107 서 론
‘Guardian of the genome’으로 불리는1) 항종양 인자 p53
의 기능은 수많은 연구 보고에서 증명되었는데 homozy- gous와 heterozygous p53 knockout mice가 초기 종양 형성 에 민감성을 보인다고 보고된 바 있다.2) 이외에도 희귀 한 유전적 암으로 알려진 Li-Fraumeni syndrome 역시 일반
항종양 인자 p53에 의해서 유도되는 Base Excision Repair (BER)에 관한 연구: DNA Polymerase
Beta의 전사적 활성화를 중심으로
경희대학교 의과대학 약리학교실, 기초의과학연구소(IRBMS) 정 화 진․서 영 록
A Study On p53 Tumor Suppressor-mediated Base Excision Repair (BER) through the Activation of DNA Polymerase Beta in Transcription Level
Hwa Jin Jung and Young Rok Seo
Department of Pharmacology, Institute for Research of Medical Science (IRMS), College of Medicine, Kyung Hee University, Seoul 130-701, Korea
BER activity has been known to increase when DNA damage is induced by ionizing radiation (IR), simple alkylating agents, as well as free radicals exposed by endogenous hydrolytic and oxidative processes. The DNA polymerase beta enzyme as an important component of BER pathway, fills in the incised repair patch and participates in removing the overhang created by the displaced strand when treated the alkylating agent such as methyl methanesulfonate (MMS). Recent work has demonstrated that the p53 affects the stability of DNA polymerase beta and apurinic/apyrimidinic (AP) endonuclease interaction although it is not known whether the expression of DNA polymerase beta is regulated by p53 or not. Our data showed that a critical participator of BER activity, DNA polymerase beta can be overexpressed in the wild-type p53 background. We also found that the consensus p53 binding site in the promoter of DNA polymerase beta gene, indicating that the DNA polymerase beta might be transcriptionally regulated by p53. The regulation mechanism of DNA polymerase beta at the transcription level as well as protein interaction between p53 and DNA polymerase beta will provide the potential evidence how repair proteins are regulated to restore DNA damage induced by the environmental carcinogenesis. (Cancer Prev Res 11, 107-113, 2006)
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ Key Words: p53, DNA polymerase beta, Base excision repair
Correspondence to:Young Rok Seo
Department of Pharmacology, Institutue for Research of Medical Science (IRMS), College of Medicine, Kyung Hee University, 1 Heogi-dong, Dongdaemun-gu, Seoul 130-701, Korea
Tel: +82-2-961-0674, Fax: +82-2-963-0674 E-mail: [email protected]
책임저자:서영록, ꂕ 130-701, 서울특별시 동대문구 회기동 1 경희대학교 의과대학 약리학교실, 기초의과학연구소 Tel: 02-961-0674, Fax: 02-963-0674
E-mail: [email protected]
접수일:2006년 6월 1일, 게재승인일:2006년 6월 19일
인의 1%가 암에 걸릴 가능성을 가지는 반면, germ-line p53 mutation을 가진 Li-Fraumeni syndrome 가족 구성원은 그중 50%가 30세까지 암이 발병할 가능성이 있다고 알 려져 종양 억제자로서의 p53의 기능을 잘 설명해 준다.3) 또한 animal model을 이용한 연구에서 inducible p53 vector 를 주입한 경우, apoptosis에 의한 tumor xenografts의 regression을 유도한다고 보고되었다.4) 이러한 보고들은 정상적인 p53의 발현과 기능이 세포에 있어서 매우 중요 한 의미를 갖는다는 것을 보여준다.
