Phylogenetic Analysis of Korean Black Cattle Based on the Mitochondrial Cytochrome b Gene
Jae-Hwan Kim
1*, Mi Jung Byun
1, Myung-Jick Kim
1, Sang Won Suh
1,2, Young-Sin Kim
1, Yeoung-Gyu Ko
1, Sung Woo Kim
1, Kyoung-Sub Jung
3, Dong-Hun Kim
1and Seong-Bok Choi
11
National Institute of Animal Science, R.D.A., Namwon 590-830, Korea
2
Division of Applied Life Science, Graduate School of Gyeongsang National University, Jinju 660-701, Korea
3
Chungbuk Institute of Livestock and Veterinary Research, Cheongwon, Chungbuk 363-931, Korea Received September 13, 2012 /Revised December 28, 2012 /Accepted January 3, 2013
The purpose of this study was to identify genetic polymorphisms of the mitochondrial cytochrome b (mtDNA cyt b) gene in Korean black (KB) cattle breed and to analyze the genetic relationship be- tween the KB and other breeds. We determined the complete sequence of the mtDNA cyt b gene in 38 KB cattle. We also analyzed their genetic diversity, and phylogenetic analysis was performed by comparison with Korean cattle (KC, called Hanwoo) and breeds from China and Japan. A nucleotide substitution was detected in the KB cattle, and two haplotypes were defined. In the neighbor-joining (NJ) tree, the haplotypes of KB were located in Bos taurus lineage with those of KC, Japanese black (JB), Yanbian and Zaosheng breeds. However, the haplotypes of Chinese breeds, excluding Yanbian and Zaosheng, were separated into B. taurus and B. indicus lineages. In the NJ tree of breeds based on Dxy genetic distances, Chinese breeds mixed with B. taurus and B. indicus lineages were located between B. indicus and B. taurus lineages. KB was contained within B. taurus lineage and was de- termined to be genetically more closely related to two Chinese (Yanbian and Zaosheng) breeds than to KC and JB. The haplotype distribution and the results of the phylogenetic analysis suggest that KB and KC have genetic differences in their mtDNA cyt b gene sequences.
Key words : Cattle, Korean black, mtDNA, cytochrome b, relationship
*Corresponding author
*Tel:+82-63-620-3522, Fax:+82-63-620-3590
*E-mail : [email protected]
This is an Open-Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Life Science 2013 Vol. 23. No. 1. 24~30 DOI : http://dx.doi.org/10.5352/JLS.2013.23.1.24
서 론
현재 우리가 사육하고 있는 소들은 약 10,000~15,000년 전 에 아시아, 유럽, 북부아프리카 등 지역적으로 넓게 분포했던 야생소(Bos primigenius)의 가축화로부터 유래되었을 것으로 추정하고 있다[19]. 하지만 최근 들어 고고학적 연구 및 mi- tochondrial DNA (mtDNA)를 이용한 유전학적 연구의 결과 들은 현재의 소들은 적어도 2개의 독립적이고 지리적인 특성 을 갖는 가축화가 서남아시아에서 발생했다는 증거들을 제시 하고 있다[5, 11]. 첫째는 Fertile Crescent 지역에서 발생한 T haplogroup (B. taurus 계열), 둘째는 Indus Valley에서 발생한 혹이 있는 I haplogroup (B. indicus 계열)이다. T haplogroup 은 mtDNA D-loop 분석에 의해 특징적인 지역적 분포를 나타 내는 5개 sub-haplogroup(T-T5)으로 나눠지며[2, 20, 32], I
haplogroup은 T haplogroup과 다른 유전적 양상을 보이며 I1, I2 등 2개의 sub-haplogroup으로 분류된다[3].
최근 들어 생물자원에 대한 국제적 패러다임의 변화로 자원 보유국의 배타적 권리가 인정되면서 자원의 확보, 보존, 관리 와 더불어 특성평가 및 활용 등에 많은 관심과 노력을 기울이 고 있다. 특히 2007년 인터라켄 선언 이후 UN 식량농업기구 (FAO)와 가입국들의 참여로 전세계에 분포하고 있는 가축유 전자원의 다양성 보존 및 특성평가에 대한 노력이 진행 중에 있다[7]. 자원보유국들은 말[1, 9], 소[14, 16], 돼지[10, 15], 양 [22, 31] 및 염소[18, 25] 등 가축유전자원에 대한 유전적 다양 성 평가 및 계통유전학적 분석 등 분자유전학적인 특성평가 결과를 보고하고 있다.
