499
Copyright © 2016 The Korean Society of Fisheries and Aquatic Science pISSN:0374-8111, eISSN:2287-8815
서 론
대구
(Gadus microcephalus)
는대구목대구과에속하는우리 나라의대표적인상업적어종중하나로써우리나라황해와동 해를비롯한베링해및미국북서부해역을포함한북태평양해 역에광범위하게서식하며,
수온5-12℃
의수심10-550 m
에떼 를지어서식하는것으로알려져있다(Cohen et al., 1990).
우리나라대구어획량은
1990
년대낮은어획량을나타내었으나,
2000
년대부터증가하기시작하여2003
년1,826
톤에서2015
년7,820
톤으로증가하는경향을보였다(MOF, 2016).
우리나라 대구는 동해와 황해
2
개 계군으로 구분해왔다(Zhang, 1984; Seo et al., 2007).
황해대구는동해에비해크기 가작아보통38 cm
내외이고동해대구는보통68 cm
내외로최소성숙전장은동해계가수컷
58 cm,
암컷62 cm
이고황해계는수컷
32 cm,
암컷38 cm
로알려져있다(Chyung, 1977).
하지만최근연구된
Gwak and Nakayama (2011)
과Kim et al.
(2010), Seo et al. (2010)
의유전적분류연구에따르면우리나 라대구는동해,
황해,
동남해3
가지의계군으로나뉘며,
그중 황해대구는남해대구와유전적으로가까우나동해대구와는 뚜렷이구분이된다고보고하였다.
또한, Lee et al. (2015)
의표 지방류연구에따르면우리나라동해해역의대구는최소2
개 의계군이있는것으로추정되며다양한지역개체군이존재할 것으로보고하였다.
그러나서로다른계군사이에서도유전자교류는꾸준히일 어나며
,
계군이라는것은시간에따라고정된단위가아니며언 제든지새로운환경에적응하여변화하는역동적인개체군단 위임을명심할필요가있다.
개체군의표현형질은유전형질에 따라대부분결정되나,
같은유전자집단으로판별되는2
개개동해와 황해 대구(Gadus macrocephalus)의 생물학적 특성 비교
이경환·차형기
1·김영혜
2·이정용
3·정석근*
제주대학교 해양생명과학과, 1국립수산과학원 연근해자원과, 2국립수산과학원 남해수산연구소, 3국립수산과학원 양식관리과
Comparison of Biological Characteristics of Pacific Cod Gadus microcephalus between the East and Yellow Seas, Korea
Kyunghwan Lee, Hyung Kee Cha
1
, Yeonghye Kim2
, Jeong-Yong Lee3
and Sukgeun Jung*Department of Marin Life Science, Jeju National University, Jeju 63243, Korea
1
Coastal Water Fisheries Resources Research Division, National Institute of Fisheries Science, Busan 46083, Korea
2
Fisheries Resources and Environment Division, South Sea Fisheries Research Institute, National Institute of Fisheries Science, Yeosu 59780, Korea
3
Aquaculture Research Division, National Institute of Fisheries Science, Busan 46083, Korea
We investigated regional differences in the growth and maturation rate of Pacific cod in the East and Yellow Seas.
Significant regional differences were detected in the von Bertalanffy growth equation and mean length at maturity ( L
50). Cod in the East Sea grew at a faster rate than those in the Yellow Sea, with females growing faster than males in both populations. Cod of both sexes matured earlier in the Yellow Sea (age of maturity: 2.3 years for males, 2.6 years for females) than in the East Sea (age of maturity: 3.9 years for males, 4 years for females). These regional dif- ferences suggest that Pacific cod in the Yellow Sea, which is at the southwestern extremity of global cod distribution and thus likely to be an inhospitable habitat for this species, have adapted to their environment by developing earlier maturation and slower growth than cod in the East Sea or the Korea Strait. These regional differences must be taken into account when setting biological reference points for management of the Pacific cod fishery in Korean waters.
Key words: Pacific cod, Yellow Sea, East Sea, Growth, Maturation
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial Licens (http://creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
http://dx.doi.org/10.5657/KFAS.2016.0499 Korean J Fish Aquat Sci 49(4) 499-508, August 2016
Received 14 June 2016; Revised 22 August 2016; Accepted 28 August 2016
*Corresponding author: Tel: +82. 64. 754. 3424 Fax: +82. 64. 756. 3493
E-mail address: [email protected]
으며
,
반대로서로다른유전자집단이라고판별된공간적으로 멀리떨어진2
개개체군도비슷한표현형질을보일수도있다(McQuinn, 1997; Secor, 1999).