지난 10년 동안 p53 단백질은 다양한 환경 스트레스에 의해서 acetylation과 phosphorylation 같은 post-translational modification 작용 기전을 통해5) 활성화되어 핵 내에 축적 된다고 알려져 왔다. 특히 가장 잘 알려져 있는 p53의 핵 내 축적을 유도하는 자극은 DNA 손상이다. 예를 들 어, 세포가 UV나 IR에 노출되면, dose-dependent하게 p53 의 핵 내 농도가 증가하게 된다. 게다가, 광범위하게 사 용되고 있는 chemotherapeutic agent들은 DNA를 손상시키 고 p53 단백질 축적을 유도한다고 알려져 있다.6) 그러나 최근에는 DNA에 직접적인 손상을 주지 않는 다른 형태 의 스트레스인 hypoxia, heat shock, nucleotide depletion 역 시 p53 축적을 유도할 수 있다고 보고되고 있다.7,8) 또한 oxidation/reduction (redox) modulation에 의해서 nuclear sig- naling protein redox factor-1 (Ref-1)을 통한 p53 activation의 조절이 제시된 바 있다.9,10)
활성화된 p53은 세포 내에서 cell cycle arrest, apoptosis, DNA repair와 같은 다양한 세포 반응을 유도하여 세포를 보호한다. 이러한 세포 반응은 p53 단백질의 transcription factor로서의 기능에 의해 특정 DNA sequence에 결합하여 target 유전자의 발현을 조절하여 유도될 수 있는데 일반 적으로 알려진 메커니즘은 target gene의 p53 consensus sequence에 p53이 binding하면서 전사적 활성을 유도해내 는 것이다. 예를 들면, p53-mediated cell cycle arrest의 작용 기전으로는 cyclin-dependent kinase inhibitor 역할을 하는 세포 내 유전자인 p21의 transactivation을 통해 cell cycle을 조절하는 메커니즘이 잘 알려져 있다.11) p53-mediated apoptosis의 경우, Fas와 DR5와 같은 death receptor의 tran- scriptional activation을 통해 유도되기도 하지만12) p53 단 백질은 Fas와 DR5뿐만 아니라, Bax, Noxa, p53 AIP1과 같 은 apoptosis에 직접적으로 관여하는 target gene들을 trans- activation시킴으로써 apoptosis를 유도하기도 한다.
DNA repair는 유전자 손상을 억제하여 세포 항상성을 유지시키는 기전으로 돌연변이를 1차적으로 제거하기 위해 매우 중요한 기작이다. 그중에서 특히 BER은 DNA 의 Base 손상을 수복하는 세포기전으로, 유해 산소나 변
이원성 alkylating agent에 의해 유발하는 유전자 손상을 복구하여 암과 노화를 억제하는 매우 중요한 생명체의 방어기전으로 알려져 있다. 이 과정은 손상된 base의 gly- cosylase에 의한 인식 또는 abasic site에 의해서 시작된다.
예를 들어 MMS 처리에 의해서 유도되는 일차적 손상은 N7-methylguanine과 3-methyladenine이다. 이것은 N-meth- ylpurine DNA glycosylase (MPG)에 의해서 인식되어 apuri- nic site를 생성하고 APE endonuclease에 의해서 AP site의 5’가 절단된다. DNA polymerase-β enzyme이 절단된 부분 의 nucleotide를 재합성하고 대체된 가닥에 의해 생성된 중복된 부분은 제거한다.13) 이차적인 절단 과정에서 관 여하는 효소는 flap endonuclease-1이다(Fen-1). 마지막으로 재합성된 가닥과 주형 가닥이 ligase I 또는 DNA ligase III/XRC1에 의해서 ligation되면서 repair 과정이 완료된다 (Fig. 1). 특히, beta-pol 유전자가 결여된 마우스는 BER 결 여 phenotype을 보이고 beta-pol-null 마우스에서 유래된 MEFs는 MMS에 민감성을 보인다.13) 이러한 연구 결과는
Fig. 1. Historical working model for alkaline-induced base exci- sion repair (BER). The DNA base damage generated by simple alkylating agents, as well as endogenous hydrolytic and oxida- tive processes was corrected by BER pathway. BER repair was initiated by a monofunctional glycosylase, then strand cleavage 5'to the apurinic/apyrimidinic (AP) site by AP endonuclease (APE). DNA synthesis and removal of the deoxyribose phos- phate (dRP) by DNA polymerase (-pol) was performed and ligation of the nick by a DNA ligase was induced. DNA pol- ymerase (-pol) was one of required genes for BER affected by p53 although the interaction mechanism between p53 and -pol was not clarified yet.
