Ziyuglycoside II Attenuates Tumorigenesis in Experimental Colitis-associated Colon Cancer
Hye-Jin Cheon and Jin-Kyung Kim*
Department of Biomedical Science, Daegu Catholic University, Gyeongsan-si 38430, Korea Received July 5, 2019 /Revised July 30, 2019 /Accepted July 31, 2019
Colorectal cancer is a major health problem in industrialized countries. Ziyuglycoside II (3β-3-α-1- ara- binopyranosyloxy-19-hydroxyurs-12-en-28-oicacid), a triterpenoid saponin isolated from the roots of Sanguisorba officinalis L., possesses antioxidant, antiangiogenic, and anticancer properties. However, the therapeutic function of ziyuglycoside II in colitis-associated colorectal carcinogenesis is undefined. In the present study, the effect of ziyuglycoside II on colitis-associated colon cancer induced in mice us- ing azoxymethane (AOM)/dextran sulfate sodium (DSS) was explored. The AOM model recapitulates many features of human colon cancer, but it lacks an inflammatory component. DSS induces colitis and promotes AOM-induced colon cancer in mice. BALB/c mice were injected with AOM and ad- ministered 2% DSS in drinking water. The mice were given ziyuglycoside II (1 or 5 mg/kg) orally three times per week, and colonic tissue was collected at 64 days. Administration of ziyuglycoside II markedly diminished the formation of colonic tumors. Western blot and immunohistological analyses showed that ziyuglycoside II noticeably decreased nuclear factor kappa-B-positive cells and levels of inflammation-related proteins, such as inducible nitric oxide synthase, cyclooxygenase-2, tumor ne- crosis factor-α, and interleukin-6 in colon tissue. It also prompted apoptosis. Ziyuglycoside II treat- ment augmented cleaved forms of caspase-3, caspase-7, and poly (ADP-ribose) polymerase in colonic tissues. In conclusion, ziyuglycoside II could defend against colitis-associated tumorigenesis in mice by inhibiting inflammation and inducing apoptosis. This shows a promising chemopreventive poten- tial for its use in colitis-associated colon cancer.
Key words : Apoptosis, colitis-associated colon cancer, inflammation, NF-κB, ziyuglycoside II
*Corresponding author
*Tel : +82-53-850-3774, Fax : +82-53-850-3774
*E-mail : [email protected]
This is an Open-Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Life Science 2019 Vol. 29. No. 9. 941~948 DOI : https://doi.org/10.5352/JLS.2019.29.9.941
Introduction
Colorectal cancer is the third most prevalent malignancy worldwide and is the fourth most-common cause of can- cer-related death [4]. More than one million new cases of colorectal cancer are reported annually, and the incidence rate has been increasing. In 2012, nearly 10% of the total worldwide cancers, 1,361,000 new cases, were reported, and nearly half died from colorectal cancer [25]. Common risk factors for colorectal cancer include genetic background and environmental risk factors, such as diabetes, cholecystec- tomy, obesity, and high-fat diets [1]. Among these factors, long-standing inflammatory bowel disease (IBD) is a higher risk for developing colorectal cancer [3, 9, 11]. The cumu-
lative risk for developing colorectal cancer in extensive IBD is a 19-fold increase over that of the general population, which also suggests that chronic intestinal inflammation is a predisposing condition for colorectal cancer [3].Indeed, colonic inflammation can simply initiate tumorigenesis and endorse carcinogenesis [9]. For example, numerous im- mune-cell penetrates in the colon enrich the environment for reactive oxygen species and reactive nitrogen species, which induce DNA impairment and accelerate the cancer initiation [9].Furthermore, inflammatory cells produce large amounts of pro-tumorigenic cytokines, including tumor ne- crosis factor (TNF)-α and interleukin (IL)-6, which shove tu- mor progression [7].