일반적으로 mtDNA는 모계유전특성을 가지고 있기 때문
에여러 가축을 대상으로 mtDNA를 이용한 가축의 기원, 가
축화 시기의 해석과 유전적 다양성 분석을 위한 많은 연구결
과가 보고되었다[12, 24, 33]. 특히 D-loop과 cytochrome b
(Cyt b) 유전자 등 2개 영역이 대표적인데, D-loop은 비-암호
화(non-coding) 영역으로 염기변이가 높게 나타나는 반면
Cyt b 유전자는 암호화(coding) 영역으로 D-loop에 비해 염
기변이율이 낮다. Cyt b 유전자는 전체 1.2 kb로서, 낮은 재
조합 빈도를 보이며, 유전자의 구조와 서열이 이미 잘 알려
져 있기 때문에 소, 돼지, 염소, 닭 등 다양한 종에서 계통유 전학적 분석을 위한 마커로 사용되고 있다[26-28, 34].
현재 FAO 산하의 가축다양성정보시스템(DAD-IS, http:
//dad.fao.org/)에는 전세계 국가로부터 약 3,000여 개의 소 품종이 등재되어 있으며, 이들 중 우리나라 소 품종은 4개 재 래품종과 4개 외래품종 등 8품종이 등재되어 있다. 재래품종 중 전신 흑모색의 특성을 보이는 한국흑우는 현재 멸종 위기 종으로 보존 및 증식을 위해 국가적으로 노력하고 있으나 이 들에 대한 분자유전학적 특성분석은 전무한 실정이다. 따라 서 본 연구는 mtDNA Cyt b 유전자의 서열에 기초한 한국흑 우의 유전적 다양성 파악 및 계통유전학적 특성 분석을 위해 수행하였다.
재료 및 방법
시료 수집 및 DNA 추출
본 연구를 위해 공시된 38두의 한국흑우는 우리나라 가축유 전자원의 보존 및 관리를 위한 관리기관 중 충청북도 축산위생 연구소 종축시험장으로부터 혈액으로 제공받았다. Genomic DNA는 Sucrose-Proteinase K 방법[4]을 이용하여 추출하였으 며, NanoDrop ND1000 (Thermo Scientific, USA)을 이용하여 genomic DNA의 농도측정 후 PCR 증폭을 위한 주형을 제작 하였다.
Cyt b 유전자 증폭 및 서열결정
소 mtDNA Cyt b 유전자에 대한 전체 coding 영역을 증폭 하기 위해서 Bcyt-F (5’- TTC TTA CAT GGA ATC TAA CCA TGA-3’)와 Bcyt-R (5’-GGG AGG TTA GTT GTT CTC CTT CTC-3’)을 이용하였다. 이 primer는 기존에 보고된 mtDNA genome 서열(GenBank accession no. V00654)를 이용하여 tRNA-Glu와 tRNA-Thr 서열을 이용하여 제작하였다. PCR 반 응은 Taq polymerase 1.5 unit (TaKaRa, Japan), 10×buffer 2.5 μ l, 0.2 mM dNTP, 1.5 mM MgCl
2, 10 pmol primer 각각 1.5 μ l, genomic DNA 20 ng 그리고 증류수를 첨가하여 최종 25 μ l로 반응하였다. PCR 증폭은 PTC-200 thermocycler (MJ Research Inc.)을 이용하여 94℃에서 2분간 초기변성 후 94℃
에서 30초, 60℃에서 30초, 72℃에서 60초를 1회로 하여 35회 반복수행 하였으며, 72℃에서 5분간 최종 신장하였다. PCR 증 폭산물은 1.5% agarose 전기영동 상에서 최종 확인한 후 QIAquick PCR purification Kit (Qiagen, USA)로 정제하였다.
염기서열 분석은 ABI 3130xl Genetic Analyzer (ABI, USA)를 이용하여 direct-sequencing을 수행하였다. 분석된 서열은 소 mtDNA genome 서열(GenBank accession no.V00654)을 이용 하여 Cyt b 유전자의 전체서열을 결정하였으며, GenBank da- tabase에 등록하였다(JX472260-1).