어류는산란기동안산란과자 어가성장하기에알맞은산란장을선택할수있으며,
서식하기 에알맞은수온이나 염분과같은환경조건이갖추어진 해역 으로이동할수있는능력을가지고있다(Royce, 1996). Secor
(1999)
에따르면어류들은알려진산란장내에서도해양환경이나산란지역의개체밀도에따라산란장을다양하게선택하 며
,
색이장과월동장선택에서개체들마다이상적인환경을찾 기 위해고정적이지 않고다양한 선택을한다고보고하였다(Hutchings, 1996). McQuinn (1997)
은여러해역의개체들이 회유에의해외부에서의개체유입과유출,
생존,
사망에의해 끊임없이변화하며,
해역간개체들의이동에의한유전적구성 또한고정적이지않고지속적으로변화하는개체군으로계군(stock)
보다는메타개체군(Metapopulation)
이라는용어를쓸 것을제안하였다.
따라서수산자원관리를위한생물학적기준 점을정할때는지역개체군들의유전형질차이뿐만아니라생 태환경차이로나타나는성장과성숙과같은표현형질차이도 종합적으로고려할필요가있다.
최근까지수산자원의평가와관리는환경은고려하지않고개 체군수준에서가입과성장
,
자연사망등네요소만을고려하는 개체군역학적분석결과에의존하여수행되어져왔다(Zhang, 2006; Gulland, 1983).
이러한전통적인수산자원평가방법론 에서는환경요인은장기적으로일정하다고가정하지만,
최근 에는지구온난화와기후변화등에따라이런장기평형가정을 더이상가정하기힘들다고보고있다(Haltuch et al., 2009).
더 욱이동해와황해는해양환경이크게다르기때문에이런해역 에따른환경차이를고려하지않은수산자원평가에따른생물 학적기준점을설정하여대구를관리한다면큰오류를범할수 있으므로환경차이와장기적변화를고려한평가와관리방안 이필요하다(Cha and Jung, 2012).
최근까지우리나라대구의 생물학적연구는동해(Seo et al., 2007; Lee et al., 2005; Cha et al., 2007),
황해(Kim et al., 2013),
남해(Choi and Gwak, 2011)
로나누어자치어를포함하는성숙과산란연구가진행했으며, Park and Gwak (2009), Lee et al. (2015), Yoon et al. (2012), Baeck et al. (2007)
은대구의식성연구와Kim et al. (2010), Seo et al. (2010), Gwak and Nakayama (2011)
은유전자분석 연구, Lee et al. (2015)
의표지방류를이용한회유방향연구가 진행되었다.
우리나라대구연구는대부분동해와남해를중심으로했으 며
,
황해대구에대한연구는부족한상황이다. Park and Gwak (2009)
이연구한해역별위내용물비교, Seo et al. (2010)
와Kim et al. (2010)
의해역별유전적연구를통하여해역에따른 식성과유전자분포차이는일부평가되었지만성장과성숙비 교에대한연구는전무한상황이다.
따라서,
본연구는선행연구의성장과성숙을계량적으로비교하였으며
,
수산자원의해 역별성장과성숙비교연구를위한분석방법과해역별해양생 태계와대구의지역적특성을고려한대구수산자원관리방안을 마련하는데목적을두고있다.
자료 및 방법
성장, 성숙 비교 자료
동해와황해대구의성장과성숙을평가하기위하여동해대 구성장평가자료는
Lee et al. (2005)
가추정하고보고한자료 를이용하였다.
이자료는Gwak and Nakayama (2011)
와Lee
et al. (2015)
의연구에의해죽변을기준으로동해북부해역의강원도주문진연안과양양군연안에서
2003
년1
월부터12
월까지매월어획된
261
개의개체의이석(otolith)
길이와전장(TL),
체장이측정된자료를이용하였다(Lee et al., 2005; Jung et al., 2009).
군성숙체장(L
50)
평가에서는Cha et al. (2007)
가 측정한2006
년1
월부터2007
년2
월까지우리나라동해북부 해역의강원도인근해역에서어선으로어획된1960
개체자료 를활용하였다.
