대한소화기학회지 2007;49:85-92
접수: 2006년 3월 6일, 승인: 2007년 1월 4일
연락처: 설상영, 614-735, 부산시 부산진구 개금2동 633-165 인제대학교 의과대학 내과학교실
Tel: (051) 890-6158, Fax: (051) 890-0273 E-mail: [email protected]
Correspondence to: Sang Yong Seol, M.D.
Department of Internal Medicine, Inje University College of Medicine, 633-165, Gaegeum 2-dong, Busanjin-gu, Busan 614- 735, Korea
Tel: + 82-51-890-6158, Fax: + 82-51-890-0273 E-mail: [email protected]
B 형 간염바이러스 중합효소의 역전사효소 활성을 억제하는 인간 단세포군 항체
인제대학교 의과대학 내과학교실, 화명한솔병원*, 인제대학교 의과대학 미생물학교실†
박성재·설상영·지삼룡·박은택·이연재·이상혁·정정명·조현대*·정영주
†·최인학
†·박세광
†H u m a n M o n o c l o n a l A n t i b o d y I n h i b i t i n g R e v e r s e T r a n s c r i p t a s e A c t i v i t y o f H e p a t i t i s B V i r u s P o l y m e r a s e P r o t e i n
S u n g J a e P a r k , M . D. , S a n g Y o n g S e o l , M . D. , S a m R y o n g J e e , M . D. , E u n T a i k P a r k , M . D. , Y o u n J a e L e e , M . D. , S a n g H y u k L e e , M . D. , J u n g M y u n g Ch u n g , M . D. ,
H y u n D a e Ch o , M . D. *, Y o u n g -J u J e o n g†, I n H a k C h o i , M . D.†, a n d S a e G w a n g P a r k , M . D.†
Departments of Internal Medicine and Microbiology†, Inje University College of Medicine, Busan;
Hwamyung Hansol Hospital*, Busan, Korea
Background/Aims: To develop a novel treatment method for hepatitis B virus (HBV) infection, we aimed to make a human monoclonal antibody inhibiting reverse transcriptase (RT) activity of P protein which was impor- tant in HBV replication by using phage display technique. Therefore, we analysed the usability of human monoclonal antibody as a protein based gene therapy. Methods: Reverse transcriptase/polymerase (RT/POL) functional motif of P protein of HBV was cloned in pMAL-c vector and expressed as maltose binding fusion protein form. The RT/POL recombinant protein (pMRT/POL) was purified by amylose resin column. Using human single chain Fv phage antibody library with 1.1×1010 size, human antibody against pMRT/POL was selected with BIAcore panning. Selected antibody fragments were analyzed for the activity of RT inhibition. Finally, they were analyzed for the affinity with BIAcore and the complementarity determining regions with nucleotide sequencing.
Results: pMRT/POL recombinant protein expressed in E. coli showed RT activity, 1μg of recombinant protein had an activity equivalent to 5 unit of MMLV RT. By BIAcore panning, we could select 3 clones; POL-A5, POL-B8 and POL-B12. Each clone's RT inhibiting activity were 52-82%, affinity against antigen were 8.15×10-8
M to 1.75×10
-6 M. Conclusions: Human monoclonal antibodies produced in this study showed low affinity, but efficiently inhibited the activity of RT in vitro. If POL-A5, POL-B8, and POL-B12 can be converted to intracellular antibody form, it can be used for protein-based gene therapy by inhibiting the replication through the neutralization of polymerase protein of HBV. (Korean J Gastroenterol 2007;49:85-92)
Key Words: Hepatitis B virus; Reverse transcriptase; Phage display; Human monoclonal antibody
86 대한소화기학회지: 제49권 제2호, 2007
서 론
B형 간염바이러스(hepatitis B virus, HBV)는 hepadnaviri- dae에 속하는 외피를 보유한 DNA 바이러스이다. 만성 보유 자가 전 세계적으로 3억 이상인 것으로 추정한다.1 전체 환 자의 5%가 만성 활동 간염을 앓는데, 이는 간경변과 간암의 주 원인 중 하나이다.2 만성 간질환은 우리나라에서 아직도 연간 인구 10만 명당 41.7명이 만성 간질환(19.6명)이나 간 세포암종(22.6명)으로 사망하는 중요한 사회 질환으로,3 만 성 간질환의 70-80%가 만성 B형 간염과 관계있다.4
최근에 만성 B형 간염에 인터페론, 페그인터페론, 항바이 러스 제제로서 라미부딘과 아데포비어, 엔테카비어 등을 사 용하여 치료 대상과 치료제 선택의 폭이 넓어졌으나 작용 기전과 치료효과에 몇 가지 한계점을 가지고 있다.5-9 이런 문제점을 극복하기 위하여 여러 가지 시도가 이루어지고 있 으며, antisense oligonucleotide-base gene therapy도 그 한 예
이다.10,11 이 방법으로 일정 범위 내에서 바이러스 복제를
억제할 수 있지만, 항바이러스 효과가 바이러스의 염기서열 의 변이에 민감하고 치료에 저항하는 새로운 바이러스들이 보고되고 있다.12
유전자 치료의 또 다른 접근 방식으로, 세포 내에서 생산 된 펩타이드나 단백질의 기능을 억제하는 단백질에 기초한 (protein-based) 방법이 있다. 이 경우 바이러스의 염기서열 변이에 영향을 상대적으로 덜 받기 때문에 oligonucleotide- base 방식에 비하여 보다 강력한 방법이다.13Single-chain 항 체(scFv)는 항체의 중쇄와 경쇄가 linker로 연결된 man-made 분자로서 항체의 친화도나 특이도를 그대로 유지하고 있
다.14-16 항원 결합에 필요한 중쇄와 경쇄의 두 가변 영역을
하나의 상보 DNA (complementary DNA, cDNA)에서 coding 함으로 scFV는 세포 내에서 하나의 cDNA로 발현할 수 있 다. 그래서 scFv는 protein-based 유전자 치료에 아주 적합한 물질로 생각하고 있다.
