGCT) and of mi- crophthalmia. Mol. Cell. Biol. 18: 694-702.
6. Fuller, B. B., J. B. Lunsford, and D. S. Iman (1987) Alpha-melano- cyte-stimulating hormone regulation of tyrosinase in Cloudman S- 91 mouse melanoma cell cultures. J. Biol. Chem. 262: 4024-4033.
7. Yokoyama, K., H. Suzki, K. Yasumoto, Y. Tomita, and S. Shiba- hara (1994) Molecular cloning and functional analysis of a cDNA coding for human DOPAchrome tautomerase/tyrosinase-related protein-2. Biochim. Biophys. Acta. 1217: 317-321.
8. Bertolotto, C., P. Abbe, T. J. Hemesath, K. Bille, D. E. Fisher, J. P.
Ortonne, and R. Ballotti (1998) Microphthalmia gene product as a signal transducer in cAMP-induced differentiation of melano- cytes. J. Cell Biol. 142: 827-835.
9. Levy, C., M. Khaled, and D. E. Fisher (2006) MITF: Master regu- lator of melanocyte development and melanoma oncogene. Trends Mol. Med. 12: 406-414 .
10. Yasumoto, K., K. Yokoyama, K. Takahashi, Y. Tomita, and S.
Shibahara (1997) Functional analysis of microphthalmia-associated transcription factor in pigment cell-specific transcription of the hu- man tyrosinase family genes. J. Biol. Chem. 272: 503-509.
11. Yavuzer, U., E. Keenan, P. Lowings, J. Vachtenheim, G. Currie, and C. R. Goding (1995) The microphthalmia gene product interacts with the retinoblastoma protein in vitro and is a target for deregula- tion of melanocyte-specific transcription. Oncogene 10: 123-134.
12. Englaro, W., C. Bertolotto, R Busca, A. Brunet, G. Pages, J. P. Or- tonne, and R. Ballotti (1998) Inhibition of the mitogen-activated protein kinase pathway triggers B16 melanoma cell differentiation.
J. Biol. Chem. 273: 9966-9970.
13. Kim, D. S., E. S. Hwang, J. E. Lee, S. Y. Kim, S. B. Kwon, and K.
C. Park (2003) Sphingosine-1-phosphate decreases melanin syn- thesis via sustained ERK activation and subsequent MITF degra- dation. J. Cell Sci. 116: 1699-1706.
14. Kim, D. S., S. Y. Kim, J. H. Chung, K. H. Kim, H. C. Eun, and K.
C. Park (2002) Delayed ERK activation by ceramide reduces mel- anin synthesis in human melanocytes. Cell. Signal. 14: 779-785.
15. Englaro, W., R. Rezzonico, M. Durand-Clement, D. Lallemand, J.
P. Ortonne, and R. Ballotti (1995) Mitogen-activated protein kinase pathway and AP-1 are activated during cAMP-induced melano- genesis in B-16 melanoma cells. J. Biol. Chem. 270: 24315-24320.
16. Kobayashi, T., W. D. Vieira, B. Potterf, C. Sakai, G. Imokawa, and V. J. Hearing (1995) Modulation of melanogenic protein expres- sion during the switch from euto pheomelanogenesis. J. Cell Sci.
108: 2301-2309.
17. Kobayashi, T., K. Urabe, A. Winder, C. Jimenez-Cervantes, G. Imo- kawa, T. Brewington, F. Solano, J. C. Garcia-Borron, and V. J.
Hearing (1994) Tyrosinase related protein 1 (TRP1) functions as a DHICA oxidase in melanin biosynthesis. EMBO J. 13: 5818-5825.
18. Branza-Nichita, N., G. Negroiu, A. J. Petrescu, E. F. Garman, F. M.
Platt, M. R. Wormald, R. A. Dwek, and S. M. Petrescu (2000).