분석에사용된자료는전장은0.1 cm
단위,
체 중(BW)
과생식소중량(GW)
은0.01 g
까지구분하였으며,
성숙 도는생식소크기,
색조,
난립의크기,
난의투명상태를기준으 로하여,
미숙(immature),
중숙(maturing),
완숙(mature),
방후(spent)
로구분된자료를이용하였다(Cha et al., 2007).
황해대구개체군의성장과성숙은
Kim et al. (2013)
가측정 한2007
년1
월부터2
월을제외한12
월까지우리나라황해안중 부해역에서어획된682
개체자료를이용하였다.
이자료는이 석(otolith)
길이와전장(TL)
은0.1 cm
단위,
체중(BW)
과생식 소중량(GW)
은0.1 g
단위까지측정하였다(Kim et al., 2013).
성장식
각개체의나이는이석연륜으로연령을추정하였으며
(NFR- DI, 2005),
동해와황해대구모두매년1
월1
일을부화일로가 정하여윤문수와어획된날까지일별연령으로계산하였다.
윤 문형성기간에는연령추정에1
년의오차가날수있으므로이석
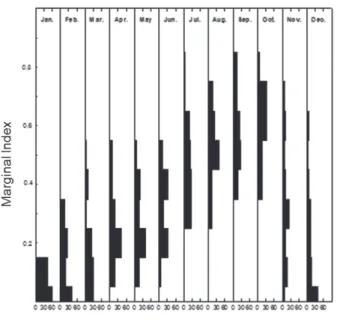
Marginal index
의월별분포변화로부터윤문형성시기를추정하였고
,
윤문형성초기에해당하는시기의자료는제외하 고분석하였다.
황해는Kim et al. (2013)
이보고한윤문형성시기인
12-3
월자료를제외하였으며,
동해는윤문길이분석을통하여추정한윤문이발생하기시작하는
11-3
월자료를제외하였다
(Fig. 1).
본버틀란피(von Berterlanffy)
방정식을이용하 여해역별대구개체의성장을비교하였으며그방정식은다 음과같다.
L
t= L
∞×(1-e
-K(t-t0)) ··· (1)
여기서
L
t는시간t
에서체장, L
∞는해당개체의이론적최대 체장, K
는성장계수, t
0는길이가0
일때의이론적인시간을의 미한다.
여기서는Seo et al. (2007)
이보고한부화직후길이(L
0)
를이용하여t
0를도출하였으며그식은다음과같다.
t
0= 1
×log(1- L
0)··· (2)
K L
∞도출된
t
0를이용하여2
개의파라미터(L
∞, K)
로이루어진방 정식으로변형후성장을비교하였다.
L
t=L
∞×(1-e
-K(
t-(1×log(1-L0 ))
)
)···(3)
K L∞
성장식파라미터는
R
통계프로그램(Crawley, 2012)
에서최 소자승법에기반한비선형회귀분석(nls)
를통하여도출하였다. 성장식 비교
본버틀란피성장식을이용하여도출한해역별암컷과수컷
대구성장식은연령별평균체장을이용하는
Kimura’s likeli-
hood ratio
검정과일단위체장자료를이용하는Wald
검정을 이용하여동해와황해대구성장차이를비교하였다.
Kimura’s likelihood ratio 검정
두집단비교시각연령별평균의자료와최소자승법을통하
여집단의성장곡선이그려지며도출된성장곡선을카이제곱 검정을통하여비교하였다
.
카이제곱값을도출하는식은다음 과같다.
χ
κ2= -N·ln [ RRS
Ω]···(4) RRS
ω여기서
K
는비교하고자하는성장식의총자유도이며N
은총개체수
, RRS
Ω는각지역별성장식에따른관측값과기대값차이
(
집단내잔차,
동·
황해잔차의합)
제곱의합을나타내 며RRS
ω는설정된가설에따른전체개체의관측값과기대값 차이(
집단전체 잔차)
제곱의 합을 나타낸다. Kimura (1980)
에 따른 검정 가설은 다음과같다. ①L
∞,1/K
1/t
0,1=L
∞,2/K
2/t
0,2②L
∞,1=L
∞,2③K
1=K
2④t
0,1=t
0,2⑤L
∞,1/K
1= L
∞,2/K
2⑥L
∞,1/t
0,1= L
∞,2/t
0,2⑦K
1/t
0,1= K
2/t
0,2의귀무가설검정을통하여성장곡 선의차이여부가결정되며, ①
귀무가설채택시두성장곡선은 동일하다고판정내릴수있다.