HBV 복제에서 가장 중심적인 역할을 하는 것은 중합효 소단백(P 단백질)이다. P 단백은 terminal protein (TP), spa- cer, reverse transcriptase/polymerase (RT/POL) 그리고 RNase H의 4개 기능 분절로 구성되어 있다.2 TP 분절은 RNA intermediates의ε(encapsidation signal) 부위를 인지하는 기능 을, RT/POL 분절은 RNA intermediates로부터 DNA를 만드는 역전사효소(RNA dependent DNA polymerase) 기능과 역전사 로 만들어진 DNA로부터 DNA를 복제하는 중합효소(DNA dependent DNA polymerase) 기능을, RNase H 분절은 RNA- DNA hybrid를 인지하여 RNA를 분해하는 기능을 각각 가지 고 있다. P 단백은 3.5 kb의 RNA intermediates로부터 낮은 효율로 번역되어 만들어져 소수만이 세포 내에 존재하며, cis-acting으로ε부위를 인지하고 역전사 효소기능을 이용하
여 minus strand DNA를 생성하여 복제를 시작한다. Radzi- will 등17은 역전사효소 기능부위를 돌연변이시키면 viral DNA가 합성되지 않아 복제가 되지 않는다고 하였는데 이 는 scFv로 역전사효소 기능부위의 활성을 억제하였을 때도 같은 결과가 나올 것을 예상할 수 있다.
이번 연구에서는 대장균에서 재조합으로 발현 정제된 P 단백의 RT/POL 기능부위에 대한 인간 단세포군 항체를 phage display 기법으로 제작하고, 제작된 인간 단세포군 항체 의 생체외적인 역전사효소 활성 억제능을 분석하여 pro- tein-based 유전자 치료로서 활용 가능성을 검색하고자 한다.
대상 및 방법
1. P 단백질의 대장균 내 발현과 정제
HBV P 단백의 RT/POL 기능부위 유전자는 HBV adw의 전 체 유전자가 들어 있는 pAM6 플라스미드(ATCC 45020)에서 중합효소연쇄반응(polymerase chain reaction, PCR)으로 획득하 였으며, 대장균 내 발현은 maltose binding protein (MBP)과 융 합단백질로 발현되게 하는 pMAL-c (NEB, USA)를 사용하였 다. PCR은 POL5 (5'-gag ctg cag ctc gtg gtg gac ttc tct-3')와 POL3 (5'-caa aag ctt ggc aca gac cag gcc gtt-3')의 프라이머 조합을 사용하였으며, 증폭된 유전자는 Qiaex II gel extraction kit (Qiagen, Germany)를 사용하여 agarose gel로부터 추출하였 다. 정제된 PCR 산물은 제한효소인 PstI과 HindIII로 절단하 고 다시 Qiaex II gel extraction kit로 정제한 후 동일한 효소로 절단하여 정제된 pMAL-c 벡터에 클로닝하였다.
클로닝된 플라스미드를 E. coli XL1-Blue (NEB, USA)에 형질 전환시킨 후 37oC에서 18시간 배양하였다. 배양된 대 장균을 100배 희석하여 600 nm 파장에서 흡광도가 0.5가 되 도록 계대 배양한 후 isopropyl thio-β-D-galactoside (IPTG)를 최종 농도 1 mM가 되도록 첨가한 후 6시간 동안 더 진탕배 양하여 재조합 단백질이 과발현되도록 하였다. 재조합 단백 질의 과발현 유무를 8% sodium dodecyl sulfate-polyacryla- mide gel electrophoresis (SDS-PAGE)상에서 전기영동하고 PVDF membrane에 이적한 후 항 MBP 항체를 사용하여 we- stern blot으로 확인하였다.
