현미 발아 특성에 따른 수용성 비타민 및 GABA 함량 변화
정범균1․문현규2․천지연2
1순천천연물의약소재개발연구센터
2순천대학교 식품공학과
Water-Soluble Vitamin and GABA Contents of Brown Rice Affected by Germination
Beom-Gyun Jeong1, Hyun-Gyu Moon2, and Jiyeon Chun2
1Suncheon Research Center for Natural Medicines
2Department of Food Science and Technology, Sunchon National University
ABSTRACT The effects of germination and rice types on the contents of water-soluble vitamins and γ-aminobutyric acid (GABA) and saccharification power of brown rice were investigated. Three types of rice, saeilmi (SI), seolgaeng (SG), and shintoheukmi (SH), were immersed separately for 24 h at 35°C and incubated at 30°C (90% humidity) for 24 h for germination. The longest length of the germinated sprout for SI, SG, and SH reached 4, 2, and 7 mm, respectively. The niacin, folic acid, and GABA contents of brown rice were increased two-three fold in all three cultivars by germination. During germination, the thiamin content increased two fold for SI and SG while it decreased remarkably in SH. The highest niacin (34.84 µg/100 g), folic acid (48.45 µg/100 g), and GABA (33.86 mg/100 g) contents were observed in SH but SI had a higher thiamin content (593.27 µg/100 g). The glucose content was increased significantly in all three varieties by germination but the largest increase was found in SH (14.38%). Increases in saccharification power were observed in SI and SH. These results show that germination would be used for the prepara- tion of brown rice with enhanced saccharification power as well as beneficial components, such as vitamins and GABA.
Key words: brown rice, germination, GABA, water-soluble vitamin
Received 19 August 2019; Accepted 9 September 2019 Corresponding author: Jiyeon Chun, Department of Food Science and Technology, Sunchon National University, Suncheon, Jeonnam 57922, Korea
E-mail: [email protected], Phone: +82-61-750-3258
Author information: Beom-Gyun Jeong (Graduate student), Hyun- Gyu Moon (Graduate student), Jiyeon Chun (Professor)
서 론
쌀은 예로부터 한국인의 주식으로 이용되었으나 최근 한 국인의 서구적인 식습관으로의 변화에 따라 1인당 연간 쌀 소비량은 2000년 93.6 kg, 2009년 74.0 kg, 2018년 61.0 kg으로 지속적인 감소를 보이고 있다(Statistics Korea, 2019). 이러한 감소 추세는 앞으로도 계속 지속할 전망으로 현재 생산대비 소비의 감소로 잉여되는 쌀 소비 확대를 위해 다각적인 쌀 활용 연구가 필요하다.
현미는 전곡립 형태로 과피, 종피 및 호분층으로 미강, 배 및 배유로 구성되어 있다. 현미에는 단백질, 비타민 B1, B2, 무기질, 식이섬유가 풍부히 존재하지만, 백미보다 수분 흡수속도가 낮고 질감이 거친 특성은 식미로의 활용에 단점 이 되고 있다(Kim 등, 2001; Cho 등, 2011). 이러한 현미의 거친 식감을 보완하면서 건강에 유용한 성분을 증진할 수
있는 방법으로 발아 가공방법이 널리 이용되고 있다.
발아(germination)란 씨앗을 적정한 수분, 온도, 산소 등 의 조건에서 싹이 나오는 것을 말한다. 발아 과정은 씨앗의 조직을 연화시켜 질감을 개선할 뿐만 아니라 각종 기능성 미 량성분들을 향상시키는 것으로 알려져 있다(Kim 등, 2001;
Kim 등, 2011). 종자 발아에 관한 연구는 비타민 E, squa- lene, phytosterols, GABA 등의 기능성 성분 변화(Choi 등, 2004; Jung 등, 2008; Moongngarm과 Saetung, 2010), 발아 조건, 건조 방법(Kim 등, 2001; Kim 등, 1998), 가공원 료로서의 이용(Oh, 2007; Cho 등, 2011), 녹두(Ko와 Park, 1983), 유채(Kim 등, 1988a; Kim 등, 1988b), 메밀(Lee 등, 1995a), 벼(Kim 등, 2012), 보리(Cha 등, 2012) 등 다양한 자원들을 활용한 연구들이 보고되어 있다. 발아현미의 경우 식혜(Kim과 Lee, 1997), 쌀 가공식품 제조(Kang 등, 2003) 등으로의 활용 가능성을 검토하는 연구들이 진행되어 왔다.
최근 한국산 쌀을 활용하여 맥주 원료로 사용하기 위한 연구 (Lee 등, 1995b; Hyeun 등, 2012)가 보고되어 있으나 아직 발아현미를 맥주 원료로 적용하기 위한 특성 연구는 거의 보고된 바가 없다.
2015년 가공식품 세분시장 현황 보고서(Korea Agro-
Fisheries & Food Trade Corporation, 2016)에서 국세통 계 기준 2009년 맥주의 출고량은 1,961,568 kL에서 2014 년 2,055,761 kL로 증가하고 있으며, 수입 맥주도 2009년 41,492톤에서 2014년에 119,501톤으로 증가하고 있다. 1 인당 주류 소비량 중 맥주의 경우 146.3병(2009년)에서 148.3병(2014년)으로 2.0병이 증가했지만 소주는 67.9병 에서 65.3병으로 2.6병 감소하였는데, 이는 국내 음주문화 가 변화되어 과음을 자제하고 저도주 문화로 변했기 때문이 라고 볼 수 있다. 한국수제맥주협회에 따르면 2010년, 2014 년, 2018년 3차례의 주세법 완화로 인하여 소규모 맥주 제 조 사업자는 2014년도 54개에서 2018년 109개로 증가하 였다. 하지만 아직 맥주 제조에 큰 비중을 차지하는 맥아는 대부분 전량 수입산 맥아를 사용하고 있기 때문에 국내산 발아벼를 맥주 제조에 적용 시 맥아 수입을 줄일 수 있으며 국내산 쌀의 소비를 높일 수 있는 경제적 이익을 창출할 수 있을 것으로 기대된다.