Phytochemicals have been drawing increasing attention for cancer prevention because of their chemical diversity, structural complexity, intrinisic biological activity, simple obtainability, affordability, lack of toxic effects, and ability to control a variety of signal transduction pathways and cell processes [8]. In fact, we demonstrated several laboratory- based studies of the antitumor effects of bioactive phyto- chemicals [12-14, 23]. Ziyuglycoside II (3β-3-α-1-arabinopyr-
A B
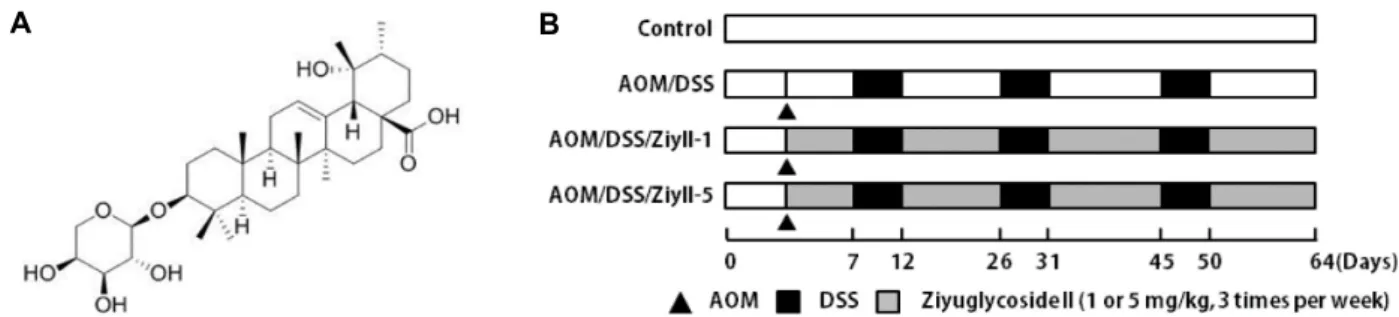
Fig. 1. Experimental protocol for evaluating the effects of ziyuglycoside II on AOM/DSS-induced colon cancer in mice. (A) The chemical structure of ziyuglycoside II. (B) Experimental protocol for the development of a colitis-associated colon carcino- genesis model in mice. Mice were injected with AOM once. After one week, the mice were administered with 2% DSS in drinking water for five days, followed by two weeks of regular drinking water for recovery. This cycle was repeated three times.
anosyloxy-19-hydroxyurs-12-en-28-oicacid) is a triterpenoid saponin isolated from the roots of Sanguisorba officinalis [17].
S. officinalis (Great Burnet) is a prolonged flowering per- ennial herb of the Rosaceae family that cultivates in grass- land environments, wet fields and grasslands in colder areas of Europe, North America and Asia [17]. The extracts of S.
officinalis are the basis of popular folk medicines, particularly in China, for the treatment of metabolic and inflammatory conditions such as diarrhea, chronic intestinal infections, du- odenal ulcers and bleeding [28].In addition, our recent study showed that ziyuglycoside II blocked various characteristics of angiogenesis, which might support to its antitumor effects [20].These previous investigations encouraged us to test the therapeutic effects of ziyuglycoside II on inflammation-re- lated colon cancer. To investigate the in vivo antitumor effect of ziyuglycoside II, we selected a mouse model of azoxy- methane (AOM) /dextran sodium sulfate (DSS)-induced col- itis-associated colon cancer.
Materials and Methods
Animals
Five-week-old female BALB/c mice were supplied by the Hyochange Science (Daegu, Korea). They were housed in the Animal Resource Facility, Daegu Catholic University, in accordance with the university’s Institutional Animal Care and Use Committee guidelines. All mice were maintained under standard conditions (12 hr light / 12 hr dark, 22 ±
1℃, 55±5% relative humidity). All experiments were per- formed in accordance with the relevant laws and approved by the Institutional Animal Care and Use Committee of Daegu Catholic University with the approval/protocol num- ber IACUC-2017-050.
Chemicals and reagents
All reagents were acquired from Sigma-Aldrich (St. Louis, MO) unless otherwise specified. Ziyuglycoside II and DSS (MW 40,000-50,000) were obtained from Avention Chemical (Incheon, Korea) and ICN Biochemicals Inc. (Aurora, OH, USA), respectively.
Induction of colitis-associated colon cancer and treatment procedure
Following an acclimation period, the mice were arbitrarily and equally distributed into four groups, with eight mice per group. Mice were injected intraperitoneally with 10 mg/
kg AOM (Sigma-Aldrich) at six weeks of age. Seven days later, 2% DSS was administered in the drinking water for five days, followed by regular tap water for 14 days. The cycle of DSS and regular water was repeated three times.