과발현이 확인된 클론들은 0.3 mM IPTG가 포함된 배지 로 22oC에서 48시간 동안 대량으로 배양하였다. 세균배양액 을 4oC에서 20분간 4,000 g로 원침한 후 0.05% Tween 20이 첨가된 아밀로즈 레진칼럼 완충액(20 mM Tris-HCl pH 7.4, 200 mM NaCl, 1 mM EDTA, 1 mM azide)에 부유시키고 액 체질소를 사용하여 3회 동결 및 해동을 반복한 다음 초음파 분쇄기로 파쇄하였다. 그런 다음 8,000 g로 원침하여 얻어진 상청액을 아밀로즈 레진칼럼(NEB, USA)에 분당 2 mL의 유
박성재 외 10인. B형 간염바이러스 중합효소의 역전사효소 활성을 억제하는 인간 단세포군 항체 87
속으로 흘려 재조합 단백질을 아밀로즈 레진에 결합시키고 10 mM maltose 용액으로 용출하였다. 단백질 농도는 Brad- ford 방법(Bio-rad laboratories Inc., Hercules, CA)으로 정량하 고, SDS-PAGE 후 Coomasie-blue 염색과 western blot으로 순 도를 확인하였다.
재조합 단백질의 역전사효소 활성은 Jeong 등18의 방법을 일부 변형하여 실시하였다. 정제된 재조합 pMRT/POL 단백 질을 poly (rA)·oligo (dT)12-18 homopolymer 0.1μg과 혼합한 후 RNA-dependent DNA 중합효소 반응용액(50 mM Tris-HCl pH 7.4, 50 mM KCl, 0.5 mM MnCl, 1 mM DTT, 0.25 mM dTTP, 5μCi32P dTTP)을 첨가하여 최종 20μL가 되게 한 후 37oC에서 1시간 30분 동안 반응하였다. 반응의 정지는 proteinase K (0.2 mg/ mL in 0.5% SDS)를 첨가하여 37oC에 서 20분간 추가로 반응하여 실시하였다. 역전사효소 활성 유무는 반응액을 6% polyacrylamide gel에서 전기영동한 후 autoradiograph으로 확인하였으며, 정량적인 분석은 반응액 을 Whatman DE81 filter paper에 점적하고 0.5 M Na2HPO4로 4번, 3차 증류수로 2번 세척한 후 Wallac TriLux Beta liquid scintillation counter (Wallac Inc., Fin)로 radioactivity를 측정하 여 실시하였다. 다른 역전사효소와의 활성정도 비교는 MMLV-RT (Promega, USA)를 사용하였다. 모든 효소 활성 측정은 3회 이상 반복 실험을 실시하였다.
2. pMRT/POL에 대한 인간 단세포군 항체의 선택
pMRT/POL에 대한 인간항체 제작에는 연구자들이 기존에 제작한 1×1010cfu의 사람 scFv 항체 분절 library를 사용하 였다.19 항체 library에서 pMRT/POL 단백을 인지하는 항체 클론의 선택은 BIAcore 2000 (Biacore AB, Uppsala, Sweden) 을 사용하였다. pMRT/POL 단백의 고정에는 CM5 chip과 amine coupling 방법을 사용하였다. pMRT/POL 단백이 고정 된 flow cell에 항체 library phabs를 2μL/min으로 흘려 결합 시킨 후 해리되는 시간별로 6시간 동안 회수하였다. 6시간 후 regeneration buffer (25 mM NaOH, 0.5 M NaCl)를 사용하 여 해리되지 않은 phage를 회수하였다.
회수된 phage들을 대수 증식기의 ER2537에 감염시킨 후 50μg/mL의 ampicillin이 첨가된 LB 고형 배지에서 배양하여 집락을 형성하게 하였다. 96 well 세균배양 plate에 형성된 각 집락을 SB 배지로 배양하고 VCSM13을 감염시켜 phage 를 제작하였다. 제작된 각각의 phage를 pMRT/POL 단백이 고정된 flow cell에 다시 흘려 각 클론의 pMRT/POL 단백에 대한 결합 강도를 확인하여 강하게 결합하는 클론들을 선택 하였다. 선택된 phage 클론을 대장균 Top10F' (Invitrogen, CA)에 감염시키고 37oC SB 배지에서 배양하고 600 nm에서 흡광도가 0.9-1.0일 때 IPTG를 넣고 8시간 더 배양함으로써 항체 분절의 수용 발현을 유도하였다. 대장균을 수확하여
lysis buffer (10 mM Na2HPO4 pH 8.0, 500 mM NaCl, 10 mM imidazole)에 부유한 후 초음파로 세포를 분쇄하였다. 원심 분리한 상청액에서 항체분절은 Ni-NTA column (Qiagen, Valencia)과 imidazole buffer를 이용하여 정제하여 사용하였 다.
3. 항체 친화도 측정
BIAcore 2000을 사용하여 정제한 항체 분절의 pMRT/POL 에 대한 친화도를 측정하였으며 이때 CM5 chip에 고정한 RU 값은 4,000이었다. 정제된 항체 분절을 농도별로 20μL/
min 속도로 흘려 결합과 해리를 측정하고 Biaevaluation soft- ware (Biacore AB, Uppsala, Sweden)로 kon과 koff를 측정하고 KD 값을 계산하였다.