이에 본 연구에서는 새일미, 설갱 및 신토흑미 세 가지 현미 품종에 대한 특성 조사와 이들을 발아시킨 발아현미와 영양성분, 기능성 성분 및 당화 특성을 비교하여 발아현미의 식품 산업적인 활용을 위한 기초자료로 활용하고자 한다.
재료 및 방법
실험재료
본 실험에 사용한 벼는 총 3품종으로 새일미(saeilmi, SI) 는 친환경농법으로 재배한 것을 일반농가(전남 신안, 2015 년산)에서 구입하였고, 설갱(seolgaeng, SG)과 신토흑미 (shintoheukmi, SH)는 농업기술실용화재단(Iksan, Korea) 에서 분양받아 사용하였다. 벼를 가정용정미기(SS-99, Hwang So Farm Machine, Daegu, Korea)를 사용하여 현 미로 도정한 후 냉장 보관(4°C)하며 시료로 사용하였다.
시약
표준품인 thiamine hydrochloride, nicotinic acid, nic- otinamide, folic acid, γ-aminobutyric acid(GABA), gal- lic acid, L-ascorbic acid와 Folin-Ciocalteu’s phenol re- agent와 1,1-diphenyl-2-picrylhydrazyl(DPPH)은 Sig- ma-Aldrich(St. Louis, MO, USA)에서 구입하여 사용하였 다. Potassium ferricyanide, trichloroacetic acid 및 fer- ric chloride는 Wako사(Osaka, Japan)에서 구입하였다. 기 타 사용된 시약은 특급 및 HPLC 등급을 사용하였으며, 3차 증류수는 water purification system(Aqua MaxTM-Ultra, Young Lin Instrument Co., Anyang, Korea)으로 정제된 물을 사용하였다.
현미 발아
현미를 35°C에서 24시간 침지한 후 24시간 동안 plant growth chamber(온도 30°C, 상대습도 90%, DS-52G4P,
Dasol, Hwaseong, Korea)에서 발아하였다. 발아된 현미 (germinated brown rice, GBR)는 drying oven(50°C)에서 수분 함량이 15% 이하가 될 때까지 건조한 후 disc mill(BM- D-100, McCoy Corporation, Seoul, Korea)을 이용해 분 쇄하고 -70°C에서 보관하며 시료로 사용하였다.
발아율 및 초엽장 길이
현미의 발아율은 현미 100개를 기준으로 하였으며 전체 현미에서 싹이 나온 수를 계수하여 발아율을 계산하였다. 최 소 측정단위가 mm인 캘리퍼스(530-101, Mitutoyo, Kawa- saki, Japan)로 현미 싹의 길이를 측정하였다. 초엽장의 길 이가 1 mm 미만인 발아현미는 발아되지 않은 것으로 분류 하여 발아율에서 제외하였으며, 1 mm 이상의 길이로 출하 된 현미만 발아된 것으로 간주하여 초엽장의 길이를 측정하 였다.
발아율
(%) = 싹이 출아된 현미 낟알의 수 전체 현미 낟알의 수, 표본 집단 100개×100
티아민(thiamin) 및 나이아신(niacin) 함량
티아민 및 나이아신 함량은 Kim 등(2014)의 방법을 변형 하여 측정하였다. 시료 3 g을 취한 후 5 mM sodium 1- hexanesulfonate 용액을 25 mL 가하고 혼합하여 초음파 추출기(8893-DTH, Cole-Parmer, Vernon Kills, IL, USA) 로 30분간 추출하였다. 추출액은 원심분리기(HM-150IV, Hanbaek Co., Bucheon, Korea)를 사용하여 374×g에서 10분간 원심분리 하고, 상층액을 취해 0.45 μm syringe fil- ter(Advantec, Tokyo, Japan)로 여과하여 HPLC(1200 Series, Agilent Technologies, Santa Clara, CA, USA)로 분석하였다. HPLC 분석은 컬럼 온도 40°C, 컬럼은 YMC- Pack ODS-AM(C18, 250×4.6 mm, 5 μm, YMC, Kyoto, Japan)을 사용하였다. 검출기는 Photo diode array de- tector(PDA)를 사용하여 파장 270 nm에서 측정하였으며, 이동상 유량은 0.8 mL/min으로 하였다. 이동상 A는 0.75%
acetic acid(v/v)와 0.2% triethylamine(v/v)이 혼합된 5 mM sodium 1-hexanesulfonate, 이동상 B는 methanol을 사용하였으며, 혼합 조건은 이동상 B를 기준으로 0~8 min은 0%, 8~20 min은 0~25%, 20~30 min은 25~40%, 30~31 min은 40~0%, 그리고 31~39 min은 0%로 하였다.