Mutations at critical N-glycosylation sites reduce tyrosinase activ- ity by altering folding and quality control. J. Biol. Chem. 275:
8169-8175.
19. Goochee, C. F. (1992) Bioprocess factors affecting glycoprotein oligosaccharide structure. Dev. Biol. Stand. 76: 95-104.
20. Petrescu, A. J., T. D. Butters, G. Reinkensmeier, S. Petrescu, F. M.
Platt, R. A. Dwek, and M. R. Wormal (1997) The solution NMR structure of glucosylated N-glycans involved in the early stages of glycoprotein biosynthesis and folding. EMBO J. 16: 4302-4310.
21. Kornfeld, R. and S. Kornfeld (1985) Assembly of asparagine-linked oligosaccharides. Annu. Rev. Biochem. 54: 631-664.
22. Winchester, B. and G. W. Fleet (1992) Amino-sugar glycosidase inhibitors: Versatile tools for glycobiologists. Glycobiology 2: 199- 210.
23. Petrescu, S. M., A. J. Petrescu, H. N. Titu, R. A. Dwek, and F. M.
Platt (1997) Inhibition of N-glycan processing in B16 melanoma cells results in inactivation of tyrosinase but does not prevent its transport to the melanosome. J. Biol. Chem. 272: 15796-15803.
24. Branza-Nichita. N., A. J. Petrescu, R. A. Dwek, M. R. Wormald, F.
M. Platt, and S. M. Petrescu (1999) Tyrosinase folding and copper loading in vivo: a crucial role for calnexin and alpha-glucosidase II. Biochem. Biophys. Res. Commun. 261: 720-725.
25. Wang, Y. and M. J. Androlewicz (2000) Oligosaccharide trimming plays a role in the endoplasmic reticulum-associated degradation of tyrosinase. Biochem. Biophys. Res. Commun. 271: 22-27.
26. Imokawa, G. and Y. Mishima (1984) Functional analysis of tyrosi- nase isozymes of cultured malignant melanoma cells during the re- covery period following interrupted melanogenesis induced by gly- cosylation inhibitors. J. Invest. Dermatol. 83: 196-201.
27. Franchi, J., M. C. Coutadeur, C. Marteau, M. Mersel, and A. Kup- ferberg (2000) Depigmenting effects of calcium D-pantetheine-S- sulfonate on human melanocytes. Pigment Cell Res. 13: 165-171.
28. Aoki, H. M., O. Ifuku, and A. S. Zervos (1998) Identification of a new microphthalmia(Mi)-interacting protein, rKr2, involved in the regulation of melanogenesis. IFSCC. 1998: 67-78
29. Seiberg, M., C. Paine, E. Sharlow, P. Andrade Gordon, M. Cos- tanzo, M. Eisinger, and S. S. Shapiro (2000) The protease-activated receptor 2 regulates pigmentation via keratinocyte-melanocyte inter- action. Exp. Cell Res. 254: 25-32.
30. Negroiu, G., N. Branza-Nichita, A. J. Petrescu, R. A. Dwek, and S.
M. Petrescu (1999) Protein specific N-glycosylation of tyrosinase and tyrosinase-related protein-1 in B16 mouse melanoma cells.
Biochem. J. 344: 659-665.
31. Bensky, D. and A. Gamble (1993) Chinese Herbal Medicine;
Materia Medica. 2nd ed., pp. 262. Eastland press, Seattle, WA, USA.
32. Mukheriee, P. K., J. Das, R. Balasubramanian, K. Saha, M. Pal, and B. Saha (1995) Antidiarrhoeal evaluation of Nelumbo nucifera rhizome extract. Indian J. Pharm. 27: 262-264.
33. Mukheriee, P. K., J. Das, K. Saha, M. Pal, and B. P. Saha (1996) Diuretic activity of the rhizomes of Nelumbo nucifera Gaertn. Phy- tother. Res. 10: 424-425.