4. 선택된 항체의 역전사효소 활성 억제
정제된 pMRT/POL 단백 1μg과 항체 1μg을 실온에서 1 시간 반응하고 pol (rA)·olig (dT)12-18 homopolymer 100 ng과 RDDP 반응 혼합액(50 mM Tris-HCl pH 7.4, 50 mM KCl, 0.5 mM MnCl, 1 mM DTT, 0.25 mM dTTP, 50μCi 32P dTTP)을 첨가하여 최종 20μL되게 하여 37oC에서 1시간 30분 동안 반응하였다. 반응액을 DE81 chromatography paper에 적시고 0.5 M Na2HPO4로 4회 세척하고 증류수로 2회, 마지막으로 95% ethanol로 세척하였다. 반응액의 radioactivity는 liquid scintilation counter (Wallac Inc, Fin)를 이용하여 인간 단세포 군 항체의 역전사효소 활성 억제 정도를 측정하였다.
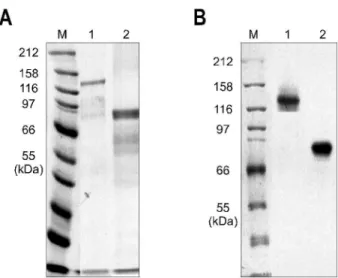
Fig. 1. Purification of recombinant HBV polymerase in E. coli.
pMTPR and pMRT/POL are purified with amylose resin column chromatography. (A) Coomassie blue stain, (B) Western blot.
Lane M, bio-rad broad range protein marker; Lane 1, pMTPR;
Lane 2, pMRT/POL.
88 The Korean Journal of Gastroenterology: Vol. 49, No. 2. 2007
5. 염기서열 분석과 CDR 확인
ABI prism 3100을 사용하여 각 항체 클론의 염기서열을 분석하였으며 VH 유전자 분석을 위하여 중쇄 유전자의 경 우 HRML-F (5' ggt ggt tcc tct aga tct tcc 3')를, 경쇄 유전자 의 경우 ompseq (5' aag aca gct atc gcg att gca g 3')를 primer 로 각각 사용하였다. 염기서열 분석 결과는 Genetyx를 사용 하여 아미노산 서열을 유추하였으며 중쇄와 경쇄의 항원 결 정 부위를 확인하였다.
결 과
1. P 단백질의 대장균 내 발현과 정제
PCR 클로닝으로 제작된 pMRT/POL 재조합 단백은 예상 하였던 크기인 82 kDa와 같았으며 anti-MBP antibody와도 잘 반응하였다(Fig. 1).
Poly (rA)·oligo (dT)12-18 homopolymer를 이용한 역전사효 소 활성 확인을 통하여 정제된 MBP 단백은 역전사효소의 활 성이 없었으나 pMRT/POL 재조합 단백은 효소 활성을 보였
Fig. 3. Purification and analysis of binding kinetics of soluble scFvs. (A) Antibody fragments are expressed in the form of soluble proteins without p3 phage protein fused to a C-terminal of scFv. Monomeric scFv are purified with size exclusion column and subjected to 12% SDS-PAGE. (B) Binding kinetics of scFvs are measured by surface plasmon resonance on a BIAcore biosensor instrument.
Overlay plot of sensorgrams obtained for POL-A5 at five different concentrations (1-100μg/ml) against immobilized pMRT/POL are seen.
The data represents three independent experiments.
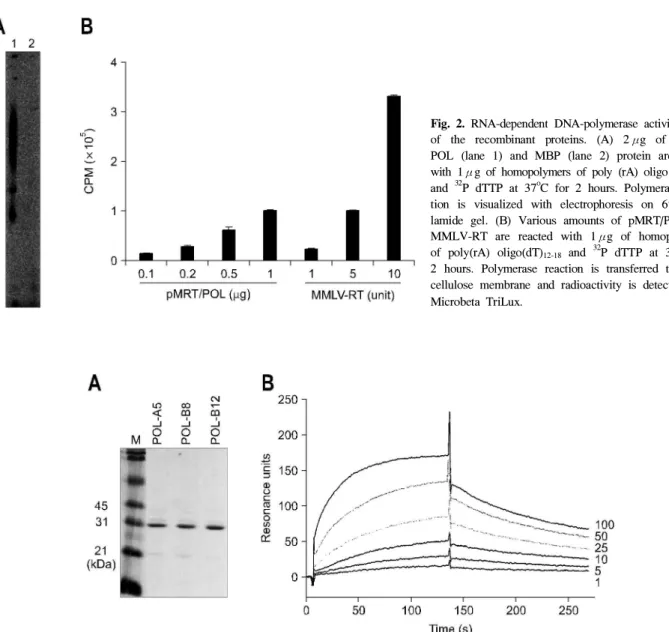
Fig. 2. RNA-dependent DNA-polymerase activity assay of the recombinant proteins. (A) 2μg of pMRT/
POL (lane 1) and MBP (lane 2) protein are mixed with 1μg of homopolymers of poly (rA) oligo (dT)12-18
and 32P dTTP at 37oC for 2 hours. Polymerase reac- tion is visualized with electrophoresis on 6% acry- lamide gel. (B) Various amounts of pMRT/POL and MMLV-RT are reacted with 1μg of homopolymers of poly(rA) oligo(dT)12-18 and 32P dTTP at 37oC for 2 hours. Polymerase reaction is transferred to DE81 cellulose membrane and radioactivity is detected with Microbeta TriLux.