엽산(folate) 함량
엽산 함량은 엽산 농도에 따라 Lactobacillus casei가 생 육하는 성장도를 측정하는 Chun 등(2006)의 방법을 이용하 여 추출 및 분석하였다. L. casei는 실험 당일 depletion me- dia(lactobacilli broth : folic acid casei medium=1:1, v/
v)에 접종하여 37°C(6시간)에서 배양해 사용하였다. Folic acid 용액(2 ng/mL), ascorbic acid 용액(0.1 g/mL) 및 시 료 추출액은 각각 0.45 μm syringe filter로 여과하여 사용 했으며, 시료 추출액은 멸균 증류수를 이용하여 농도에 따라
단계 희석하여 사용했다. Folic acid casei media 배지에 6시간 동안 depletion media에서 배양시킨 L. casei broth 를 접종(5 μL/mL)하고, ascorbic acid 용액(10 μL/mL)을 가해 잘 혼합하여 분석배지로 준비하였다. 96-well micro- plate에 단계 희석한 표준용액과 시료액을 150 μL씩 넣은 후 준비한 분석배지를 150 μL씩 가해 잘 혼합한 후 뚜껑을 덮고 37°C 배양기(HB-103M, Hanbaek Co.)에서 18~20 시간 배양시켰다. Microplate reader(Eon, BioTek Instru- ments, Inc., St. Winooski, VT, USA)를 이용하여 595 nm 에서 표준용액과 시료 농도에 따른 L. casei의 성장 정도를 측정하였다. 표준용액 농도에 따른 L. casei 생장곡선을 이 용하여 시료 중의 엽산 함량을 Gen5 데이터 분석 소프트웨 어(Bio-Tek Instruments, Inc.)를 이용하여 계산했으며, 시 료의 엽산 함량은 μg/100 g으로 나타냈다.
GABA 함량
GABA 함량은 Jo 등(2011)의 방법을 변형하여 측정하였 다. 시료 5 g에 증류수 25 mL를 가하고 homogenizer(T10, IKA, Janke & Kunkel-Str., Staufen, Germany)를 이용하 여 2분간 추출한 후 증류수 5 mL를 homogenizer probe를 세척하여 총 추출액으로 혼합하였다. 추출물을 8,184×g로 4°C에서 10분간 원심분리(Supra R30, Hanil Co., Incheon, Korea) 하여 상층액을 100 mL 용량플라스크에 취하였다.
침전물에 증류수 35 mL를 가한 후 반복해서 추출하여 100 mL가 되도록 정용하였다. 추출액을 잘 혼합한 후 1 mL를 취하여 374×g에서 10분간 원심분리 한 다음 상등액을 0.45 μm syringe filter로 여과하였다. AccQ-Tag Amino Acid Analysis 방법(Waters Co., Milford, MA, USA)에 따라 여 과액을 유도체화하여 HPLC(Agilent Technologies)로 분 석하였다. HPLC 분석에는 ZORBOX Eclipse XDB C18 (150×4.6 mm, 5 μm, Agilent Technologies) 컬럼을 이용 하였다. 검출기는 Fluorescence detector(FLD)를 사용했 으며, 여기파장은 340 nm, 측정파장은 435 nm에서 검출하 였다. 이동상 유량은 1 mL/min이었고 이동상 A 40 mM NaH2PO4(pH 7.8)와 이동상 B acetonitrile : methanol : water=4.5:4.5:1(v/v/v)을 사용했으며, 이동상 B를 기준으 로 다음과 같은 혼합조건을 이용하여 분리했다: 0~3.8 min, 0%; 3.8~28 min, 0~50%; 28~36 min, 50~100%; 36~44 min, 100~0%; 44~50 min, 0%.
유리당 함량
유리당 함량은 Wilson 등(1981)의 방법에 준하여 분석하 였다. 시료 2 g에 증류수를 가해 교반한 후 100 mL로 정용하 여 원심분리(3,000×g, 30 min) 하고 상층액을 취하여 여과 (Whatman No.2, GE Healthcare Life Science, Maidstone, UK)하였다. 여과액은 Sep-pak C18로 정제시킨 다음, 0.45 μm membrane filter(Millipore Co., Billerica, MA, USA) 로 여과 후 HPLC를 이용해 분석하였다. 분석 컬럼은 car-
bohydrate column(250×4.6 mm, Alltech Co., Lexing- ton, KY, USA)이었고, 검출기는 Evaporative Light Scat- tering Detector(ELSD, Agilent Technologies)를 사용했 으며, 이동상 및 유량은 75% acetonitrile, 1 mL/min이었다.
당화 특성 조사
발아현미의 당화 특성은 당화액을 제조하여 당도를 측정 하였다. 발아현미 6 g에 증류수 25 mL를 혼합한 후 진탕항 온수조(HB-205SW, Hanbaek Co.)에서 다음과 같이 온도 변화를 주었다: 45°C(20분), 60°C(25분), 65°C(80분), 72
°C(15분), 78°C(5분). 당도계(PR-101, Atago, Tokyo, Ja- pan)를 이용했으며, 3 반복하여 평균값을 계산하였다.
통계처리
실험결과는 SPSS(Statistics Package for the Social Science, ver. 22.0 for window, SPSS Inc., Chicago, IL, USA) 프로그램을 이용하여 분석했으며, 평균과 표준편차를 구하고 Duncan 다중범위검정(Duncan’s multiple range test)과 Student’s t-test를 이용하여 P<0.05 수준에서 유 의성을 검정하였다.