BER에 beta-pol이 매우 중요한 역할을 하고 있음을 제시 한다고 볼 수 있다. 그러나 최근 연구들이 DNA poly- merase beta에 초점을 맞춰 BER을 연구하고 있지만 아직 BER 과정에서 결정적인 인자 또는 단계가 무엇인지는 아직 알려져 있지 않다. 또한 p53과의 연관성은 거의 연 구가 되어있지 않았고, 특히 p53과 DNA polymerase beta 의 단백질 수준의 증가에 대한 작용 기전은 아직도 밝혀 지지 않고 있다. 본 연구에서는 p53에 의한 BER의 유도 에 관한 기전을 알아보기 위해 p53 protein과 BER 과정에 서 중요한 효소로 알려져 있는 DNA polymerase beta와의 연관성을 전사적 활성화의 측면에서 접근하였다.
재료 및 방법
1. 암세포 배양 및 base damaging agent인 MMS의 처리
본 연구에 사용된 세포주는 인간 대장 유래 세포인 wild type p53 RKO cell line과 mutant p53의 RKO-mp53 cell linedm로 90%의 RPMI-1640 (Gibco BRL, Grand Island, NY, USA)에 10% fetal bovine serum (FBS), 1%의 penicillin 및 streptomycin (Gibco BRL, Grand Island, NY, USA)이 포함된 배지를 사용하여 배양하였다. MMS는 Sigma Chemical Co.
(St. Luis, MO, USA)에서 구입하였으며, PBS (Gibco BRL, Grand Island, NY, USA)에 용해하여 -20oC에 보관하였 고, 매회 처리 전 배지에 희석 후 10μM 농도로 30분간 처리하였다.
2. Western blot analysis에 의한 단백질 발현의 분석 10μM의 MMS를 처리한 세포를 PBS로 washing하고 세 포를 수집한 다음 lysis buffer를 첨가하고 4oC에서 20분간 반응시킨 후 14,000 rpm에서 5분 동안 원심분리하여 상 층액의 단백질을 분리하였다. Bio Rad사의 Bradford rea- gent를 이용하여 단백질 정량을 수행하고 동량의 전체 세포 내 단백질(20 g)을 4∼12% gradient SDS-polyacryla- mide gel (Novex)로 분리하고 electroblotting을 통해 nitro- cellulose membrane으로 단백질을 transfer시킨다. 10% skim milk를 함유한 TBS-T (0.1% Tween 20)에 4oC에서 1시간 이상 blocking을 실시하였다. 본 실험에서 사용된 β- pol 항체는 rabbit anti-β-pol polyclonal antibody를 일차, horse- radish peroxidase (Santa Cruz)에 결합되어 있는 rabbit immunoglobulin G의 antibody를 사용하여 검출하였다. 고 정된 horseradish peroxidase 활성은 chemiluminescence (Pi- erce)로 검출한다.
3. Gel mobility assay in vitro
탐색된 DNA polymerase beta의 consensus sequence와 정 제된 p53 protein의 결합을 in vitro 및 세포에서 연구하기 위해 DNA polymerase beta의 공통 염기 서열(consensus se- quence) 후보 중에서 p53 protein에 결합 가능성이 높은 sequence를 결정하고, 동위원소로 표지된 probe를 만든다.
그런 다음, 동위 원소가 표지된 probe와 정제된 p53 protein을 Gel mobility buffer (BSA, NP-40, salmon sperm DNA, DTT 포함)와 p53 antibody (Pab 421)와 함께 반응 시킨 후 전기 영동을 하고, gel을 건조시킨 후, 암실에서 film에 조사한다.
결 과
1. Western blot을 통한 p53 단백질에 따른 DNA polymerase beta의 발현 조사
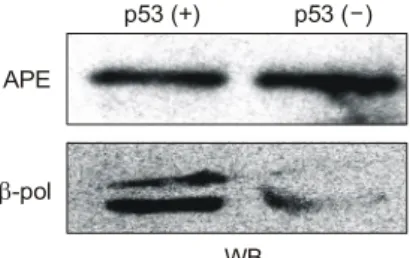
p53 wild-type과 mutant type의 세포에서 BER pathway에 중요한 역할을 한다고 알려져 있는 효소인 APE (apurinic endonuclease)와 DNA polymerase beta의 단백질 발현을 Western blot을 통해 조사한 결과, APE는 발현에 변화가 없었지만, DNA polymerase beta의 경우, p53 wild-type cell 에서는 정상적으로 발현하는 반면, mutant p53 cell에서는 거의 발현하지 않는 것이 관찰되었다. 이 결과를 통해 DNA polymerase beta의 발현이 p53 의존적으로 조절된다 는 것을 알 수 있다(Fig. 2).