Park SJ, et al. Human Monoclonal Antibody Neutralizing HBV Reverse Transcriptase 89
다(Fig. 2). MMLV-RT와 비교하였을 때 pMRT/POL 1μg은 5 unit의 MMLV-RT와 같은 정도의 활성을 보였다(Fig. 2). 대 장균에서 재조합으로 발현, 정제한 pMRT/POL 단백이 역전 사효소 활성을 유지하고 있어 이것을 항원으로 하여 인간 단세포군 항체를 제작하였다.
2. pMRT/POL에 대한 인간 단세포군 항체의 선택
BIAcore 2000의 recover 기능을 사용하여 항체 분절 library 로부터 pMRT/POL을 인지하는 POL-A5, POL-B8 그리고 POL-B12의 3개 클론을 선택하였다. 각 클론의 항체분절 발현 을 확인하기 위하여 대장균 Top10F'에서 IPTG로 발현을 유 도한 결과 34 kDa의 anti-His 항체와 강하게 반응하는 특이 band를 SDS-PAGE와 western blot에서 확인하였다(Fig. 3A).
3. 항체 친화도 측정
BIAcore로 선택된 항체 분절의 pMRT/POL 단백에 대한 친화도를 측정하였다. POL-A5은 kon 값이 3.81×103 1/Ms, koff값이 3.11×10-41/s로 KD는 8.15×10-8M이었으며 POL- B8은 kon값이 8.94×102 1/Ms, koff값이 3.61×10-41/s로 KD
는 4.04×10-7 M, POL-B12는 kon 값이 1.38×103 1/Ms, koff
값이 2.41×10-3 1/s로 KD는 1.75×10-6 M이었다(Fig. 3B).
4. 항체 분절의 역전사효소 억제능 분석
선택된 항체분절을 Ni-NTA column을 사용하여 정제한 후 pMRT/POL 단백질 5μg과 실온에서 10분간 반응한 후 역전 사효소 반응을 실시한 결과 POL-A5, POL-B8 그리고 POL- B12의 역전사효소 억제능은 각각 82%, 73% 그리고 52%이 었으며, 음성대조군으로 사용된 RN3020에서는 역전사효소 의 억제가 관찰되지 않았다(Fig. 4).
5. 염기서열 분석과 CDR 확인
HRML-F와 ompseq primer를 사용하여 항체분절 유전자의 염기서열을 분석한 결과 경쇄는 POL-A5이 327 nt, POL-B8 이 333 nt, 그리고 POL-B12가 324 nt로 구성되어 있었으며, 중쇄는 408 nt, 405 nt, 그리고 396 nt로 각각 구성되어 있었 다. 분석된 염기서열을 kabat database (http://immuno.bme.
nwu.edu/)를 사용하여 항체분절의 항원 결정 부위들을 확인 하였다(Table 1).
Table 1. Amino Acid Sequences of Anti-pMRT/POL Antibody Fragments VL
CDR1 CDR2 CDR3
POL-A5 RASHFIGGTTYVA GASARAS HQYGNSPTT
POL-B8 SGSSSNIGSNFVT DNDKRPS GTWDSSLSVRV
POL-B12 RASQSVDRYLA DTSTRAT QQRKFWPPIT
VH
CDR1 CDR2 CDR3
POL-A5 GFSFENYAMH AINWNSDTIGYADSVKG DRGPTARDMDA
POL-B8 GFTFSSYGMH VIWYDGSNKYYADSVKG GGPYYYYYMDV
POL-B12 GFTFSTYSMN YISSRYGNIYYSDSVKG DWSYSFDY
Fig. 4. Inhibition assay of reverse transcritase activity of pMRT/
POL. One microgram of pMRT/POL is reacted with 1μg of each anti- RT/POL antibody fragments or RN30 (anti-RNase H) at room temperature for 1 hour. After adding RDDP reaction mixture (50 mM Tris-HCl pH 7.4, 50 mM KCl, 0.5 mM MnCl, 1 mM DTT, 0.25 mM dTTP, 50μCi 32P dTTP), they are incubated at 37oC for 90 minutes. Radioactivity are detected with MicroBeta TriLux. The reverse transcriptase activity of pMRT/
POL only is calculated as 100%. The data represents three independent experiments.