유전특성 및 계통유전학적 분석
본 연구에서 결정된 한국흑우 Cyt b 유전자의 서열을 대상 으로 CLUSTAL W 프로그램[30]을 이용하여 다중염기정렬을 실시하였다. 변이부위 탐색, 유전적 변이성 추정 및 haplotype 의 결정은 DNASP (ver. 5.1) [17]을 사용하였다. 결정된 한국 흑우 서열과 GenBank database로부터 수집된 12품종 101개 서열을 이용한 계통유전학적 분석을 위해 Neighbor-joining (NJ) tree [23]를 작성하였다. Haplotype 간에 작성된 NJ tree는 Kimura-2 parameter model을 적용한 Mega 5.05 package [29]
를 이용하였으며, 계통도의 신뢰도를 평가하기 위해 1,000회 반복실행 후 bootstrap value [8]를 산출하였다. 또한 품종간 NJ tree는 DNASP (ver. 5.1)로 산출된 Dxy 유전적 거리를 토대 로 작성하였다.
결과 및 고찰
한국흑우 38두로부터 결정된 mtDNA Cyt b 유전자의 전 체 염기서열 길이는 1,140 bp로 확인되었고, 개시코돈과 종 료코돈이 ATG와 AGA로 모든 개체에서 동일하게 나타났다.
또한 4개 염기 중 G 염기 구성비율(13.4%)이 A (31.2%), C (30.2%), T (25.2%) 염기에 비해 상대적으로 낮게 나타났으 며, 이런 결과는 기존에 보고된 다른 소 품종의 결과와 매우 유사하였다[6].
결정된 38개 Cyt b 유전자 서열의 다중염기정렬에서 염기 삽입 및 결실 없이 단지 1개의 염기변이(GenBank accession no. DQ186215 기준 997번째)가 동정되었으며, 이 염기변이는 아미노산 치환에 영향을 주지 않는 silent mutation으로 확인 되었다. 2개의 haplotype으로 분류되었으며, haplotype 다양 성 지수는 0.235, 염기변이율은 0.00021로 나타났다(Table 1).
한국흑우의 haplotype 다양성 지수 및 염기변이율은 기존에 보고된 중국 품종들보다 현저히 낮게 나타났다[6]. 본 연구에 서 분석에 이용한 한국흑우의 시료수집 영역의 한계가 이러한 유전적 다양성의 감소에 일부 작용했을 것으로 판단된다.
한국흑우 38두와 타 품종들과의 haplotype 분포 및 연관성 을 확인하기 위하여 GenBank database로부터 한국, 일본, 중 국에 분포하는 12품종 101개 Cyt b 유전자의 전체서열을 수집 하여 분석에 이용하였다(Table 2). 전체적으로 30개 haplotype 으로 분류되었는데, 한국흑우 38두 중 33두가 haplotype-1 (H1)에 속하였고, 나머지 5두는 H2에 포함되었다(Table 3).
H1에는 중국의 Luxi 품종을 제외한 모든 품종이, H2에는 4품
종(Kazakh, Menggu, Jinnan, Jiaxian)을 제외한 나머지 품종이
포함되었다. 한국흑우와 Yanbian을 제외한 모든 품종은 1~7
개의 품종-특이 haplotype을 갖는 것으로 나타났고, 특히 한우
(KC) 품종에서 4개, Kazakh 품종에서는 7개가 확인되었다. 한
국흑우와 Yanbian은 품종-특이 haplotype 없이 H1과 H2에
모든 개체가 포함되었다. H2에 비해 H1에 상대적으로 많은
Table 1. Genetic diversity of KB based on the complete mtDNA Cyt b gene sequence Region Population
1Sample size Number of
polymorphic sites Number of
haplotypes Haplotype
diversity Nucleotide diversity
Total 38 1 2 0.235 0.00021
Table 2. Information on reference sequences for comparison with KB
Country Breeds Abbreviation GenBank accession no.