2. DNA polymerase beta 내의 p53 protein- binding sequence 구축
p53 protein에 의해 조절 받는 다양한 downstream gene의 p53 binding sequence를 통해 consensus sequence를 조사하
Fig. 2. Expression of DNA polymerase beta in p53 wild type and -deficient cells, determined by immunoblotting. No ap- preciable differences were observed in APE, while DNA poly- merase beta level was found to decrease in p53-deficient cells.
Shown are RKO cells, p53 wild type or p53-mutant untreated cell.
p53 (+) p53 ( )- APE
β-pol
WB
고(PPPCWWGYYY N (0∼13) PPPCWWGYYY), 이를 통 해 DNA polymerase beta 내의 p53 protein과 결합가능성이 높은 sequence (GcGCAAGTaT N (5) AAtCAAGaTC)를 구축 하였다(Table 1).
3. DNA polymerase beta의 consensus sequence 와 p53 protein과의 결합 조사
DNA polymerase beta의 consensus sequence에 probe를 표 지시켜 p53 protein과의 결합을 Gel mobility assay 통해 조 사한 결과, p53 protein을 첨가하였을 때 band가 나타나는 것을 확인하였다. 이 결과를 통해 DNA polymerase beta의 consensus sequence와 p53 protein이 결합한다는 것을 알 있 고 나아가 β-pol activity가 p53에 의한 전사적 활성화를 통해 유도될 수 있는 가능성을 제시할 수 있다(Fig. 3).
고 찰
종양은 돌연변이를 야기하는 DNA sequence의 변화에 의해서 유도된다. DNA 변화는 식생활, life style과 같은 exogenous factor, 그리고 solar radiation과 같은 환경성 스트 레스뿐만 아니라, 일반적인 대사 과정에 의해서 유도될 수 있다. 이러한 유전적 변화의 축적을 막기 위해서 나타 나는 세포 반응이 DNA repair이다. 따라서 DNA repair의 결여는 carcinogenesis의 중요한 요인이 될 수 있다. DNA repair 중 Nucleotide Excision Repair (NER)는 UV-induced pyrimidine dimmer와 polycyclic aromatic hydrocarbone과 같 은 bulky DNA adducts를 제거하는 일차적 과정이다. 이와 대조적으로 Base Excision Repair (BER)는 reactive chemical에 의한 공격으로부터 유도되는 DNA 변형을 복구시키는
경로로 알려져 있다. Superoxide, hydroxyl radical, hydrogen peroxide와 같은 활성 산소종에 의해서 빈번히 생성되는 base damage는 BER에 의해서 복구된다. Endogenous alkyla- ting agent (예, S-adenosylmethionine)나 monofunctional alky- lating agent (예, methyl methanesulfonate (MMS), vinyl chlo- ride)의 노출에 의해 야기된 base alkylation damage 또한 BER에 의해서 복구된다. NER과 BER 간의 유사성은 두 경로 모두 20가지의 단백질로 구성된 multiprotein com- plex가 관여한다는 점이다.14) 두 번째 유사성은 손상된 Table 1. Comparison of known p53-binding regulatory sequences of known genes
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
Consensus PPCWWGYYY N (0∼13) PPPCWWGYYY (el-Deiry et al., 1994)
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
p21/Cip1/Waf1 GAACATGTCC N (0) cAACATGTTg (el-Deiry et al., 1993)
Gadd45 GAACATGTCT N (0) AAGCATGCTg (Kastan et al., 1992)
Mdm2 GGtCAAGTTg N (0) GGACAcGTCC (Barak et al., 1993)
Bax tcACAAGTTa N (1) AGACAAGCCT (Miyashita et al., 1995)
Rb AGCTAGagg N (3) GGGCGTGCCC (Osifchin et al., 1994)
Cyclin G AGACcTGCCC N (0) GGGCAAGCCT (Okamotoa and Beach, 1994)
EGF-R GAGCTAGCCC N (29) AGGCcAcCTC (Sheikh et al., 1997)
C-met GGACAAaCCT N (18) AGACAcGTgC (Seol et al., 1999)
KILLER/DR5 GGGCATGTCC N (0) GGGCAAGaCg (Takimoto et al., 2000)
β-polymerase GcGCAAGTaT N (5) AAtCAAGaTC (this paper)
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ The consensus sequence is indicated; P = (A, G); Y= (C, T); W = (A, T); N = (A, C, G, T).