90 대한소화기학회지: 제49권 제2호, 2007
고 찰
이번 연구는 HBV P 단백의 RT/POL 기능 부위의 기능을 억제하는 인간 단세포군 항체를 phage display 기법을 이용 하여 제작하여 protein-based gene therapy의 가능성을 타진하 고자 하였다. HBV의 복제에 있어 필수적인 과정은 prege- nomic RNA의 역전사와 second-strand DNA의 합성이며,2,21 이 과정을 수행하는 것이 바로 P 단백이다. Pregenomic RNA로부터 만들어진 P 단백은 cis-acting으로 작용하며, TP, spacer, RT/POL 그리고 RNAse H의 4개 기능 부위를 가지고 있다.2 기존 연구에서 제작된 재조합 P 단백은 94 kDa 크기 의 전체 단백질을 발현하였다.18,22 전체 P 단백을 대장균에 서 재조합으로 발현하는 경우 크기 때문에 발현율이 낮다.
이번 연구에서는 전체 P 단백 중 RT/POL 기능 부위만을 MBP 융합단백의 형태로 발현하여 재조합 단백의 발현율을 높였다.
이번 연구에서는 phage display 기법을 이용하여 항체를 제작하였는데, 실험동물에서 제작되는 일반적인 단세포군 항체는 진단이나 분석과 같이 생체외적으로 사용하는 데 아 무런 문제가 없지만, 치료나 진단의 목적으로 인체 내 투여 하면 이종단백으로 인지되는 human anti-mouse antibody 반 응을 유발하여 반감기가 감소되고 재차 사용시 과민 반응을 일으킬 수 있다. 이러한 단점을 극복하기 위한 방법으로 인 간 B 림프구를 in vitro immunization한 후 하이브리도마 제 작, 항체의 불변영역을 인간 유전자로 치환된 transgenic mouse 이용, 마우스 단세포군 항체의 humanization 그리고 phage display 기법을 이용한 human antibody library의 제작 등을 시도하고 있다.23-26 In vitro immunization은 하이브리도 마의 제작 시에 사용하는 Epstain-Barr virus의 B 세포 불멸 화의 성공률이 1-3%로 낮으며, transgenic mouse를 이용하는 경우 면역이나 하이브리도마를 쉽게 만들 수 있으나 trans- genic mouse를 가지고 있어야 하며, 마우스 단세포군 항체의 humanization은 각각의 항체에 따라 다른 방식이 요구된다는 단점이 있다. Phage display 기법은 항체에서 항원을 인지하 는 분절만을 filamentous phage의 표면에 발현하는 기법으로 phage의 life cycle을 이용하여 in vitro panning이 가능하다.
In vitro panning이 가능함으로써 면역을 거치지 않고도 단세 포군 항체를 제작할 수 있어 면역되지 않은 항체 library (naive library)나 synthetic library를 활용하고 있다. HBV가 감염된 환자에서 P 단백에 대한 항체가 만들어지지 않기 때 문에 이번 연구에서도 naive antibody library를 사용하였으 며, 연구에 사용된 library는 50인의 정상 성인 말초혈액 림 프구로부터 제작된 것으로 1.1×1010 cfu의 크기이다.
항체 library에서 특정 항원에 결합하는 클론을 선택하기 위하여 다양한 방법이 사용된다. 가장 많이 사용되는 방법
은 고전적인 panning이다.27 이외에도 columns이나 BIAcore sensorchips 같은 고형 보조물에 고정된 항원, biotinylated 항 원, 원핵세포에 고정된 항원, 진핵세포에 고정된 항원, 조직 절편이나 단편 등을 이용한 방법들이 사용된다. Surface plasmon resonance에 기반을 둔 BIAcore는 생물특이적인 상 호작용의 실시간 분석이 가능하여 항원-항체반응의 역동 학적인 측면을 이용한 항체 선택을 가능하게 한다. 이번 연구에서도 BIAcore를 항체 선택에 사용하였으며 koff 값이 높은 클론을 선택하기 위하여 6시간의 해리시간을 사용하 여 10-6-10-8M 정도의 친화도를 가진 POL-A5, POL-B8 그 리고 POL-B12의 세 개 클론을 찾아냈다. POL-A5 클론만이 10-8M의 치료용 항체의 일반적인 친화도를 보였지만 naive antibody library를 사용하여 제작된 항체의 친화도가 library 크기에 비하여 102 정도 낮다는 점을 고려할 때 적절하다.
제작된 항체분절들은 52-82%의 역전사효소 활성 억제를 보였다. 이 결과는 Gargano 등28이 synthetic library를 사용하 여 human immunodeficiency virus RT의 역전사 효소 활성을 정제된 항체 분절 100 nM 농도로 95% 이상 억제하는 단세 포군 항체를 제작하였다는 보고에 비하여 낮은 결과다. 그 러나 항체 분절들의 역전사효소 활성 억제능이 항원에 대한 친화도와 비례한다는 점을 고려할 때 낮은 친화도를 가진 항체 분절 항체사슬을 새로운 항체사슬과 서로 치환하는 chain shuffling29,30이나 변이유발 세균을 이용하는 방법31,32 또는 특정 부위의 염기서열에 변이를 유발시키는 site-di- rected mutagenesis33,34등의 방법을 사용하여 10배에서 1,000 배까지 친화도를 증가시킨다면 더욱 높은 역전사효소 활성 억제능을 가진 항체를 제작할 수 있을 것이다.