Normal mice, which were not administered with AOM/DSS, served as a negative control group. An AOM/DSS-only group served as a positive control. Mice were given ziyugly- coside II for 1 and 5 mg/kg of body weight three times a week via gastric intubation using an oral Zonde needle. The overall experimental schedule is depicted in Fig. 1B.
Body weights of mice were documented every week. At the end of the experiment, the mice were sacrificed, and co- lon tissues were collected. The colons were cut open longi- tudinally along the main axis and washed with phosphate buffered saline (PBS, pH 7.4). The number of tumors was counted. Consequently, a remained colon tissues were fixed in 4% paraformaldehyde buffer for further analysis. For his- topathological analysis, following fixation and embedding in paraffin, blocks were sectioned and stained with hematox- ylin and eosin (H&E).
Immunohistochemistry (IHC)
Paraffin-embedded colonic sections (5 μm) were dewaxed, boiled in citrate buffer for antigen retrieval, and stained overnight with anti-nuclear factor kappa-B (NF-κB) p65 anti- body (Cell Signaling Technology, Beverly, MA, USA) at a 1:100 dilution at 4℃. Sections were managed sequentially with biotin-conjugated secondary antibody and streptavi- din-horseradish peroxidase (DAKO, Carpinteria, CA) and stained using a DAB Kit (DAKO). Subsequently, the samples were counterstained with hematoxylin. Apoptotic cells in co- lonic tissues were identified with the terminal deoxy- ribonucleotidyl transferase-mediated dUTP-digoxigenin nick end-labeling (TUNEL) method using an In Situ Apoptosis Detection Kit (R&D Systems, Minneapolis, MN, USA) in ac- cordance with the manufacturer’s instructions. Negative control sections were operated similarly but with the ex- clusion of the primary antibodies. Images were acquired with a DM500 microscope (Leica Biosystems Richmond Inc., Richmond, IL, USA) and quantified by counting the number of positively stained cells in ten randomly selected fields.
The number of positively stained cells was expressed as the percent of total cells in the ten fields.
Western blot analysis
Colon tissue samples were homogenized in a Pro-prep protein-extraction solution (iNtRon, Sungnam, Korea) con- taining 1x protease-inhibitor cocktail (Roche, Indianapolis, IN, USA). The concentrations of protein were determined using a BCA protein assay kit (Thermo Fisher Scientific, Waltham, MA, USA). Colonic homogenates were separated by 10% SDS-polyacrylamide gel electrophoresis, and the re- solved proteins were transferred to polyvinylidene fluoride membranes (Roche, Basel, Switzerland). Membranes were blocked in PBS containing Tween (PBST) and 3% skim milk for 1 hr at room temperature. After washes in PBST, the membranes were incubated with following antibodies: an- ti-inducible nitric oxide synthase (iNOS) mouse monoclonal (BD Bioscience, Becton Dickinson, USA), anti-cyclooxygenase (COX)-2 rabbit polyclonal (Cayman Chemical, Ann Arbor, MI, USA), anti-TNF-α mouse monoclonal (Thermo Fisher Scientific), anti-IL-6 goat polyclonal (Thermo Fisher Scientific), anti-poly ADP-ribose polymerase (PARP) rabbit monoclonal (Cell Signaling Technology), anti-cleaved caspase-3 rabbit polyclonal (Cell Signaling Technology), anti-cleaved cas- pase-7 rabbit monoclonal (Cell Signaling Technology) and anti-β-actin mouse monoclonal (Sigma). After overnight in-
cubation at 4℃, the secondary horseradish peroxidase-con- jugated antibodies were employed and developed using the ECL Western blot detection reagents (GE Healthcare, Little Chalfont, UK). Densitometric data from western blots were analyzed using Total Lab software (Davinch-K, Seoul, Korea).
Statistical analysis
The results were analyzed by GraphPad Prism 6.0 soft- ware (GraphPad Software Inc., San Diego, CA, USA) and are presented as the mean ± SEM. Statistically significant values were evaluated by one-way analysis of variance with Bonferroni multiple comparison post tests. Differences with p<0.05 were considered statistically significant.