결과 및 고찰
발아율 및 초엽장 길이
현미 품종별 발아율과 초엽장 길이를 측정한 결과는 Table 1과 같다. 현미의 발아율은 81.6~87.4% 범위로 유의적으로 차이가 관찰되지 않았으나, 현미의 발아율이 84.3%로 보고 된 Moongngarm과 Saetung(2010)의 연구와 동일한 결과 를 나타냈다. 발아된 싹의 길이를 나타내는 초엽장의 길이 분포율에서는 유의적인 차이를 나타냈다(P<0.05). 현미 품 종 중에서 새일미와 설갱은 초엽장의 길이가 1~2 mm 이내 로 짧은 것이 83.0%와 87.4%였고, 초엽장의 길이가 4 mm 이상 긴 현미는 관찰되지 않았다. 반면 신토흑미 픔종의 경 우 초엽장이 1~4 mm를 보이는 발아현미가 고루 분포되어 발아현미 중에서 약 55.4%를 차지하고 있었으며, 새일미나 설갱 현미에서는 관찰되지 않았던 4 mm 이상 긴 초엽장을 가진 발아현미는 약 26.2%를 차지하는 것으로 나타났다.
현미의 발아력은 품종 특성, 수확 후 저장 조건, 여러 환경 요인에 따라 영향을 받을 수 있으며(Ching, 1972; Hoshi- kawa, 1989; Lee 등, 1999), 도정 시 배(embryo)의 물리적 손상 등이 큰 영향을 주는 것으로 알려져 있다. 이는 벼를 현미로 도정 시 왕겨가 제거되며 외기에 직접 노출되어 산화 가 진행되기 쉬우며, 발아에 필요한 효소들이 활성화되어 가수분해 되기 때문에 발아력이 상실되는데, 특히 lipoxy- dase, lipase와 같은 효소의 작용은 저장 중 고미취를 생성 하거나 산도의 증가를 가져오는 요인이 될 수 있는 것으로 알려져 있다(Luh 등, 1991; Yusumatsu와 Moritaka, 1964, Yusumatsu 등, 1965). Jeong 등(2018)의 연구에서 도정하
Table 1. Sprout length and germination rate of brown rice Sprout length of germinated
brown rice (mm)
Germinated brown rice1)
Saeilmi Seolgaeng Shintoheukmi
None (<1) 1∼2 2∼3 3∼4 4∼5 5∼6 6∼7
>7
14.6±4.3Ab3) 83.0±4.4Aa 1.4±0.9Bc 1.0±1.0Bc
0Bc 0Bc 0Bc 0Bc
12.6±4.5Ab 87.4±4.5Aa
0Bc 0Bc 0Bc 0Bc 0Bc 0Bc
18.4±5.4Aab 23.8±10.9Ba 17.0±5.6Aabc 14.6±2.6Abcd 10.8±5.0Acde 7.4±2.5Adef 5.2±2.7Aef 2.8±2.3Af Total germination ratio (%)2) 85.4±4.3A 87.4±4.5A 81.6±5.4A
1)Samples were germinated at 30°C for 24 h after steeping in water at 35°C for 24 h.
2)Germination ratio (%)=(Number of germinated brown rice/ Total number of brown rice)×100.
3)Values with different small letters in the same column are significantly different by Duncan’s multiple range test (P<0.05). Values with different capital letters in the same row are significantly different by Duncan’s multiple range test (P<0.05).
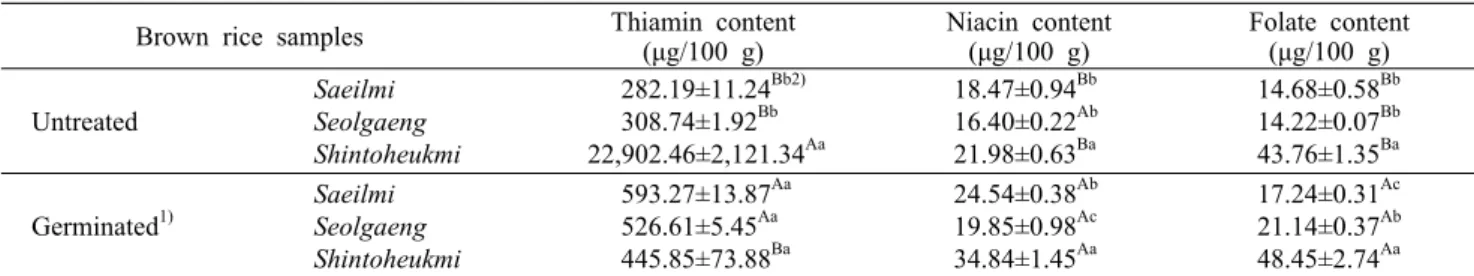
Table 2. Thiamin, niacin, and folate contents of brown rice germinated at 30°C for 24 h (dry basis) Brown rice samples Thiamin content
(μg/100 g) Niacin content
(μg/100 g) Folate content (μg/100 g)
Untreated
Saeilmi Seolgaeng Shintoheukmi
282.19±11.24Bb2) 308.74±1.92Bb 22,902.46±2,121.34Aa
18.47±0.94Bb 16.40±0.22Ab 21.98±0.63Ba
14.68±0.58Bb 14.22±0.07Bb 43.76±1.35Ba
Germinated1)
Saeilmi Seolgaeng Shintoheukmi
593.27±13.87Aa 526.61±5.45Aa 445.85±73.88Ba
24.54±0.38Ab 19.85±0.98Ac 34.84±1.45Aa
17.24±0.31Ac 21.14±0.37Ab 48.45±2.74Aa
1)Germinated at 30°C for 24 h after steeping in water at 35°C for 24 h.
2)Values with different small and capital letters in the same column are significantly different by variety (Duncan’s test, a>b>c) and germination (t-test, A>B), respectively (P<0.05).