34. Mukheriee, P. K., K. Saha, J. Das, M. Pal, and B. P. Saha (1996)
Antipyretic activity of Nelumbo nucifera rhizome extract. Indian J.
Exp. Biol. 34, 275-276.
35. Mukheriee, P. K. (2002) Quality control of herbal drugs-an approach to evaluation of botanicals, 604, Business Horizons, New Delhi, India.
36. Mukheriee, P. K., S. N. Giri, K. Saha, M. Pal, and B. P. Saha (1995) Antifungal screening of Nelumbo nucifera (Nymphaeaceae) rhi- zome extract. Indian J. Microbiol. 35: 327-330.
37. Mukheriee, P. K., R. Balasubramanian, K. Saha, M. Pal, and B. P.
Saha (1995) Antibacterial efficiency of Nelumbo nucifera (Nym- phaeaceae) rhizome extract. Indian Drugs 32: 274-276.
38. Mukheriee, P. K., S. R. Pal, K. Saha, and B. P. Saha (1995) Hypo- glycemic activity of Nelumbo nucifera rhizome (methanolic extract) in streptozotocin induced diabetic rats. Phytother. Res. 9: 522-524.
39. Cho, E. J., T. Yokozawa, D. Y. Rhyu, S. C. Kim, N. Shibahara, and J. C. Park (2003) Study on the inhibitory effects of Korean medici- nal plants and their main compounds on the 1,1-diphenyl-2-picryl- hydrazyl radical. Phytomedicine 10: 544-551.
40. Hu, M. and L. H. Skibsted (2002) Antioxidative capacity of rhi- zome extract and rhizome knot extract of edible lotus (Nelumbo nuficera). Food Chem. 76: 327-333.
41. Jung, H. A., J. E. Kim, H. Y. Chung, and J. S. Choi (2003) Antiox- idant principles of Nelumbo nucifera Stamens. Arch. Pharm. Res.
26: 279-285.
42. Park, J. H., D. W. Kim, B. G. Lee, and K. I. Byun (2010) Antioxi- dant activities and inhibitory effect on oxidative DNA damage of Nelumbinis semen extracts. Korean J. Herbol. 25: 55-59.
43. Seo, J. H., Y. H. Choi, M. Y. Yoo, K. S. Hong, B. H. Lee, G. H.
Yon, Y. S. Kim, Y. K. Kim, and S. Y. Ryu (2006) Isolation of dihy- drophaseic acid from seed extract of Nelumbo nucifera. Korean J.
Pharmacogn. 37: 290-293.
44. Song, G. S., B. Y. Ahn, K. S. Lee, L. K. Maeng, and D. S. Choi (1997) Effect of hot water extracts from medical plants on the mutagenicity of indirect mutagens. J. Food Sci. Technol. 29: 1288- 1294.
45. Lee, J. W., M. C. Hong, M. K. Shin, and H. S. Bae (2006) Com- parison of Nelumbinis semen extract with hypericum perforatum and fluoxetine in animal model of depression. Korean J. Ori. Med.
Physiol. Pathol. 20: 830-843.
46. Kim, H. G. and M. S. Oh (2009) Protective effects of Nelumbinis Semen against neurotoxicity induced by 6-hydroxydopamine in dopaminergic cells. Korean J. Herbol. 24: 87-92.
47. Ann, C. J., G. H. Lee, Y. S. Kim, M. C. Hong, H. S. Bae, J. H.
Kim, and M. G. Shin (2010) Effect of Nelumbinis semen on the recovery of the cardiac muscle activity by proteome analysis.
Korean J. Ori. Med. Physiol. Pathol. 24: 962-969.
48. Kim, S. H. and J. W. Choi (2011) Action mechanism of anticon- vulsive effect of Nelumbo Nucifera in pentylenetetrazole-induced animal models. Korean J. Ori. Med. Physiol. Pathol. 25: 614-619.