만약①
의귀무가설이기각이될 경우②-⑦
의귀무가설검정을통하여결과를해석한다.
본연 구에서는문헌조사에의해확인된L
0값을통하여도출한t
0값 을이용하였으므로2
개의파라미터비교가설인①L
∞,1/t
0,1= L
∞,2/t
0,2②L
∞,1=L
∞,2③K
1=K
2의귀무가설에대하여동해와황 해대구성장차이를검정하였다(Haddon, 2001).
Wald 검정
각해역별성장식파라미터는통상최소자승법
(OLS)
로도출 되며도출된파라미터의값을카이제곱값에따라SAS 9.3
통 계프로그램을이용하여두성장곡선의동일성을검정하였다(Stewart, 2009).
개체군 성숙
동해대구의산란기는동일한
Cha et al. (2007)
의자료를이 용하여성숙을연구한Jung et al. (2009)
의연구와같은산란 시기를적용하였으며,
황해는12-1
월(Kim et al., 2013)
로추 정하였다.
산란기에해당하는 동해대구에서생식소숙도지수(gonadosomatic index, GSI)
가5
보다큰개체들이성숙을이 루었으며,
황해또한Kim et al. (2013)
이연구한월별생식소숙 도지수분포에서산란기는생식소숙도지수가5
이상으로나타 나동해와황해대구모두생식소숙도지수5
를기준으로성숙 과미숙을평가하였다.
생식소숙도지수
(GSI)
를도출하는식은다음과같다. GSI= GW
× 10
3··· (5) BW
체장에따른대구성숙확률을다음로지스틱방정식을이용 하여도출하였다
.
Fig. 1. Monthly changes in the marginal index of Gadus macro- cephalus collected in the East Sea from January to December 2003.
P(X)= ··· (6) 1+e
α+βx여기서
P(X)
는체장이X
일때개체의성숙확률을나타내며α
와β
는각각직선회귀식에서절편과기울기를나타낸다.
각해 역별,
성별개체의군성숙체장(L
50)
은도출된파라미터α
와β
를 이용하여다음과같이계산한다(Chen and Paloheimo, 1994).
L
50= - α
···(7) β
여기서 파라미터는
R
통계 프로그램 최소자승법을 기반한일반화 선형 회귀분석
(glm)
을 통하여 도출하였다(Crawley,
2012).
군성숙체장(L
50) 비교
두 해역의성별군성숙체장
(L
50)
의 차이를검정하기 위하여Loretta (1999)
이제안한절차에따라L
50 의분산을추정한뒤 적절한t
검정을선택하여해역별L
50 차이를검정하였다.
해역별성별
L
50의분산을추정하기위하여Ashton (1972)
의연구방법에따라사용된각성별자료는
5 cm
단위로묶은후각그룹당실제성숙과미숙개체의수를이용하여다음의식을 통하여분산을추정하였다
(Fig. 2).
S
2(y) = 1
[S
2(a')+(y- -χ)
2×S
2(b)]··· (8) b
2여기서
y
는군성숙체장L
50, S
2(y)
는각해역별,
성별로예측된L
50의분산이며, S
2(b)
는각각회귀분석을통해도출된파라미 터b
의분산, S
2(a')
와-χ
는아래의식을통하여도출하며각식 에서n
과P, Q, χ
는개체수와그룹당성숙률,
미성숙률,
그룹대 표값(
중앙값)
을나타낸다.
S
2(a')=1/∑nPQ ··· (9)
Fig. 2. Length distribution of Gadus macrocephalus in the East (from January to February 2007) and the Yellow Sea (from January to De- cember 2007). (a) Female cod in the East Sea; (b) Male cod in the East Sea; (c) Female cod in the Yellow Sea; (d) Male cod in the Yellow Sea.
χ =∑nPQ/∑nPQ ···(10) -
성별 동해와 황해 군성숙체장 등분산 검정과 차이검정
추정된해역별성별L
50의분산은F
검정을통하여등분산여 부를확인하였으며,
분산이서로다른것으로판정되었을경우,
그차이여부를t
검정으로판정하였으며, t
값은다음식에따라 도출하였다(Seong, 2007).
t = L
50,1-L
50,2··· (11) σ
12+ σ
22n
1n
2여기서분자는각해역별개체의
L
50 차이를의미하는데L
50,1 은표본1
의군성숙체장, L
50,2는표본2
의군성숙체장을말하며, σ
12, σ
22는각모집단1
과2
의L
50분산을나타내고n
1, n
2는표본1
과2
의개체를나타낸다. t
값의유의성여부를판단하기위한자유도는다음공식의역수로도출하였다
(Seong, 2007).