지 않은 새일미 품종의 벼 발아에서 1~2 mm 초엽장이 52.0
%로 현미(83.0%)에 비해 낮게 나타났으며, 4~6 mm까지 고르게 자란 것으로 보고되어 있다. 또한 총 발아율도 현미 의 경우 85.4%인 것에 비해 벼는 94.0%로 높게 나타났다.
벼를 현미로 도정하여 상온 저장하며 발아율을 측정할 경우 3주가 지나면 발아율이 현저히 감소하다 4주 이상이 되었을 때는 80%도 못 미친다고 보고되어 있다(Kim 등, 2001). 따 라서 발아에 따른 초엽장 길이 및 발아율의 차이는 실험에 사용된 현미의 품종 및 상태에 의한 영향인 것으로 판단된 다.
티아민, 나이아신 및 엽산 함량
품종별 현미와 발아현미의 티아민, 나이아신 및 엽산 함 량을 분석한 결과는 Table 2와 같다. 품종에 따라 현미의 수용성 비타민 함량이 다르게 나타났는데, 특히 신토흑미에 서 티아민과 엽산 함량이 두드러지게 높게 나타났다. 발아 전후로 티아민 함량은 새일미와 설갱에서 약 2배 정도 높아 졌으나, 신토흑미에서는 1/5 수준으로 급격히 감소하였다.
한편 나이아신과 엽산 함량은 모든 품종에서 발아에 의해 다소 증가한 것으로 나타났다. Mo 등(2006)은 48시간 침지 후 48시간 동안 현미 발아 시 40시간이 경과하면 비타민 B2는 증가하나 티아민과 나이아신은 모두 감소한다고 보고 한 바 있다. 하지만 Moongngarm과 Saetung(2010)은 12시
간 상온에서 침지 후 28~30°C(90~95% 상대습도)에서 24 시간 동안 벼 발아 시 티아민의 경우 유의적인 차이를 보이 지 않았으나, 나이아신의 경우 유의적으로 높아졌다고 보고 한 바 있다. Jeong 등(2018)의 연구에서 침지시간이 길어짐 에 따라 티아민 함량은 유의적인 차이를 보이지 않았으나, 나이아신 함량이 유의적으로(P<0.05) 낮아지는 것으로 보 고되어 있다. 이렇게 다른 결과가 나타나는 원인은 침지시간 에 따라 수용성 비타민이 용출되는 정도가 다르기 때문으로 보인다. 하지만 본 연구에서는 발아로 인하여 신토흑미의 티아민 함량을 제외한 세 품종 모두에서 티아민, 나이아신 및 엽산 함량이 증가하는 것으로 나타났다.
GABA 함량
품종별 현미와 발아현미의 GABA 함량 변화는 Table 3과 같다. GABA는 발아현미의 많은 영양소 중에서 주목받는 물질로 많은 연구가 진행되었다. GABA는 혈중 콜레스테롤 증가 억제, 비만 방지, 시력 증진, 혈압상승억제 등 다양한 생리적인 메커니즘 조절에 관여하는 기능성 성분으로 식품 가공 중 일어나는 변화에 관심이 높아지고 있다(Leventhal 등, 2003; Zhang 등, 2007). 곡류의 발아와 같이 생물 성장 에 의해 GABA가 생성되는 현상 이외에도 녹차 가공이나 전통 된장의 숙성 과정에서도 GABA 함량이 증가한다는 연 구가 보고되어 있어(Chang 등, 1992; Jo 등, 2011) 식품산
Table 3. The GABA contents of untreated and germinated brown rice at 30°C for different time
Brown rice samples GABA content (mg/100 g)
Untreated
Saeilmi Seolgaeng Shintoheukmi
3.84±0.16Bb2) 5.08±0.33Bb 13.82±1.48Ba
Germinated1)
Saeilmi Seolgaeng Shintoheukmi
6.90±0.32Ac 12.15±0.07Ab 33.86±1.25Aa
1)Germinated at 30°C for 24 h after steeping in water at 35°C for 24 h.
2)Values with different small and capital letters in the same col- umn are significantly different by variety (Duncan’s test, a>b>c) and germination (t-test, A>B), respectively (P<0.05).
Table 4. Free sugar contents of untreated and germinated brown rice Free sugar content
(%)
Untreated brown rice Germinated brown rice1)
Saeilmi Seolgaeng Shintoheukmi Saeilmi Seolgaeng Shintoheukmi
Fructose Glucose Sucrose
−Ab2) 1.08±0.15Bb
−Ba
−Ab 1.15±0.09Bb
−Ba
0.32±0.02Aa 2.83±0.29Ba
−Aa
−Ab 1.50±0.16Ab 0.75±0.20Aa
−Ab 1.49±0.15Ab 0.69±0.18Aa
0.35±0.02Aa 14.38±0.55Aa
−Ab Total 1.08±0.15Bb 1.15±0.09Bb 3.15±0.30Ba 2.25±0.36Ab 2.18±0.18Ab 14.73±0.55Aa
1)Germinated at 30°C for 24 h after steeping in water at 35°C for 24 h.
2)Values with different small and capital letters in the same row are significantly different by variety (Duncan’s test, a>b) and germination (t-test, A>B), respectively (P<0.05).
업에서 다양한 가공의 적용을 통한 GABA 성분의 증가를 기대할 수 있을 것으로 보인다.
현미의 GABA 함량은 3.84~13.82 mg/100 g의 범위를 보였으며 품종 중에서는 신토흑미> 설갱> 새일미의 순으로 GABA 함량이 높았다. 발아현미의 경우 GABA 함량이 모든 품종에서 발아 후 약 2배 정도 높아지는 것으로 나타났다.