Korea Korean black KB JX472260-1
Korean cattle (Hanwoo) KC AY526085, DQ124371-86, NC_006853
Japan Japanese black JB AB074962-8
China Yanbian Yan AY903438, DQ186215-9
Zaosheng Zao DQ186220-6
Kazakh Kaz DQ186203-13
Menggu Men AY885291-3, AY952959
Qinchuan Qin AY885304-6, AY903439, AY952952-3, DQ186241-2
Jinnan Jin DQ186229-36
Bohai Black Boh AY885297-300, AY952962-3, DQ186227-8
Nanyang Nan AY885283-7, AY952955, DQ186253-4
Luxi Lux AY885294-6, AY952960-1, DQ186237-40
Jiaxian Jia AY906832-4, DQ186243-6
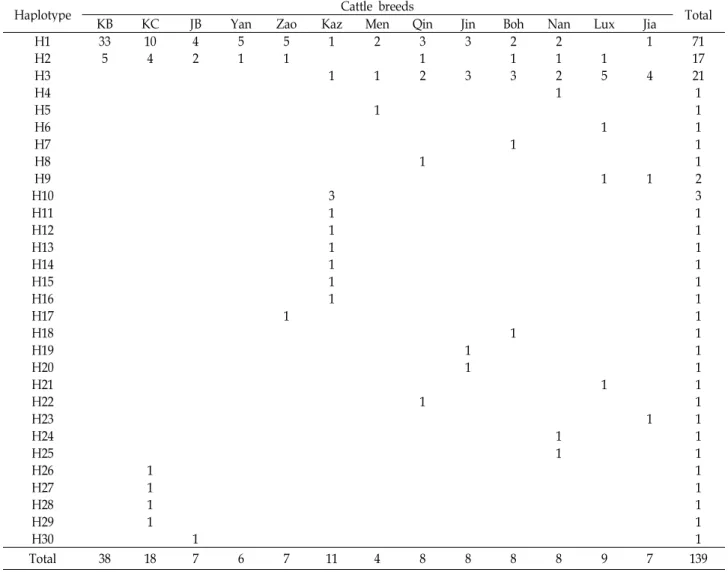
Table 3. Distribution of mtDNA haplotypes in 13 cattle breeds based on Cyt b gene sequences
Haplotype KB KC JB Yan Zao Kaz Cattle breeds Men Qin Jin Boh Nan Lux Jia Total
H1 33 10 4 5 5 1 2 3 3 2 2 1 71
H2 5 4 2 1 1 1 1 1 1 17
H3 1 1 2 3 3 2 5 4 21
H4 1 1
H5 1 1
H6 1 1
H7 1 1
H8 1 1
H9 1 1 2
H10 3 3
H11 1 1
H12 1 1
H13 1 1
H14 1 1
H15 1 1
H16 1 1
H17 1 1
H18 1 1
H19 1 1
H20 1 1
H21 1 1
H22 1 1
H23 1 1
H24 1 1
H25 1 1
H26 1 1
H27 1 1
H28 1 1
H29 1 1
H30 1 1
Total 38 18 7 6 7 11 4 8 8 8 8 9 7 139
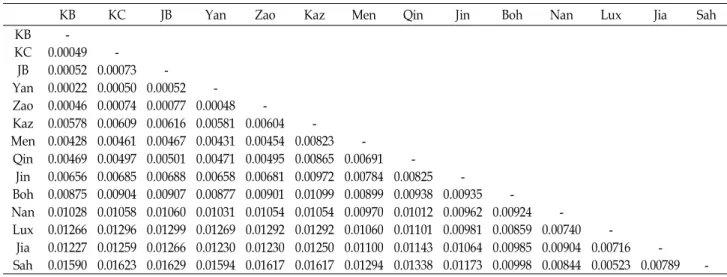
Table 4. D xy genetic distances among 14 cattle breeds based on mtDNA Cyt b gene sequences
KB KC JB Yan Zao Kaz Men Qin Jin Boh Nan Lux Jia Sah
KB -
KC 0.00049 - JB 0.00052 0.00073 - Yan 0.00022 0.00050 0.00052 - Zao 0.00046 0.00074 0.00077 0.00048 - Kaz 0.00578 0.00609 0.00616 0.00581 0.00604 - Men 0.00428 0.00461 0.00467 0.00431 0.00454 0.00823 -
Qin 0.00469 0.00497 0.00501 0.00471 0.00495 0.00865 0.00691 - Jin 0.00656 0.00685 0.00688 0.00658 0.00681 0.00972 0.00784 0.00825 - Boh 0.00875 0.00904 0.00907 0.00877 0.00901 0.01099 0.00899 0.00938 0.00935 - Nan 0.01028 0.01058 0.01060 0.01031 0.01054 0.01054 0.00970 0.01012 0.00962 0.00924 - Lux 0.01266 0.01296 0.01299 0.01269 0.01292 0.01292 0.01060 0.01101 0.00981 0.00859 0.00740 -