1 = 1
×(1- w
1)
2+ 1
×(1- w
2)
2··· (12)
df df
1w df
2w
여기서
df
1과df
2는각각해당그룹의자유도를나타내며각해 당그룹의자유도는n
1-1, n
2-1
로구해진다. w
1, w
2와w
는아래 의공식으로유도된다(Seong, 2007).
w
i= n
i/σ
i2···(13)
w=w
1+w
2···(14)
결 과
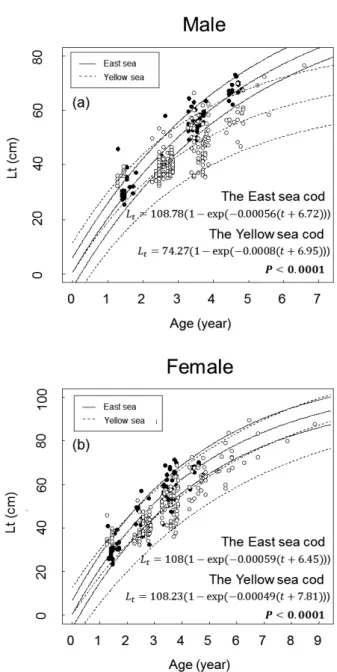
성장 성장식
나이를연단위로놓고
,
연령별로평균을낸체장자료로도출한해역별암컷과수컷의대구성장식은다음과같다
(Fig. 3).
동해수컷
(n=78): L
t=127.56(1-e
(-0.17(t+0.019)))
동해암컷(n=87): L
t=106.25(1-e
(-0.22(t+0.018)))
황해수컷(n=235): L
t=94.13(1-e
(-0.23(t+0.019)))
황해암컷(n=238): L
t=103.91(1-e
(-0.21(t+0.019)))
나이를일단위까지추정하여도출한해역별암컷과수컷대 구의성장식은다음과같다
(Fig. 4).
동해수컷
(n=78): L
t=108.78(1-e
(-0.00056(t+6.72)))
동해암컷(n=87): L
t=108(1-e
(-0.00059(t+6.45)))
황해수컷(n=235): L
t=74.27(1-e
(-0.0008(t+6.95)))
황해암컷(n=238): L
t=108.23(1-e
(-0.00049(t+7.81)))
Fig. 3. von Bertalanffy growth curve of Gadus macrocephalus de- rived from the averaged lengths of wach age class in the East and the Yellow Sea. (a) Male; (b) Female.
Wald
검정과Kimura’s likelihood ratio
검정에따라두해역의 대구성장식을비교한결과두검정모두동해와황해수컷과암 컷대구의성장은유의한차이를나타내었으며,
황해개체보다 동해개체가전연령에서체장이더크고두해역모두수컷보다 암컷이더큰특징을나타내었다(Table 1-4, P<0.05).
성숙
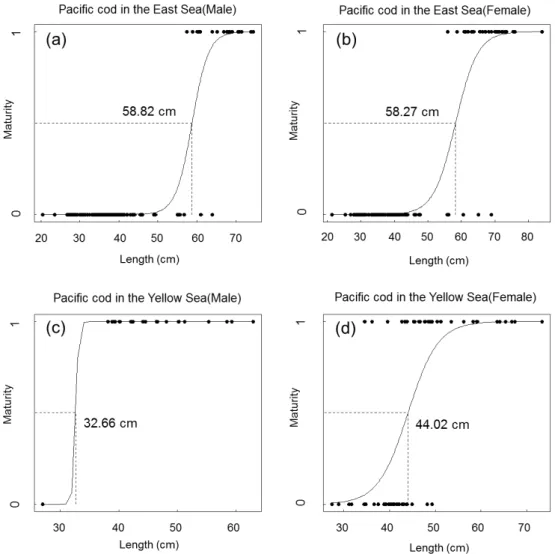
로지스틱 회귀분석과 군성숙체장
로지스틱회귀분석으로도출한해역별대구수컷과암컷의성
동해수컷
: P(X)= e
-27.87+0.47x1+e
-27.87+0.47x동해암컷
: P(X)= e
-17.82+0.31x1+e
-17.82+0.31x황해수컷
: P(X)= e
-133.55+4.09x1+e
-133.55+4.09x황해암컷
: P(X)= e
-13.5+0.31x1+e
-13.5+0.31x해역별길이에따른군성숙체장
L
50 은동해수컷58.82 cm,
암 컷58.27 cm,
황해수컷32.66 cm,
암컷44.02 cm
로나타났다(Fig. 5).