Results
Ziyuglycoside II administration decreases AOM/
DSS-induced tumorigenesis in mice
The colitis-associated colon cancer model was made by intraperitoneal injection of AOM followed by three cycles of DSS exposure as shown in Fig. 1B. The changes of body weight did not differ between the groups during the experi- ment (Fig. 2A), indicating that ziyuglycoside II does not have any toxic effect. As expected, several colonic tumors were seen in all mice receiving AOM/DSS (Fig. 2B, Fig. 2C). A high dose of ziyuglycoside II (5mg/kg) treatment sig- nificantly reduced the incidence and size of lesions (Fig. 2B, Fig. 2C). Correspondingly, mean tumor load, which was de- fined as the sum of diameters of all tumors in a certain mouse, was reduced in 5 mg/kg ziyuglycoside II-treated mice (Fig. 2C, Fig. 2D). It was also observed that colon length challenged by AOM/DSS was shorter than that in the con- trol group as shown in Fig.2B and 2E. In mice to which ziyu- glycoside II (5 mg/kg) was administered, the colon length was restored slightly without statistical significances (Fig.
2E). In contrast, the colon weight of AOM/DSS mice was heavier than that in the control or 5 mg/kg ziyuglycoside II-given mice (Fig. 2F).
Macroscopically, nodular and polypoid colonic tumors were noticed in the middle and distal colon of all AOM/DSS treated mice, as shown in Fig. 2B. To analyze the pathology of AOM/DSS-induced tumors, we performed H&E staining of colon tissue. H&E stained sections of the colons of mice at the end of the study are shown in Fig. 3. AOM/DSS treat- ed mice had histological alteration of the colonic mucosa, including inflammation with mucosal ulcer, infiltration of
A B
C D E F
Fig. 2. Ziyuglycoside II treatment ameliorated tumor load in AOM/DSS mice. (A) Body weight changes of all groups after treatment.
(B) Representative macroscopic features of colon in mice after anatomy. Colon was opened longitudinally after sacrifice and (C) the number of tumors and (D) area were calculated. (E) Colon length and (F) colon weight after AOM/DSS induction at day 64. Results are expressed as means ± SEM. *p<0.05, **p<0.01, ***p<0.001 compared with AOM/DSS-treated mice.
Fig. 3. Ziyuglycoside II treatment suppressed colitis-associated tumorigenesis. Hematoxylin and eosin (H&E) staining of colon tumor in the control, AOM/DSS, and AOM/DSS/ ziyuglycoside II groups (upper, 10× objective; middle, 40× objective; lower, 100×
objective; bar = 100 μm). Middle and lower panel show enlarge images of the region defined by the solid line.
inflammatory cells into the lamina propria, and loss of crypts. The polyps exhibited a high-grade of dysplasia; and were mostly adenoma and adenocarcinoma. However, ziyu- glycoside II treatment, especially in the 5 mg/kg-treated
group, decreased the loss of crypts in the colonic mucosa and relieved the inflammation of the colonic mucosa. These results showed that tumor incidence was suppressed in ziyuglycoside II-treated mice, indicating that ziyuglycoside
A B
C D
Fig. 4. Treatment with ziyuglycoside II inhibited AOM/DSS-induced inflammatory mediators in colon. (A) Western blot was used to detect iNOS, COX-2, TNF-α, and IL-6 in the colon of mice. Representative results are shown and β-actin served as an equal loading control. (B) Detected iNOS, COX-2, TNF-α, and IL-6 proteins from eight mice were quantified by densitometric analysis and normalized to β-actin. Results are expressed as mean ± SEM. *p<0.05, **p<0.01, ***p<0.001, compared with AOM/DSS-treated mice. (C) Immunohistochemical analysis of nuclear p65 expression in the colon tissues (upper, 40× objective;
lower, 100× objective; bar = 100 μm). Lower panel show enlarge images of the region defined by the solid line. (D) The percentage of nuclear p65 positive cells in colon tissues was calculated as described in Methods. The results are expressed as mean ± SEM. ***p<0.001 compared with AOM/DSS-treated mice.
II administration blocked colitis-associated tumor induction.