An 등(2010)의 연구에서는 침지(18°C, 20시간) 후 30°C에 서 싹의 길이가 3 mm가 되도록 발아시켰을 때 현미, 홍미 및 흑미의 경우 GABA 함량이 2.5~11배 증가했지만, 녹미 에서는 발아 전후에서의 유의적인 GABA 함량 변화가 없는 것으로 나타났다. Oh 등(2012)의 연구에서도 품종에 따라 발아 시 GABA 함량의 증가에 차이가 있음이 확인되었는데 평균적으로 발아에 의해 GABA 함량이 약 2.02~6.77배 증 가했다고 보고하였다. Moongngarm과 Saetung(2010)의 연구도 벼 및 현미가 발아에 의해 GABA 함량이 2.9~4.8배 증가했다고 보고하였다. 본 연구에서도 발아를 통해 GABA 성분이 증가함을 알 수 있었다.
유리당 함량
품종별 현미와 발아현미의 유리당 함량 차이를 분석한 결 과는 Table 4와 같다. 현미와 발아현미에서 fructose, glu- cose 및 sucrose가 확인되었으며, 세 가지 품종 모두에서 glucose 함량이 증가하였으나, 증가폭은 품종에 따라 차이 를 나타내었다. 신토흑미 품종의 경우 발아에 의해 glucose
함량이 큰 폭으로 증가하여 발아 전보다 약 5배 높아진 반면, 새일미와 설갱 품종에서는 glucose 함량의 증가가 미미하 였다.
현미의 유리당 함량이 발아 중에 변화되는 현상은 여러 식물체에서 보고되어 있다. Ko와 Park(1983)은 녹두가 발 아되는 동안 총당이 감소하고 환원당은 증가한다고 하였고, Åman(1979)도 녹두와 이집트콩(chickpea)은 발아 시 poly- saccharides, starch와 같은 고분자 탄수화물의 함량이 감 소하고 glucose, fructose와 같은 단당류의 함량은 증가하 는 경향을 나타냈다고 보고하였다. 또한, 식물종자 발아의 경우 fructose, glucose와 같은 유리당 함량은 발아 초기에 급격히 감소하다가 발아가 진행됨에 따라 다시 증가하는 현 상이 나타난다고 보고된 바 있다(Kim 등, 1988b). 이러한 현상은 발아 초기에는 종자의 성장에 수반되는 대사 작용을 위해 에너지원이나 생합성에 많은 유리당이 필요하기 때문 에 이들의 함량이 감소하지만, 발아가 지속하면서 α-amy- lase의 활성이 증가하게 되고 이와 함께 고분자 탄수화물의 가수분해가 가속화되면서 유리당의 함량 증가가 나타나는 것으로 보인다(Lee 등, 1995a). 본 연구에서는 발아현미에 서 glucose 함량이 증가하였으나, fructose 함량은 유의적 인 차이가 없는 것을 관찰할 수 있었다.
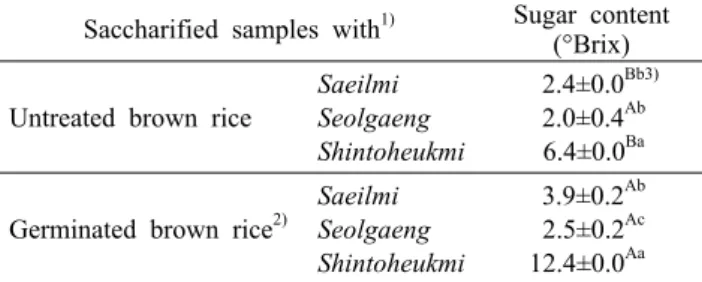
발아현미의 당화력
품종에 따른 현미와 발아현미의 당화력 변화를 측정하기 위해 이들을 일정 조건에서 당화를 진행한 후 당화용액의 당도를 측정한 결과를 Table 5에 나타내었다. 발아 전 현미 에 의한 당화력은 품종에 따라 다르게 나타나 당화액의 당도 가 2.0~6.4°Brix의 범위를 나타냈다. 현미 품종 중에서 특 히 신토흑미가 보이는 당화액의 당도가 가장 높게 나타났다.
이들의 당화력을 발아시킨 현미와 비교한 결과, 설갱의 경우 는 발아에 의해 당화액의 당도가 유의적으로 증가하지 않았 으나, 발아된 새일미와 신토흑미의 경우 당화액의 당도가 발아 전에 비해 약 1.6~1.9배 증가하는 것으로 나타났다.
발아에 의한 현미의 당화력 증가는 쌀 품종별 특성, 발아 온도 및 기간에 영향을 받는 것으로 알려져 있는데, Kim과 Lee(1997)는 발아 온도를 각각 15, 20, 25, 30°C로 처리했 을 때 30°C에서 당화력이 가장 우수하게 나타났다고 보고한 바 있다. 한편 Kim과 Lee(1997)는 동일 온도에서 벼보다는
Table 5. Saccharification power of untreated and germinated brown rice
Saccharified samples with1) Sugar content (°Brix)
Untreated brown rice
Saeilmi Seolgaeng Shintoheukmi
2.4±0.0Bb3) 2.0±0.4Ab 6.4±0.0Ba
Germinated brown rice2)
Saeilmi Seolgaeng Shintoheukmi
3.9±0.2Ab 2.5±0.2Ac 12.4±0.0Aa
1)Saccharified samples were obtained by incubation of brown rice in water at 45°C for 20 min, at 60°C for 25 min, at 65°C for 80 min, at 72°C for 15 min, and at 78°C for 5 min.
2)Germinated at 30°C for 24 h after steeping in water at 35°C for 24 h.