도출된성장식을이용하여해역별연령추정시동해수컷
3.9
세,
동해암컷4
세,
황해암컷2.6
세로나타나며,
동해 대구는4
세부터성숙시기로추정된다른연구의보고(NFRDI,
2010)
와비교하여비슷한결과를나타내었다.
해역별대구L
50은수컷과암컷모두동해가황해보다크게나타났다
. 군성숙체장 분산 도출
Ashton (1972)
의분산추정방법과R
통계프로그램을이용 하여추정한해역별성별L
50 분산은Table 5
과같다.
동해암컷 의경우동해수컷과황해암컷에비교하여분산이더크게나타났다
.
하지만표준편차(
동해암컷: ±2.17)
로확인한다면동해수컷
(±1.31)
과황해암컷(±1.4)
과비교하여차이가크지않았 다.
황해수컷은자료부족으로분산을추정할수없어암컷에 국한하여해역별L
50 차이를비교하였다.
t 검정(L
50)
등분산검정결과동해와황해암컷
L
50 의분산은해역별로 다른것으로판정내렸으며,
분산이다른경우를가정한t
검정 을이용하여동해와황해대구암컷의L
50 차이를검정하였다.
그결과암컷의L
50은 통계적으로유의한해역간차이를나타내 었으며,
동해가황해보다더큰체장에서성숙하였다(Table 6).
고 찰
우리나라 동해와 황해 대구의 성장과 성숙 비교
도출된각각의본버틀란피성장식을
Wald
검정과Kimura’s
likelihood ratio
검정을통하여유의한차이를확인하였다.
추정 된성장식을통하여대구평균연령인8
세(Jung et al., 2009)
까 지의체장변화를예측해보면동해가황해보다체장이더크며 수컷보다암컷이더큰특징을나타내었다(Fig. 4).
Fig. 4. von Bertalanffy growth curve of Gadus macrocephalus de- rived from daily lengths of the otoliths collected in the East and the Yellow Sea. (a) Male; (b) Female.
해역별성숙추정연령은동해수컷
3.9
세,
동해암컷4
세,
황해 암컷2.6
세로황해대구가더빠른성적성숙을나타내었다.
동 해대구는4
세부터성숙시기로추정된다른연구의보고(NFR-
DI, 2010)
와비교하여비슷한결과를나타내었다.
동해와황해암컷의군성숙체장
(L
50)
은유의한차이를나타내며동해해역 의개체가더큰체장에서성숙이이루어지는것으로나타났다.
Stark (2007)
의베링해와알래스카만개체군에대한성장과성숙연구보고에따르면해역별암컷개체군으로한정하여추 정한최소성숙체장과나이는알래스카만
50.3 cm,
베링해58 cm
로추정연령은약4.4
세와약4.9
세로보고하였다.
베링해 와알래스카만대구또한우리나라동해와황대대구성장과성 숙연구결과와마찬가지로해역에따른차이를나타내었으며,
베링해해역의개체가더큰체장에서성숙이이루어지는것으 로나타났다.
베링해와알래스카만대구개체군의성장과성숙을우리나라
동해와황해개체군과비교해볼때암컷에한정한해역별성숙 연령은우리나라대구가더빠른것으로
,
성숙체장은동해대 구와비슷하거나작은것으로나타났다.
또한,
두해역의암컷 과수컷개체군의성장을비교하면이론적최대체장은베링해 와알래스카만의개체군이크지만성장계수가낮은값의결과 를나타내어우리나라대구가연령에따른성장이더빠른것으 로나타났으며,
황해와알래스카만대구는동해와베링해대구 보다작은체장에서더빠른성숙을하는공통된특징을나타내 었음을짐작할수있다.
