Buddleja officinalis prevents the normal cells from oxidative damage via antioxidant activity
8
0
0
전체 글
(2)
(3)
(4)
(5)
(6)
(7)
(8)
수치
관련 문서
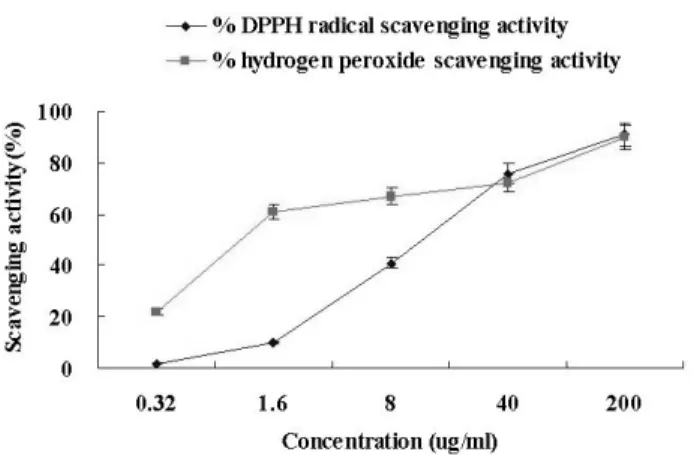
To measure antioxidant effects of the yuzu seed oil, DPPH radical scavenging and lipid peroxidation were investigated... DPPH radical scavenging
The characteristics of typhoons were examined via synoptic analysis and track examination, and the damage types and scales were analyzed to contribute to
In chlorination, the transition state resembles the reactant (small radical character), therefore the tertiary radical is slightly more stable than the primary one. In
Fresh samples were immediately tested for antioxidant activity, and antioxidant activity was compared according to gestational age and measured the difference
DPPH radical scavenging activity of Yanggaeng added with different levels of Molokhia( Corchorus olitorius L.) powder.. Bars with different letters
According to the result of comparative study on nutrient components and antioxidant activity in Abelmoschus esculentus depending on drying methods amino
Antiinflammatory and superoxide radical scavenging activities of procyanidins containing extract from the bark of Pinus pinaster Sol.. and
Effect of hydrogen peroxide on photooxidation rate of aquouse Phenol and Chlorophenols solution in continuous flow system... Total ion chromatogram by