3)Values with different small and capital letters in the same col- umn are significantly different by variety (Duncan’s test, a>b>c) and germination (t-test, A>B), respectively (P<0.05).
현미의 발아속도가 빠르고 이는 왕겨의 제거로 겨층이 노출 되어 환경과의 상호반응이 빨라졌기 때문이라고 보고하였 으며, 당 함량도 벼보다는 현미에서 높게 나타난다고 보고하 였다. 하지만 본 연구의 침지 및 발아조건과 동일한 조건에 서 새일미를 연구한 선행 연구 결과(Jeong 등, 2018)와 비 교해볼 때 발아벼는 9.7°Brix, 발아현미는 6.9°Brix의 당 함량을 보여 같은 품종의 발아벼가 발아현미보다 높은 당 함량을 보여 Kim과 Lee(1997)와는 상반된 결과를 나타냈 다. 이러한 차이는 벼의 품종별 특성과 연구된 침지 및 발아 조건에 영향을 받을 수 있는 것으로 보인다.
최근 장류, 주류 등의 산업에서 제품의 품질을 개선하기 위해 보리를 발아시킨 맥아를 대신하여 발아미를 사용해 가 공식품을 제조하는 시도가 이루어지고 있다. 본 연구에서 제조된 세 가지 발아현미의 유용성분의 변화와 당화 특성에 대한 기초자료는 식품가공 산업에서의 활용도를 검토할 수 있는 기초자료로 활용될 수 있을 것이다.
요 약
세 가지 현미 품종(새일미, 설갱, 신토흑미)과 이를 30°C에 서 24시간 침지 후 35°C(90% 상대습도)에서 24시간 발아 시킨 발아현미의 유용성분 및 당화 특성을 비교하여 식품산 업에서 이들의 활용도를 높이기 위한 기초자료로 제공하고 자 하였다. 발아 후 새일미, 설갱, 신토흑미 세 품종은 모두 초엽장 길이가 각각 4, 2, 7 mm 이하의 범위를 나타내어 품종별 특성의 차이를 보였다. 특히 새일미와 설갱은 1~2 mm의 짧은 초엽장이 각각 83%와 87%로 발아현미의 대부 분을 차지하였으나, 신토흑미 품종의 경우는 1~2 mm의 짧 은 초엽장이 약 24%의 수준이었고 대부분의 발아현미가 긴 초엽장을 나타냈다. 발아에 의한 티아민 함량은 신토흑미의 경우 감소하였으나, 새일미와 설갱 품종에서는 증가하는 것 으로 관찰되었다. 현미의 나이아신, 엽산 및 GABA 함량은
발아에 의해 세 품종 모두 증가하였다. Glucose 함량은 현미 세 품종 모두에서 발아에 의한 증가가 확인되었으며, su- crose 함량은 새일미와 설갱에서만 증가하는 것으로 나타 났다. 현미의 당화력은 새일미와 신토흑미 품종에서만 증가 하였는데, 특히 초엽장의 길이가 고르게 분포된 신토흑미에 서 2배 높아졌다. 본 연구에서는 쌀 품종에 따라 차이를 보이 지만 현미 발아 시 GABA와 같은 기능성 성분과 일부 수용 성 비타민 함량이 높아질 수 있으며, 발아를 위한 효소 활성 증가로 인해 당화력이 개선되는 것을 확인할 수 있었다. 본 연구 결과는 발아현미 자체로서의 활용뿐만 아니라 당화력 이 필요한 맥주와 장류 등의 식품산업 분야에서도 유용한 자료로 활용될 수 있을 것으로 보인다.
감사의 글
본 연구는 2018년도 식품의약품안전처의 연구개발비(17162 MFDS082)로 수행되었으며 이에 감사드립니다.
REFERENCES
Åman P. Carbohydrates in raw and germinated seeds from mung bean and chick pea. J Sci Food Agric. 1979. 30:869-875.
An MK, Ahn JB, Lee SH, Lee KG. Analysis of γ-aminobutyric acid (GABA) content in germinated pigmented rice. Korean J Food Sci Technol. 2010. 42:632-636.
Cha MN, Jun HI, Song GS, Kim YS. The effects of germination conditions on GABA and the nutritional components of barley.
Korean J Food Sci Technol. 2012. 44:41-47.
Chang JS, Lee BS, Kim YG. Changes in γ-aminobutyric acid (GABA) and the main constituents by a treatment conditions and of anaerobically treated green tea leaves. Korean J Food Sci Technol. 1992. 24:315-319.
Ching TM. Metabolism of germinating seeds. In: Kozlowski TT, editor. Seed Biology: Germination Control, Metabolism, and Pathology. Academic Press, New York, NY, USA. 1972. Vol
Ⅱ, p 103-218.
Cho D, Chung HJ, Cho HY, Lim ST. Health functions and uti- lization products of germinated brown rice. Food Science and Industry. 2011. 44(1):76-86.
Choi HD, Park YK, Kim YS, Chung CH, Park YD. Effect of pretreatment conditions on γ-aminobutyric acid content of brown rice and germinated brown rice. Korean J Food Sci Technol. 2004. 36:761-764.
Chun J, Martin JA, Chen L, Lee J, Ye L, Eitenmiller RR. A differential assay of folic acid and total folate in foods con- taining enriched cereal-grain products to calculate μg dietary folate equivalents (μg DFE). J Food Compos Anal. 2006. 19:
182-187.
Hoshikawa K. The growing rice plant: an anatomical monograph.
Nobunkyo, Tokyo, Japan. 1989. p 25-50.