Fig. 3. Interaction between p53 protein-binding sequences in DNA polymerase beta and p53 protein. Electrophoretic mobility shift assays were performed using p53 and oligonucleotides containing p53 protein-binding sequences as indicated. Lanes 1: free probes. Lanes 2: p53 protein-probes. An arrow indicates the band of shifted p53 protein bound in oligonucleotide of p53 protein binding sequences.
Probe Probe+p53
EMSA
DNA 부분의 절단과 함께 상보적인 주형 가닥을 이용해 수정된 정보의 연속적인 재합성이 이루어진다는 점이 다. 그러나 NER과 BER의 다른 점은 repair patch size로 BER의 경우 repair patch가 1∼6 base이지만 NER의 경우 는 repair patch가 30 base 정도이다.15) BER과 NER이 이루 어질 때 관여되는 multiprotein complex 또한 proliferating cell nuclear antigen (PCNA)과 XP-G (the product of the xero- derma pigmentosum complementation group G gene)과 같은 몇몇 complex는 BER, NER 모두에서 발견되기도 하지만 대부분 다르다.14) 모든 repairosome complex는 수복 과정에 서 재합성이 이루어질 때 관여하는 효소인 DNA poly- merase를 포함하는데 DNA polymerase ε와 δ는 NER에 DNA polymerase β는 BER에 관여한다.16)
지금까지 p53 활성에 의해 유도되는 cell cycle arrest와 apoptosis의 기전은 많이 연구되어 왔지만, p53 의존적 DNA repair 기전에 관해서는 거의 연구가 되어있지 않았 다. 그러나 최근 연구들은 p53이 DNA repair의 기능 증진 에 기여함으로써 유전적 돌연변이를 억제하는 데도 관 여한다고 보고하고 있어 새로운 암 억제 연구의 패러다 임으로서 p53-mediated DNA repair 연구가 높은 잠재력을 가지고 있음을 알 수 있다. p53은 UV-induced DNA dam- age, bulky carcinogen adducts 및 cisplatin과 같은 항암 치료
제에 의해 야기된 DNA 손상에 대한 수복 경로인 nucleo- tide excision repair (NER)에 중요한 역할을 하는 것으로 알 려져 있다. 종양 세포에서 흔히 나타나는 p53 단백질 기 능의 결여는 이런 형태의 손상에 대한 DNA 수복 현상을 감소시키고, 이로 인해 DNA 손상을 야기하는 물질들에 대한 민감성을 증가시키기도 한다. 최근에는 base-damag- ing agent인 methyl methanesulfonate (MMS)에 의한 DNA 손 상을 수복하는 base excision repair (BER) pathway에 p53이 관여한다는 사실이 연구되고 있다.
실제로, 최근 보고들과 본 연구자의 연구 결과에 따르 면 DNA polymerase beta 유전자가 결여된 세포는 BER ac- tivity의 결여와 base-damaging agent에 민감성을 보여 BER 에 DNA polymerase beta의 역할의 중요성이 제시되고 있
다.13,17,18) 뿐만 아니라 p53-null cell에서 MMS와 Thio-TEPA
와 같은 base-damaging agent에 대한 높은 민감성을 보이 는 연구 결과를 토대로, p53과 DNA polymerase beta간의 상호 작용에 대해 주목하게 되었다. 본 연구에서는 먼저 p53 단백질이 DNA polymerase beta protein의 발현에 영향 을 주는지 알아보기 위해 p53 wild-type과 deficient cell에 서 DNA polymerase beta 단백질의 발현을 Western blot을 수행하여 알아본 결과, DNA polymerase beta의 발현 수준 은 p53-deficient cell에서 더 낮다는 것을 확인하였다(Fig. 2).