Ziyuglycoside II treatment reduces inflammatory responses
Because the AOM/DSS-induced colon cancer model is linked with inflammation, we tested whether the antitumor action of ziyuglycoside II was related with anti-inflamma- tory effects via regulation of pro-inflammatory mediators, such as cytokines and enzymes, in colonic tissue. The protein levels of the pro-inflammatory cytokines, TNF-α and IL-6, and the inflammatory enzymes, iNOS and COX-2, were sig- nificantly raised in AOM/DSS-treated mice, more than in non-treated control group (Fig. 4A, Fig. 4B). Increased pro- tein levels of pro-inflammatory cytokines, iNOS and COX-2, by AOM/DSS treatment were notably decreased by ziyugly- coside II treatment, especially at the 5 mg/kg concentration.
NF-κB cascades are supposed the essential pathways me- diating the synthesis of numerous inflammatory mediators.
Therefore, we next observed the activation of NF-κB in co- lonic tissues by measuring the nuclear expression of NF-κB.
The level of NF-κB in nuclei was significantly increased in
the AOM/DSS-treated group, and was substantially reduced by ziyuglycoside II treatment as shown in Fig. 4C and Fig.
4D.
Ziyuglycoside II treatment induces apoptosis in colon tissues
To further analyze the mechanism of the antitumor activ- ity of ziyuglycoside II, we analyzed the apoptosis-related protein in colonic tissues by Western blot analysis. The ef- fects of ziyuglycoside II on the cleavage of procaspases-3 and -7 in the caspase cascade and PARP were investigated by Western blot analysis. Ziyuglycoside II stimulated the cleavage of pro-caspase-3 into its two active fragments (17 and 19 kDa) and pro-caspase-7 (Fig. 5A, Fig. 5B). Ziyuglyco- side II also induced the cleavage of PARP, a DNA repair enzyme that is one of the cleavage targets of activated cas- pase-3 (Fig. 5A, Fig. 5B).
In addition, TUNEL staining in colonic crypt cells and tumor epithelia showed that ziyuglycoside II administration induced apoptosis more than in the AOM/DSS group (Fig.
5C, Fig. 5D). The treatment with 5 mg/kg ziyuglycoside II
A B
C D
Fig. 5. Administration of ziyuglycoside II induces apoptosis in the colon. (A) Western blot was used to detect cleaved-PARP, cleaved-caspase3 and cleaved-caspase7 in the colon of mice. Representative results are shown and β-actin served as an equal loading control. (B) Cleaved-PARP, cleaved-caspase3 and cleaved-caspase7 proteins from eight mice were quantified by densi- tometric analysis and normalized to β-actin. Results are expressed as mean ± SEM. *p<0.05, **p<0.01, ***p<0.001, compared with AOM/DSS-treated mice. (C) Apoptosis was determined via TUNEL staining of colon sections from mice (upper, 40×
objective; lower, 100× objective; bar = 100 μm). Lower panel show enlarge images of the region defined by the solid line.
(D) The percentage of TUNEL-positive cells in the colonic tissue was estimated as described in Methods. The results are expressed as mean ± SEM, ***p<0.001, compared with AOM/DSS-treated mice.
in the AOM/DSS mice led to a 21.8% increase of apoptosis in tumor tissue areas over that in the non-treated AOM/DSS mice.
Discussion
Until now, there have been a few studies about the anti- cancer effect of ziyuglycoside II, one of the major efficient components of S. officinalis L. For example, Zhu et al., [31]
demonstrated the anti-proliferative actio of ziyuglycoside II in breast cancer cells, which was facilitated through arresting the cell cycle in the G2/M phase and inducing apoptosis, possibly via controlling the ROS/JNK signaling pathway.
Zhu et al., [30] also showed that apoptosis caused by ziyugly- coside II was accompanied by the activation of the mitochon- drial pathway, in particular a reduced mitochondrial mem- brane potential as well as an elevated Bax/Bcl-2 ratio, cyto- chrome c release, and the activity of caspase-3 and caspase-9.
Another study showed that ziyuglycoside II treatment sup- pressed the proliferation of human gastric cancer cells by
inducing apoptosis but not cell cycle arrest, which was con- nected with control of Bax/Bcl-2 expression, and activation of the caspase-3 pathway [29]. Although these studies have reported its anticancer activities, all studies were carried out in vitro using human cancer cell lines. However, this study is the first to report the antitumor potential of ziyuglycoside II in a mouse model of colitis-associated colon cancer.