우리나라 동해와 황해 대구 먹이환경과 영향
우리나라동해와황해대구의
L
50과성숙추정연령은황해대 구가작고빠른성숙을하는것으로나타났으며,
이러한차이는 해역별먹이환경차이가우리나라동해와황해대구의성장과 성숙에영향을끼치는한가지의요인인것을보인다.
Fig. 5. The derived logistic curves and the estimated mean maturity length for Gadus macrocephalus by sex in the East Sea and the Yellow Sea.
Yoon et al. (2012), Baeck et al. (2007)
그리고Park and
Gwak (2009)
의식성연구에따르면진해만을제외한우리나라해역별대구의먹이중요도는새우류
,
어류,
두족류순서로보 고했으며, Yamamura et al. (1993)
와Yang (2004)
이보고한일 본훗카이도와베링해대구의식성연구결과에따르면공통적 으로새우류를주요먹이생물로보고하였으며,
이외에조개류,
게류를주로먹는다고보고하였다.
또한,
우리나라대구위내 용물과차이점으로어류중우리나라대구개체위내용물에서 발견되지않은명태를섭이하는것으로보고하였으며,
훗카이 도대구는체장30 cm,
베링해대구는체장60 cm
이상에서명 태를섭이하는것으로보고하였다.
이러한해역별먹이생물차이는각해역의먹이생물환경에따라나타난차이로추측된다
. Yoon et al. (2012), Baeck et al. (2007), Park and Gwak (2009), Lee et al. (2015), Yamamura et al. (1993), Yang (2004)
에따르면해역별대구는체장이증가하면서각먹이생물비 율이 변화하는 공통적인 특징을나타내었으며, Baeck et al.
(2007)
과Yoon et al. (2012)
는우리나라대구는계절별로식성 차이가난다고보고하였다.
body length of each group, K is von Bertalanffy growth coefficient of each group and t0 is age of cod at 0 length Hypothesis Linear
constraints L∞,1 L∞,2 K1 K2 t0,1 t0,2 Residual sum of
squares(E) Residual sum of
squares(Y)
χ
2 PH0 None 127.56 94.13 0.17 0.23 -0.019 -0.019 - - - -
H1 L∞,1=L∞,2 100.04 100.04 0.23 0.2 -0.019 -0.019 21.51 112.88 18.32 <0.001 H2 K1=K2 106.78 98.03 0.21 0.21 -0.019 -0.019 17.99 111.72 19.76 <0.001 H3 L∞,1=L∞,2
K1=K2 92.02 92.02 0.25 0.25 -0.019 -0.019 45.91 122.38 12.99 0.0015(<0.05) Table 2. Kimura’s likelihood ratio test of the growth equation between the East and the Yellow Sea cod (female). L∞ is theoretical maximum body length of each group, K is von Bertalanffy growth coefficient of each group and t0 is age of cod at 0 length
Hypothesis Linear
constraints L∞,1 L∞,2 K1 K2 t0,1 t0,2 Residual sum of
squares(E) Residual sum of
squares(Y)
χ
2 PH0 None 106.25 103.91 0.22 0.21 -0.018 -0.019 - - - -
H1 L∞,1=L∞,2 104.05 104.05 0.23 0.21 -0.018 -0.019 1.86 93.13 47.2 <0.001
H2 K1=K2 110.12 103.49 0.21 0.21 -0.018 -0.019 1.98 93.15 46.46 <0.001
H3 L∞,1=L∞,2
K1=K2 99.62 99.62 0.23 0.23 -0.018 -0.019 14.25 102.68 25.25 <0.001
Table 3. Wald test of the growth equation between the East and the Yellow Sea cod (male)
East Sea Yellow Sea
χ
2L∞ 108.78 74.23
P <0.0001
K 0.21 0.29
Table 4. Wald test of the growth equation between the East and the Yellow Sea cod (female)
East Sea Yellow Sea
χ
2L∞ 108 108.22
P <0.0001
K 0.22 0.18
Table 5. Variance of mean maturity length (L50) of male and female cod in the East and the Yellow Sea
East Sea Yellow Sea
Male Female Male Female
L50 58.82 cm 58.27 cm 32.66 cm 44.02 cm
Var (L50) 1.73 4.69 - 1.97
Sample size 172 150 52 62
Table 6. Result of t test of mean maturity length between the East and the Yellow Sea female cod
Standard error (SE) 0.25
Degree of freedom (df) 171
t value 56.77
P value < 0.0001