Fig. 4. The roles of p53 tumor suppressor in response to genotoxic stesses. Activated p53 plays a role as a transcription factor for stimulating a range of cellular responses. p53 induces apoptosis to kill the damaged cells if the damage in cells is too extensive.
On the other hand, p53 enhances DNA repair and cell cycle arrest to protective cells against DNA damages. Recent reports suggested that p53 might play a direct or indirect role in DNA repair including nucleotide excision repair (NER) and base excision repair (BER) activating downstream gene Gadd45a or affecting the stabilization of repair protein, DNA polymerase beta, although the DNA repair mechanism of p53 was not clarified yet. p53-mediated cellular responses might provide a cancer preventive effect against various environmental stresses (Seo and Jung, 2004).
이 결과는 DNA polymerase beta의 단백질의 발현 또는 활 성이 p53-dependent하게 영향을 받는다는 것을 의미하는 것으로 DNA polymerase beta의 활성화 기전으로 pro- tein-protein interaction에 의한 발현 조절을 제시할 수 있 다. 뿐만 아니라 DNA polymerase beta와 p53의 복합체 형 성이 DNA polymerase beta의 안정성에 영향을 줄 수 있고 이러한 안정한 상호 작용은 p53-mutant와 null cell에서는 결여되어 있다는 사실에서 알 수 있다. 게다가, DNA polymerase beta activity는 p53-deficient cell의 extract에서는 반대로 나타난다.19) 그러나 p53의 소실과 관련된 DNA polymerase beta의 defection은 좀 더 신중히 생각되어야 한 다. 왜냐하면 p53-deficient cell은 적어도 MMS 처리 후에 검출하였을 때 여전히 DNA polymerase beta protein이 발 현되기 때문이다.
따라서 이러한 p53과 DNA polymerase beta의 활성화 기 전을 p53의 target gene으로서 DNA polymerase beta의 전 사적 활성화에 초점을 두고 연구를 진행해 보았다. 먼저, 탐색된 DNA polymerase beta의 consensus sequence와 정제 된 p53 protein의 결합을 in vitro 및 세포에서 연구하기 위 해 DNA polymerase beta의 공통 염기 서열(consensus se- quence) 후보 중에서 p53 protein에 결합 가능성이 높은 sequence를 결정하고(Table 1), in vitro에서 공통염기서열과 p53 protein 간의 결합을 조사하는 gel mobility assay를 수 행한 결과, p53 protein을 첨가했을 때 DNA polymerase beta의 공통 염기 서열과 결합한 band가 나타나는 것을 확인할 수 있었다(Fig. 3). 이 결과를 통해 p53 protein이 일반적으로 downstream gene의 consensus sequence에 결합 하여 전사를 조절하는 것과 같이 DNA polymerase beta도 p53의 target gene으로서 RNA 수준에서의 조절 받을 가능 성을 제시할 수 있다.
본 연구는 지금까지 p53에 의해서 유도되는 DNA repair pathway 중 현재 다양한 환경변이원 또는 생체 내 에서 대사 과정 중 발생하는 활성 산소와 같은 위험군에 의해 발생하는 DNA damage를 수복하는 BER에 관여하 는 것으로 알려진 DNA polymerase beta의 활성 메커니즘 을 밝히기 위한 것으로 p53의 전사적 조절 가능성에 대 한 심층적인 연구가 필요함을 제시할 수 있었다(Fig. 4).
이는 앞으로의 p53과 DNA polymerase beta의 단백질 수준 의 활성 조절이 어떻게 이루어지는지에 대한 메커니즘을 연구하는 데 중요한 지침이 될 수 있을 것이라 생각한다.
감사의 글
이 논문은 정부(교육인적자원부)의 재원으로 한국학
술진흥재단의 지원을 받아 수행된 연구의 일부임(KRF- 2003-003-E00014).
참 고 문 헌
1) Lane DP. Cancer. P53, guardian of the genome. Nature 358, 15-16, 1992.