Jia 0.01227 0.01259 0.01266 0.01230 0.01230 0.01250 0.01100 0.01143 0.01064 0.00985 0.00904 0.00716 - Sah 0.01590 0.01623 0.01629 0.01594 0.01617 0.01617 0.01294 0.01338 0.01173 0.00998 0.00844 0.00523 0.00789 -
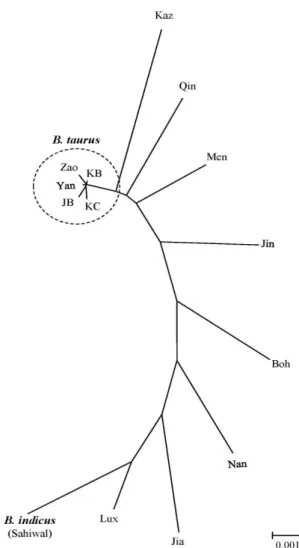
Fig. 1. Phylogenetic relationship among 30 hap- lotypes of the mtDNA Cyt b gene from cattle breeds of Korean, Japan and China.
The tree was constructed with neigh- bor-joining method. The numbers at branches stand for bootstrap values high- er than 50% out of 1000 replications. The bar scale indicates genetic distance.
Abbreviations for population or breed are the same as those in Table 2.
개체가 속하였으며, 특히 한국흑우, 한우, 일본흑우(JB), Yanbian, Zaosheng 품종의 60% 이상의 개체가 H1에 포함되 었다. 이런 결과는 전체 30개의 haplotype 중 H1과 H2가 한국, 일본, 중국의 품종들에서 상대적으로 높은 빈도로 나타나는 major haplotype임을 시사한다.
한국, 일본, 중국의 소품종에서 나타난 30개 haplotype 간의 유연관계를 확인하기 위하여 NJ tree를 작성하였다(Fig. 1). 이
때 Bos indicus에 속하는 파키스탄의 Sahiwal 품종 3개 서열
(GenBank accession no. JN117611-3)을 포함시켰다. 그 결과,
Group I과 Group II로 명확히 구분되었는데, Group I에는 H1
과 H2를 포함한 21개의 haplotype이 포함된 반면 Group II는
9개의 haplotype과 Sahiwal 품종 3개 개체 모두를 포함하였
다. 기존 보고에 의하면 중국재래소인 경우 지리적인 분포 및
유전적 특성에 따라 북부, 중부, 남부 등 3개의 그룹으로 나눌
수 있는데[6, 16], 북부그룹은 B. taurus의 영향을 많이 받은 반면 남부그룹은 B. taurus보다는 B. indicus에 의한 영향이 크 고, 중부그룹인 경우 B. taurus와 B. indicus 모두의 영향을 받았 을 것으로 추정하고 있다. 반면에 한국과 일본의 소 품종은 모두 B. taurus이며 아직까지 B. indicus에 속하는 개체 혹은 집단에 대한 보고는 없다[13, 14, 21]. 이와 같은 결과들을 토대 로 해석해보면 본 연구의 haplotye에 기초한 NJ tree 상에서 Sahiwal과 함께 Group II에 위치하고 있는 9개 haplotype은 B. indicus 계열이며, 기존에 B. taurus로 알려진 한우, 일본흑우 를 포함하고 있는 Group II의 21개 haplotype은 B. taurus 계열 로 판단된다. 따라서 본 연구의 분석에 포함된 품종들 중 B.
taurus와 B. indicus의 특징이 동시에 나타나는 품종은 Yanbian 과 Zaosheng를 제외한 모든 중국 품종이며, B. indicus의 특징 을 보이는 9개의 haplotype에 포함되는 30개 개체 중 21개를 포함하는 H3이 major haplotype으로 나타났다. 결국 한국흑 우는 38두 전체가 B. taurus lineage 내부의 major haplotype인 H1, H2에 속하는 것으로 확인되었다.