Chronic inflammation is assumed to be a driving strength for the development and progression of many human can- cers [15].This is, for example, evident in patients with ulcer- ative colitis or Crohn’s disease. Indeed, the natural history of ulcerative colitis patients, can be marked by the develop- ment of colorectal cancer [2].There is also evidence that the risk of colitis-associated colorectal cancer is severely related to the duration and extension of inflammation [5]. Since col- itis-associated colorectal carcinogenesis is possibly encour- aged by chronic inflammation, regulation of inflammatory responses in the colonic environment are critical for prevent- ing and/or curing colon cancer. Inflammation engages con- nections between various immune cells, inflammatory cells,
cytokines, and inflammatory enzymes, such as COX and iNOS, which may lead to signaling that helps cancer cell propagation and tumor growth. Among inflammatory regu- lators, NF-κB is a key factor in controling inflammatory re- sponses [19, 27]. By potentially stimulating the expression of several pro-inflammatory genes, NF-κB strappingly af- fects the course of mucosal inflammation and tumorigenesis [18]. In the present study, we showed a significant inhibition of NF-κB-positive cells in the nuclei of AOM/DSS-treated mice following exposure to ziyuglycoside II (Fig. 4C, Fig.
4D). These results suggest that the suppressed expression of pro-inflammatory cytokines and enzymes in ziyuglyco- side II-administered AOM/DSS treated colonic tissue might be attributed to inhibition of NF-κB activation.
Apoptosis is a decisive process used as an intrinsic pro- tection mechanism against cancer initiation [6, 16, 21].
Apoptosis can be prompted by intrinsic or extrinsic path- ways via stimulation of several targets, such as membrane receptors of TNF-α and transforming growth factor-β, and also mitochondria [6]. Confrontation to apoptosis is the ma- jor cause for chemoresistance of most of the cancers, and also one of the crucial targets for improving cancer therapy is the regulation of apoptosis signaling pathways [8, 22].
Indeed, deregulation of apoptosis contributes to the patho- genesis of colon cancer and resistance to chemotherapeutic drugs and radiotherapy, which act at least in part, by killing cancer cells [26]. Previous studies using human breast and gastric cancer cells suggested that anticancer effects of ziyu- glycoside II were mediated apoptosis via mitochondrial dys- function and activation of the caspase-3 pathway [29, 30].
As in these previous studies, we observed an increased amount of cleaved capspase-3 and amplified number of apoptotic cells in TUNEL staining of colonic tissue (Fig. 5).
These observations authorize that the anticancer effects of ziyuglycoside II in colitis-associated colon cancer are parti- ally facilitated by the induction of apoptosis.
Taken all together, our study demonstrated that ziyugly- coside II administration ameliorated colitis-associated tu- morigenesis by inhibiting inflammation and inducing apoptosis. Our data might offer evidence for chemopre- vention of colon cancer, since it has suggested that ziyugly- coside II could be a promising option for colon cancer pre- vention and treatment.
Acknowledgement
This work was supported by research grants from Daegu
Catholic University in 2019.
References
1. Arnold, M., Sierra, M. S., Laversanne, M., Soerjomataram, I., Jema,l A. and Bray, F. 2017. Global patterns and trends in colorectal cancer incidence and mortality. Gut 66, 683-691.
2. Bernstein, C. N., Blanchard, J. F., Kliewer, E. and Wajda, A. 2001. Cancer risk in patients with inflammatory bowel disease: A population-based study. Cancer 91, 854-862.
3. Chen, J., Pitmon, E. and Wang, K. 2017. Microbiome, in- flammation and colorectal cancer. Semin. Immunol. 32, 43-53.
4. Das, V., Kalita, J. and Pal, M. 2016. Predictive and prognostic biomarkers in colorectal cancer: a systematic review of re- cent advances and challenges. Biomed. Pharmacother. 87, 8-19.
5. Eaden, J. A., Abrams, K. R. and Mayberry, J. F. 2001. The risk of colorectal cancer in ulcerative colitis: A meta-analysis.
Gut 48, 526-535.