49. Luo, X. and S. Yao (2005) Stimultaneous analysis of N-nornuci- ferine, O-nornuciferine, nuciferine, and roemerine in leaves of Ne- lumbo nucifera Gaertn by high-performance liquid chromatogra- phy-photodiode array detection- electrospray mass spectrometry.
Analytica. Chimica. Acta. 538: 129-133.
50. Wu, S. and Y. Pan (2004) Prepatative counter-current chromatog- raphy isolation of liensinine and its analogues from embryo of the seed of Nelumbo nucifera Gaertn using upright coil planet centri- fuge with for multilayer coil connected in series. J. Chromatogr. A.
1041: 153-162.
51. Kashiwada, Y., A. Aoshima, Y. Ikeshiro, Y. P. Chen, H. Furukawa, M. Itoigawa, T. Fujioka, K. Mihashi, L. M. Cowedtino, S. L. Mor- ris-Natschke, and K. H. Lee (2005) Anti-HIV benzylisoquinoline alkaloids and flavonoids from the leaves of Nelumbo nucifera and structure-activity correlations with related alkaloids. Bioorg. Med.
Chem. 13: 443-448.
52. Kim, J. S., S. M. Cho, J. H. Kim, and M. W. Lee (2001) Phenolic compounds from the node of Lotus Rhizome. Yakhak Hoeji. 45:
599-603.
53. Mosmann, T. (1983) Rapid colorimetric assay for the cellular growth and survival: application to proliferation and cytotoxic assay. J. Im- mun. Methods 65: 55-63.
54. Oka, M., M. Ichihashi, and A. K. Chakraborty (1996) Enhanced expression of protein kinase C subspecies in melanogenic com- partments in B16 melanoma cells by UVB or MSH. J. Invest. Der- matol. 106: 377-378.
55. Angelov, A., M. Putyrski, and W. Liebl (2006) Molecular and bio- chemical characterization of alpha-glucosidase and alpha-man- nosidase and their clustered genes from the thermoacidophilic ar- chaeon Picrophilus torridus. J. Bacteriol. 188: 7123-7131.
56. Nakamura, S., S. Nakashima, G. Tanabe, Y. Oda, N. Yokota, K.
Fujimoto, T. Matsumoto, R. Sakuma, T. Ohta, K. Ogawa, S. Nishi- da, H. Miki, H. Matsuda, O. Muraoka, and M. Yoshikawa (2013) Alkaloid constituents from flower buds and leaves of sacred lotus (Nelumbo nucifera, Nymphaeaceae) with melanogenesis inhibi- tory activity in B16 melanoma cells. Bioorg. Med. Chem. 21: 779- 787.
57. Xu, W., L. Gong, M. M. Haddad, O. Bischof, J. Campisi, E. T. Yeh, and E. E. Medrano (2000) Regulation of microphthalmia-associ- ated transcription factor MITF protein levels by association with the ubiquitin-conjugating enzyme hUBC9. Exp. Cell Res. 255: 135- 143.
58. Choi, H., S. Ahn, H. Chang, N. S. Cho, K. Joo, B. G. Lee, I. Chang, and J. S. Hwang (2007) Influence of N-glycan processing disrup- tion on tyrosinase and melanin synthesis in HM3KO melanoma cells. Exp. Dermatol. 16: 110-117.
59. Mishima, Y. and G. Imokawa (1983) Selective aberration and pig- ment loss in melanosomes of malignant melanoma cells in vitro by glydosylation inhibitors: Premelanosomes as glycoprotein. J. Invest.
Dermatol. 81: 106-114.
60. Halaban, R. E., E. Cheng, Y. Zhang, G. Moellmann, D. Hanlon, M.
Michalak, V. Setaluri, and D. N. Hebert (1997) Aberrant retention of tyrosinase in the endoplasmic reticulum mediates accelerated degradation of the enzyme and contributes to the dedifferentiated phenotype of amelanotic melanoma cells. Proc. Natl. Acad. Sci.
USA 94: 6210-6215.