2) Donehower LA, Harvey M, Slagle BL, McArthur MJ, Mont- gomery CA Jr, Butel JS, Bradley A. Mice deficient for p53 are developmentally normal but susceptible to spontaneous tumours. Nature 356, 215-221, 1992.
3) Markin D. p53 and the Li-Fraumeni syndrom. Biochem Biophys Acta 1198, 197-213, 1994.
4) Shaw P, Bovey R, Tardy S, Sahli R, Sordat B, Costa J. Induc- tion of apoptosis by wild-type p53 in a human colon tumor- derived cell line. Proc Natl Acad Sci USA 89, 4495-4499, 1992.
5) Ryan km, Phillips AC Vousden KH. Regulation and function of the p53 tumor suppressor protein. Curr Opin Cell Biol 13, 332-337, 2001.
6) Fritsche M, Haessler C, Brandner G. Induction of nuclear accumulation of the tumor-suppressor protein p53 by DNA- damaging agents. Oncogene 8, 307-318, 1993.
7) Graeber TG, Peterson JF, Tsai M, Monica K, Fornace AJ Jr, Giaccia AJ. Hypoxia induces accumulation of p53 protein, but activation of a G1-phase checkpoint by low-oxygen condition is independent of p53 status. Mol Cell Biol 14, 6264-6277, 1994.
8) Yin Y, Terauchi Y, Solomon GG, Aizawa S, Rangarajan PN, Yazaki Y, Kadowaki T, Barrett JC. Involvement of p85 in p53-dependent apoptotic response to oxidative stress. Nature 391, 707-710, 1998.
9) Ueno M, Masutani H, Arai RJ, Yamauchi A, Hirota K, Sakai T, Inamoto T, Yamaoka Y, Yodoi J, Nikaido T. Thiore- doxin-dependent redox regulation of p53-mediated p21 activation. J Biol Chem 274, 35809-35815, 1999.
10) Gaiddon C, Moorthy NC, Prives C. Ref-1 regulates the transactivation and pro-apoptotic functions of p53 in vivo.
EMBO J 185, 5609-5621, 1999.
11) El-Deiry WS, Tokino T, Velculescu VE, Levy DB, Parsons R, Trent JM, Lin D, Mercer WE, Kinzler KW, Vogelstein B.
WAF1, a potential mediator of p53 tumor suppression. Cell 75, 817-825, 1993.
12) Sheikh MS, Fornace AJ Jr. Death and decoy receptors and p53-mediated apoptosis. Leukemia 14, 1509-1513, 2000.
13) Wilson SH, Kunkel TA. Passing the baton in base excision repair. Nat Struct Biol 7, 176 -178, 2000.
14) Matsumoto Y, Kim K, Hurwitz J, Gary R, Levin DS, Tomkinson AE, Park MS. Reconstitution of proliferating cell nuclear antigen-dependent repair of apurinic/apyrimidinic sites with purified human proteins. J Biol Chem 274, 33703-33708, 1999.
15) Friedberg EC, Bardwell AJ, Bardwell L, Feaver WJ, Kornberg RD, Svejstrup JQ, Tomkinson AE, Wang Z. Nucleotide exci- sion repair in the yeast Saccharomyces cerevisiae: its rela- tionship to specialized mitotic recombination and RNA polymerase II basal transcription. Philos Trans R Soc Lond B Biol Sci 347, 63-68, 1995.
16) Stucki M, Pascucci B, Parlanti E, Fortini P, Wilson SH, Hubscher U, Dogliotti E. Mammalian base excision repair by DNA polymerase delta and epsilon. Oncogene 7, 835-843, 1998.
17) Sobol RW, Prasad R, Evenski A, Baker A, Yang XP, Horton JK, Wilson SH. The lyase activity of the DNA repair protein beta-polymerase protects from DNA-damage-induced cytotox- icity. Nature 15, 807-10, 2000.
18) Seo YR, Sweeney C, Smith ML. Slenomethionine induction of DNA repair response in human fibroblasts. Oncogene 21, 3663-3669, 2002.
19) Zhou J, Ahn J, Wilson SH, Prives C. A role for p53 in base excision repair. EMBO J 20, 914-923, 2001.