6. Elmore, S. Apoptosis: A review of programmed cell death.
2007. Toxicol. Pathol. 35, 495-516.
7. Francescone, R., Hou, V. and Grivennikov, S. I. 2015. Cyto- kines, IBD, and colitis-associated cancer. Inflamm. Bowel. Dis.
21, 409-418.
8. Fujita, K., Iwama, H., Oura, K., Tadokoro, T., Samukawa, E., Sakamoto, T., Nomura, T., Tani, J., Yoneyama, H., Morishita, A., Himoto, T., Hirashima, M. and Masaki, T.
2017. Cancer therapy due to apoptosis: Galectin-9. Int. J.
Mol. Sci. 18, pii: E74.
9. Grivennikov, S. I. 2013. Inflammation and colorectal cancer:
colitis-associated neoplasia. Semin. Immunopathol. 35, 229-244.
10. Jang, E., Inn, K. S., Jang, Y. P., Lee, K. T. and Lee, J. H.
2018. Phytotherapeutic activities of Sanguisorba officinalis and its chemical constituents: A review. Am. J. Chin. Med. 46, 299-318.
11. Keller, D. S., Windsor, A., Cohen, R. and Chand, M. 2019.
Colorectal cancer in inflammatory bowel disease: review of the evidence. Tech. Coloproctol. 23, 3-13.
12. Kim, H. J., Park, J. H. and Kim, J. K. 2014. Cucurbitacin-I, a natural cell-permeable triterpenoid isolated from Cucurbi- taceae, exerts potent anticancer effect in colon cancer. Chem.
Biol. Interact. 219, 1-8.
13. Kim, J. K., Shin, E. K., Park, J. H., Kim, Y. H. and Park, J. H. 2010. Antitumor and antimetastatic effects of li- cochalcone A in mouse models. J. Mol. Med. 88, 829-838.
14. Kim, Y. H., Kwon, H. S., Kim, D. H., Shin, E. K., Kang, Y. H., Park, J. H., Shin, H. K. and Kim, J. K. 2009. 3,3'-diindo- lylmethane attenuates colonic inflammation and tumori- genesis in mice. Inflamm. Bowel. Dis. 15, 1164-1173.
15. Kundu, J. K. and Surh, Y. J. 2008. Inflammation: Gearing the journey to cancer. Mutat. Res. 659, 15-30.
16. Liu, Y. and Zeng, G. 2012. Cancer and innate immune sys- tem interactions: Translational potentials for cancer im- munotherapy. J. Immunother. 35, 299-308.
17. Liu, X., Cui, Y., Yu, Q. and Yu, B. 2005. Triterpenoids from Sanguisorba officinalis. Phytochemistry 66, 1671-1679.
18. Marusawa, H. and Jenkins, B. J. 2014. Inflammation and gas-
초록:AOM/DSS로 유도된 마우스 대장암 모델에서의 Ziyuglycoside-II의 항염증효과
천혜진․김진경*
(대구가톨릭대학교 의생명화학부 의생명과학전공)
Ziyuglycoside-II (3β-3-α-1-arabinopyranosyloxy-19-hydroxyurs-12-en-28-oicacid)는 오이풀(Sanguisorba officina- lis L.)의 주요 활성 화합물 중 하나로 지혈작용, 항산화활성, 항염증활성 등의 활성이 보고되어 있다. 그러나 염증 성 대장암에서의 ziyuglycoside-II의 활성에 관해서는 아직 보고되지 않아 본 연구에서 azoxymethane (AOM)/
dextran sulfate sodium (DSS)으로 유발된 대장암 모델에서 ziyuglycoside-II 항종양활성을 조사하였다. 염증성 대 장암을 유발하기 위해 BALB/c 마우스에게 AOM을 1회 복강 주사하고 AOM 투여 1주일 후 총 3 cycle의 2% 농도 의 DSS를 음용수로 공급 하였다. Ziyuglycoside-II (1 또는 5 mg/kg)는 1주일에 3회 경구 투여하고, 64일에 대장을 적출하였다. 대장 조직에서의 종양 개수를 관찰한 결과 ziyuglycoside-II의 투여가 종양의 형성을 유의적으로 감소 시키는 것을 확인하였다. 또한 Western blot 방법과 면역 조직학 분석의 결과, ziyuglycoside-II의 투여가 대장 조 직에서 nuclear factor kappa-B 양성 세포와 염증 관련 단백질의 양을 현저히 감소시킴을 관찰하였다. 또한 ziyu- glycoside-II 투여에 의해 대장조직내의 세포사멸이 촉진되었고 cleaved-caspase 3, cleaved-caspase 7과 같은 세포 사멸 관련 단백질의 발현이 증가함을 확인하였다. 이러한 결과는 ziyuglycoside-II의 투여가 염증반응을 억제하고 세포 자멸을 유도하여 염증성대장암의 발생을 억제함을 시사하고 있다.
trointestinal cancer: an overview. Cancer Lett. 345, 153-156.