이번 연구로 제작된 재조합 RT/POL 기능부위 단백은 역 전사효소 활성을 가지고 있었으며, P 단백질의 역전사효소 활성을 억제하는 항체 제작에서 panning과 screening에 적합 하다고 판단된다. RT/POL 재조합 단백은 항체이외의 역전 사효소 활성 억제 물질의 발굴에도 활용될 수 있을 것이다.
Naive library로 HBV의 P 단백의 RT/POL 효소 활성 부위를 중화하는 항체 제작 가능성을 확인하였으며 이를 기반으로 보다 높은 중화 능력을 가진 항체를 제작하고 세포내 항체 로 발현한다면 HBV 감염의 새로운 치료법이 될 수 있을 것 이다.
요 약
목적: 간경변과 간암의 주 원인 중 하나인 B형 간염바이 러스(hepatitis B virus, HBV)에 대한 새로운 치료법의 개발 의 일환으로 phage display 기법을 이용하여 바이러스 복제 에 있어 중요한 P 단백의 역전사효소 활성을 억제하는 인간 단세포군 항체를 제작하여 protein based 유전자 치료 가능
박성재 외 10인. B형 간염바이러스 중합효소의 역전사효소 활성을 억제하는 인간 단세포군 항체 91
성을 분석하고자 하였다. 대상 및 방법: HBV P 단백의 역 전사효소 기능 부위인 RT/POL 기능 부위를 pMAL-C 벡터에 클로닝하고 maltose binding protein과 융합 단백의 형태로 재 조합 단백으로 발현시켰다. 1.1×1010의 인체 single chain Fv phage antibody library를 사용하여 RT/POL 재조합 단백에 대 한 항체를 BIAcore로 선택하였다. 선택된 항체의 역전사효 소, RNA-dependent DNA 중합효소 기능 정도를 분석하였으 며 BIAcore로 항원에 대한 친화도를 그리고 염기서열 분석 을 통하여 항원 결정 부위를 확인하였다. 결과: 대장균에서 발현 정제된 RT/POL 기능 부위 재조합 단백은 역전사효소 활성을 유지하고 있었으며, 1μg의 재조합 단백은 5 unit의 MMLV 역전사효소와 동일한 효소 활성을 가지고 있었다.
BIAcore를 이용한 RT/POL에 대한 항체의 선택으로 POL- A5, POL-B8 그리고 POL-B12의 3개 클론을 확보하였다. 선 택된 클론들의 역전사효소 활성 억제능은 각각 82- 52%였 으며, 항원에 대한 친화도는 8.15×10-8 M to 1.75×10-6M 이었다. 결론: 이번 연구를 통하여 제작된 인간 단세포군 항체는 항원에 대한 친화도가 낮았으나 in vitro에서 효과적 으로 역전사효소 활성을 억제하여 protein based gene therapy 로서 세포 내 항체로 발현한다면 HBV P 단백의 기능을 억 제하여 바이러스의 복제를 억제할 것이다.
색인단어: B형 간염바이러스, 역전사효소, 파지 디스플레 이, 인간 단세포군 항체
참고문헌
1. Lau JY, Wright TL. Molecular virology and pathogenesis of hepatitis B. Lancet 1993;342:1335-1340.
2. Ganem D. Hepadnaviridae and their replication In: Fields BN, Knipe DM, Howley PM, et al, ed. Fields virology. 3rd ed.
Philadelphia: Lippincott-Raven, 1996:1199-1233.
3. 통계청. 2004년 사망원인통계결과. http://www.nso.go.kr.
4. Jung SH, Kim BH, Joung YH, et al. Clinical features of hepatocellular carcinoma in the 1990s. Korean J Gasteroen- terol 2003;42:649-657.
5. Krogsgaard K, Bindslev N, Christensen E, et al. The treat- ment effect of alpha interferon in chronic hepatitis B is inde- pendent of pre-treatment variables. Results based on indivi- dual patient data from 10 clinical controlled trials. European Concerted Action on Viral Hepatitis (Eurohep). J Hepatol 1994;21:646-655.
6. Liaw YF, Leung NW, Chang TT, et al. Effects of extended lamivudine therapy in Asia patients with chronic hepatitis B.
Asia Hepatitis Lamivudine Study Group. Gasteroenterol 2000;
119:172-180.
7. Leung NW, Lai CL, Chang TT, et al. Extended lamivudine treatment in patients with chronic hepatitis B enhancers he- patitis B e antigen seroconversion rates: results after 3 years of therapy. Hepatology 2001;33:1527-1532.
8. Hadzyiyannis S, Tassopoulos NC, Heathcote EJ, et al. Long- term therapy with adefovir dipivoxil for HBe Ag-negative chronic hepatitis B. N Engl J Med 2005;352:2673-2681.
9. Fung SK, Chae HB, Fontana RJ, et al. Virologic response and resistance to adefovir in patients with chronic hepatitis B.