Hyeun SK, Kwon YA, Lee SJ. Quality characteristics of brewed beer with rice adjunct. Food Eng Prog. 2012. 16:139-144.
Jeong BG, Choi KS, Chun J. Physicochemical properties of Saeil- mi (Oryza sativa Linne) germinated with different steeping and germination time. Korean J Food Preserv. 2018. 25:311- 320.
Jo SJ, Hong CO, Yang SY, Choi KK, Kim HK, Lee KW.
Changes in contents of γ-aminobutyric acid (GABA) and iso-
flavones in traditional Korean Doenjang by ripening periods.
J Korean Soc Food Sci Nutr. 2011. 40:557-564.
Jung HY, Lee DH, Baek HY, Lee YS. Pre- and post-germination changes in pharmaceutical compounds of germinated brown rice. Korean J Crop Sci. 2008. 53(S):37-43.
Kang MY, Lee YR, Nam SH. Characterization of the germinated rices to examine an application potentials as functional rice processed foods. Korean J Food Sci Technol. 2003. 35:696- 701.
Kim DJ, Oh SK, Yoon MR, Chun A, Choi IS, Lee DH, et al.
The change in biological activities of brown rice and germi- nated brown rice. J Korean Soc Food Sci Nutr. 2011. 40:781- 789.
Kim GP, Lee J, Ahn KG, Hwang YS, Choi Y, Chun J, et al.
Differential responses of B vitamins in black soybean seeds.
Food Chem. 2014. 153:101-108.
Kim HY, Lee SH, Hwang IG, Kim TM, Park DS, Jeong HS, et al. Antioxidant activity and anticancer effects of rough rice (Oryza sativa L.) by germination periods. J Korean Soc Food Sci Nutr. 2012. 41:14-19.
Kim IS, Kwon TB, Oh SK. Study on the chemical change of general composition, fatty acids and minerals of rapeseed dur- ing germination. Korean J Food Sci Technol. 1988a. 20:188- 193.
Kim IS, Kwon TB, Oh SK. Study on the compositional change of free sugars and glucosinolates of rapeseed during germina- tion. Korean J Food Sci Technol. 1988b. 20:194-199.
Kim SL, Son YK, Son JR, Hur HS. Effect of germination con- dition and drying methods on physicochemical properties of sprouted brown rice. Korean J Crop Sci. 2001. 46:221-228.
Kim SS, Kim SY, Lee WJ. Microwave vacuum drying of germi- nated brown rice as a potential raw material for enzyme food.
Korean J Food Sci Technol. 1998. 30:1107-1113.
Kim SS, Lee WJ. Characteristics of germinated rice as a poten- tial raw material for Sikhe production. Korean J Food Sci Technol. 1997. 29:101-106.
Ko MS, Park BH. Changes of sugar contents of mung bean dur- ing germination. Korean J Food Nutr. 1983. 12:236-239.
Korea Agro-Fisheries & Food Trade Corporation. Processed food market-liquor. Naju, Korea. 2016.
Lee MH, Son HS, Ju JS, Oh SK, Kwon TB. Change in α-amy- lase activity and free sugar contents of buckwheat during ger-
mination. Korean J Food Nutr. 1995a. 8:32-36.
Lee WJ, Cho MK, Chung KM. Quality characteristics of Korean rice as brewing adjunct. Korean J Food Sci Technol. 1995b.
27:516-519.
Lee YT, Seo SJ, Chang HG. Quality characteristics of barley varieties related to enzymatic activity in malt. Korean J Food Sci Technol. 1999. 31:1421-1426.
Leventhal AG, Wang Y, Pu M, Zhou Y, Ma Y. GABA and its agonists improved visual cortical function in senescent mon- keys. Science. 2003. 300:812-815.
Luh BS, Barber S, Benedito de Barber C. Rice bran: chemistry and technology. In: Rice. Springer, Boston, MA, USA. 1991.
p 313.
Mo KH, Choi Y, Choi SG, Lee J. The change of some com- pounds in brown rice germinated by filtrate of loess suspen- sion. J Agric Life Sci. 2006. 40(2):41-48.
Moongngarm A, Saetung N. Comparison of chemical composi- tions and bioactive compounds of germinated rough rice and brown rice. Food Chem. 2010. 122:782-788.
Oh SH. Effects and applications of germinated brown rice with enhanced levels of GABA. Food Science and Industry. 2007.
40(3):41-46.
Oh SK, Lee JH, Yoon MR, Kim DJ, Lee DH, Choi IS, et al.
Physiochemical properties of germinated brown rice. J Kore- an Soc Food Sci Nutr. 2012. 41:963-969.
Statistics Korea. 2018 annual grain consumption per capita.
2019 [cited 2019 Aug 6]. Available from: http://kosis.kr/stat Html/statHtml.do?orgId=101&tblId=DT_1ED0001&conn_pa th=I2
Wilson AM, Work TM, Bushway AA, Bushway RJ. HPLC deter- mination of fructose, glucose, and sucrose in potatoes. J Food Sci. 1981. 46:300-301.
Yasumatsu K, Moritaka S, Bichu S, Ishii K, Shimazono H, Fujita E. Studies on cereals (Ⅱ). Flavor deterioration of polished rice during storage. J Japan Soc Food Nutr. 1965. 18:130-133.
Yasumatsu K, Moritaka S. Fatty acid compositions of rice lipid and their changes during storage. Agric Biol Chem. 1964.
28:257-264.
Zhang H, Yao HY, Chen F, Wang X. Purification and character- ization of glutamate decarboxylase from rice germ. Food Chem.
2007. 101:1670-1676.