한국흑우를 포함한 품종 간의 유연관계를 확인하기 위하여 Dxy 유전적 거리를 산출하였고(Table 4) 이를 이용하여 NJ tree를 작성하였다(Fig. 2). 한국흑우, 한우, 일본흑우, Yanbian, Zaosheng 등 5개 품종들은 가까운 유연관계를 보이며 하나의 group을 형성하였으나, Yanbian과 Zaosheng 품종을 제외한 8개 중국 품종들은 B. indicus (Sahiwal 품종)와 B. taurus 사이 에 배열되어 나타났다. 이와 같은 결과는 앞서 haplotype 분석 (Table 3, Fig. 1)에서도 제시한 바와 같이 한국흑우, 한우, 일본 흑우, Yanbian, Zaosheng 등 5개 품종은 단지 B. taurus의 특징 만을 보유하는 반면 나머지 품종들은 B. taurus와 B. indicus의 유전적 특성을 동시에 가지고 있기 때문으로 사료된다. B. taur us에 속하는 5개 품종들 간에 산출된 Dxy 유전거리 확인 결과 (Table 4), 한국흑우는 한우, 일본흑우와 각각 0.00049, 0.00052 의 유전적 거리를 보인 반면 Yanbian, Zaosheng과는 0.00022, 0.00046으로 확인되었다. 유전거리는 비교된 개체 혹은 집단 간에 나타나는 염기서열의 유사성에 근거하며, 유전거리가 낮 을수록 유전적 유연관계가 높음을 의미한다[6, 14]. 따라서 본 연구에서의 한국흑우는 한우보다 Yanbian, Zaosheng 품종과 유전적 유연관계가 높은 것으로 확인되었다. 최근에 보고된 mtDNA Cyt b 유전자의 서열에 기초한 연구결과[6]에 따르면 분석에 이용된 중국소 18품종 중 Yanbian 및 Mongolia 품종은 B. taurus의 모계특징만을 보였으며, Zaosheng 품종도 일부 B. indicus의 특징이 나타나지만 B. taurus의 모계특징이 85.7%
로 높게 나타남을 보고하였다. Yanbian과 Mongolia 품종은 한반도와 인접한 중국의 북부에 분포하고 있으며, Zaosheng 품종도 한반도와는 상대적으로 멀긴 하지만 중국 북부에 분포 하는 품종이다. 본 연구와 기존에 보고된 연구들의 결과를 종 합해보면, 한국흑우와 한우 품종은 한반도와 인접한 중국북부 에 분포하는 Yanbian, Zaosheng 품종들과 모계적으로 가깝게
Fig. 2. Phylogenetic relationship among 12 cattle breeds of B.
taurus lineage and a breed of B. indicus lineage (GenBank accession no. JN117614-5). The tree was con- structed with neighbor-joining method on the basis of D xy distances. The bar scale indicates genetic distance.
Abbreviations for population or breed are the same as those in Table 2.
연관되어 있으며, 특히 한국흑우는 Yanbian 품종과 상대적으 로 높은 유연관계가 있음을 시사한다.
본 연구에서는 mtDNA Cyt b 유전자 서열을 토대로 한국흑
우의 유전적 다양성 및 타 품종들과의 계통유전학적 분석을
통해 한국흑우와 중국의 일부 품종과의 상대적으로 가까운
유전적 유연관계가 있음이 확인되었다. 비록 분석에 이용된
한국흑우가 한 지역에서만 수집되었고 비교 품종의 개체수가
적다는 한계가 있으나, 본 연구의 결과는 가축유전자원으로서
한국흑우의 보존 및 유전적 특성 구명을 위한 중요한 자료로
활용이 가능할 것으로 판단된다. 한국흑우의 기원 및 유입과
정에 대한 더욱 명확한 해석을 위해서는 여러 품종에 대한
충분한 시료확보가 필요하며, 또한 핵 DNA (초위성체마커,
Y 염색체 등) 분석이 추가되어야 할 것으로 사료된다.
감사의 글
본 연구는 농촌진흥청 연구개발사업 어젠다(2-5-12) 과제 (과제번호: PJ008431) 지원에 의한 연구결과의 일부임.
References