19. Mitchell, J. P. and Carmody, R. J. 2018. NF-κB and the Transcriptional Control of Inflammation. Int. Rev. Cell Mol.
Biol. 335, 41-84.
20. Nam, S. H., Lkhagvasuren, K., Seo, H. W. and Kim, J. K.
2017. Antiangiogenic effects of ziyuglycoside II, a major ac- tive compound of Sanguisorba officinalis L. Phytother. Res. 31, 1449-1456.
21. Nomi, N., Kodama, S. and Suzuki, M. 2010. Toll-like re- ceptor 3 signaling induces apoptosis in human head and neck cancer via survivin associated pathway. Oncol. Rep. 24, 225-231.
22. Owen, H. C., Appiah, S., Hasan, N., Ghali, L., Elayat, G. and Bell, C. 2017. Phytochemical modulation of apoptosis and autophagy: strategies to overcome chemoresistance in leuke- mic stem cells in the bone marrow microenvironment. Int.
Rev. Neurobiol. 135, 249-278.
23. Park, J. H. and Kim, J. K. 2018. Pristimerin, a naturally oc- curring triterpenoid, attenuates tumorigenesis in experi- mental colitis-associated colon cancer. Phytomedicine 42, 164- 171.
24. Priyadarsini, R. V. and Nagini, S. 2012. Cancer chemo- prevention by dietary phytochemicals: promises and pitfalls.
Curr. Pharm. Biotechnol. 13, 125-136.
25. Siegel, R. L., Miller, K. D., Fedewa, S. A., Ahnen, D. J., Meester, R. G. S., Barzi, A. and Jemal, A. Colorectal cancer statistics, 2017. 2017. CA. Cancer J. Clin. 67, 177-193.
26. Watson, A. J. An overview of apoptosis and the prevention of colorectal cancer. 2006. Crit. Rev. Oncol. Hematol. 57, 107- 121.
27. Zeligs, K. P., Neuman, M. K. and Annunziata, C. M. 2016.
Molecular Pathways: The balance between cancer and the immune system challenges the therapeutic specificity of tar- geting nuclear factor-κB signaling for cancer treatment. Clin.
Cancer Res. 22, 4302-4308.
28. Zhao, Z., He, X., Zhang, Q., Wei, X., Huang, L., Fang, J. C., Wang, X., Zhao, M., Bai, Y. and Zheng, X. 2017. Traditional uses, chemical constituents and biological activities of plants from the Genus Sanguisorba L. Am. J. Chin. Med. 45, 199-224.
29. Zhu, A. K., Zhou, H., Xia, J. Z., Jin, H. C., Wang, K., Yan, J., Zuo, J. B., Zhu, X. and Shan, T. 2013. Ziyuglycoside II-in- duced apoptosis in human gastric carcinoma BGC-823 cells by regulating Bax/Bcl-2 expression and activating caspase-3 pathway. Braz. J. Med. Biol. Res. 46, 670-675.
30. Zhu, X., Wang, K., Zhang, K., Huang, B., Zhang, J., Zhang, Y., Zhu, L., Zhou, B. and Zhou, F. 2013. Ziyuglycoside II inhibits the growth of human breast carcinoma MDA-MB- 435 cells via cell cycle arrest and induction of apoptosis through the mitochondria dependent pathway. Int. J. Mol.
Sci. 14, 18041-18055.
31. Zhu, X., Wang, K., Zhang, K., Zhu, L. and Zhou, F. 2014.
Ziyuglycoside II induces cell cycle arrest and apoptosis through activation of ROS/JNK pathway in human breast cancer cells. Toxicol. Lett. 227, 65-73.