J Hepatol 2006;44:283-290
10. Blum HE, Galun E, von Weizsacker F, Wands JR. Inhibition of hepatitis B virus by antisense oligodeoxynucleotides. Lan- cet 1991;337:1230.
11. Offensperger WB, Offensperger S, Walter E, et al. In vivo inhibition of duck hepatitis B virus replication and gene expression by phosphorothioate modified antisense oligode- oxynucleotides. EMBO J 1993;12:1257-1262.
12. Carman W, Thomas H, Domingo E. Viral genetic variation:
hepatitis B virus as a clinical example. Lancet 1993;341:349- 353.
13. Weizsacker F, Wieland S, Kock J, Offensperger WB, et al.
Gene therapy for chronic viral hepatitis: ribozymes, antisense oligonucleotides, and dominant negative mutants. Hepatology 1997;26:251-255.
14. Winter G, Milstein C. Man-made antibodies. Nature 1991;
349:293-299.
15. McCafferty J, Griffiths AD, Winter G, Chiswell DJ. Phage antibodies: filamentous phage displaying antibody variable do- mains. Nature 1990;348:552-554.
16. Bird RE, Hardman KD, Jacobson JW, et al. Single-chain antigen-binding proteins. Science 1988;242:423-426.
17. Radziwill G, Tucker W, Schaller H. Mutational analysis of the hepatitis B virus P gene product: domain structure and RNase H activity. J Virol 1990;64:613-620.
18. Jeong JH, Kwak DS, Rho HM, Jung G. The catalytic properties of human hepatitis B virus polymerase. Biochem Biophys Res Commun 1996;223:264-271.
19. Park SG, Jeong YJ, Lee YY, et al. Hepatitis B virus- neutralizing anti-pre-S1 human antibody fragments from large naive antibody phage library. Antiviral Res 2005;68:109-115.
20. Lee SR, Song EY, Jeong YJ, et al. Development of human antibody inhibiting RNase H activity of polymerase of hepa- titis B virus using phage display technique. Immune Network 2004;4:16-22.
21. Tsurimoto T, Fujiyama A, Matsubara K. Stable expression and replication of hepatitis B virus genome in an integrated state in a human hepatoma cell line transfected with the
92 The Korean Journal of Gastroenterology: Vol. 49, No. 2. 2007
cloned viral DNA. Proc Natl Acad Sci USA 1987;84:444- 448.
22. Seifer M, Standring DN. Recombinant human hepatitis B virus reverse transcriptase is active in the absence of the nucleocapsid or the viral replication origin, DR1. J Virol 1993;67:4513-4520.
23. Winter G, Harris WJ. Humanized antibody. Immunol Today 1993;14:139-143.
24. Andrzejewski C, Young PJ, Goldman J, Spitainik SL, Silberstein LE. Production of human IgM warm reacting red cell monoclonal autoantibodies by Epstein-Barr virus trans- formation. Transfusion 1989;29:196-200.
25. Lonberg N, Taylor LD, Harding FA, et al. Antigen-specific human antibodies from mice comprising four distinct modi- fications. Nature 1994;368:856-859.
26. Winter G, Griffiths AD, Hawkins RE et al. Making anti- bodies by phage display technology. Ann Rev Immunol 1994;
12:433-455.
27. Barbas CF III, Burton DR, Scott JK, Silverman GJ. Phage display, a laboratory manual. New York: CSHL Press, 2001.
28. Gargano N, Biocca S, Bradbury A, Cattaneo A. Human recombinant antibody fragments neutralizing human immuno- deficiency virus type 1 reverse transcriptase provide an exper- imental basis for the structural classification of the DNA
polymerase family. J Virol 1996;70:7706-7712.
29. Park SG, Lee JS, Je EY, Kim IJ, Chung JH, Choi IH.
Affinity maturation of natural antibody using a chain shu- ffling technique and the expression of recombinant antibodies in Escherichia coli. Biochem Biophys Res Commun 2000;
275:553-557.
30. De Wildt RM, Hoet RM, van Venrooij WJ, Tomlinson IM, Winter G. Analysis of heavy and light chain pairings indi- cates that receptor editing shapes the human antibody reper- toire. J Mol Biol 1999;285:895-901.
31. Low NM, Holliger PH, Winter G. Mimicking somatic hyper- mutation: affinity maturation of antibodies displayed on bac- teriophage using a bacterial mutator strain. J Mol Biol 1996;
260:359-368.
32. Irving RA, Kortt AA, Hudson PJ. Affinity maturation of recombinant antibodies using E. coli mutator cells. Immuno- technology 1996;2:127-143.
33. Chowdhury PS, Vasmatzis G, Beers R, Lee B, Pastan I.
Improved stability and yield of a Fv-toxin fusion protein by computer design and protein engineering of the Fv. J Mol Biol 1998;281:917-928.
34. Kast P, Hilvert D. 3D structural information as a guide to protein engineering using genetic selection. Curr Opin Struct Biol 1997;7:470-479.