Protective Effects of Yuzu and Its Component
against Cardiovascular Disease
by
Hye Yon Yu
Major in Molecular Medicine
Department of Biomedical Sciences
The Graduate School, Ajou University
Protective Effects of Yuzu and Its Component
against Cardiovascular Disease
by
Hye Yon Yu
A Dissertation Submitted to The Graduate School of
Ajou University in Partial Fulfillment of the Requirements for
The Degree of Master of Biomedical Sciences
Supervised by
Soo Hwan Lee, Ph.D.
Yi-Sook Jung, Ph.D.
Major in Molecular Medicine
Department of Biomedical Sciences
The Graduate School, Ajou University
This certifies that
of Hye Yoⅱ
Yu
the dissertation
is approved●
The Gradμ
ate
December,
S UPERV工
SORY COMMITTEE
Schoo1; Aj˚
u
19th, 2014
i
-ABSTRACT-
Protective Effects of Yuzu and Its Component
against Cardiovascular Disease
Left ventricular (LV) remodeling, which includes ventricular dilatation and increased
interstitial fibrosis after myocardial infarction (MI), is the critical process underlying the
progression to heart failure (HF). Therefore, a novel approach for preventing LV remodeling
after MI is highly desirable. Yuzu is a citrus plant originating in East Asia, and has a number
of cardioprotective ingredients such as hesperidin. However, no study has proved whether
yuzu can prevent LV remodeling. The aim of this study was to determine the effects of yuzu
on HF and its potential impact on the LV remodeling process after MI and platelet
aggregation. In this study, it was investigated whether yuzu and its components hesperidin
and naringin, have anti-platelet activities. Yuzu and hesperidin inhibited collagen-,
arachidonic acid (AA)-, ADP- and thrombin-induced rat platelet aggregation in vitro and ex
vivo. Naringin also inhibited platelet aggregation induced by collagen, AA, or thrombin, but
not by ADP. The oral administration of yuzu or hesperidin prolonged mouse tail vein
bleeding time in a dose-dependent manner. LV remodeling in vivo study using the
permanent left anterior descending coronary artery (LAD) occlusion model demonstrated
ii
occlusion significantly attenuated cardiac dysfunction, myocyte apoptosis and inflammation.
Not only yuzu but also hesperidin inhibited caspase-3 activity, myeloperoxidase expression,
α-smooth muscle actin expression, and matrix metalloproteinase-2 activity in a permanent LAD occlusion rat model. These results suggest that yuzu and hesperidin have anti-platelet
activity, and that intake of yuzu, which contains various flavonoids such as hesperidin, may
be beneficial for individuals at high risk of cardiovascular diseases. These findings provide
the first evidence that yuzu and hesperidin prevent MI-induced ventricular dysfunction and
structural remodeling of myocardium.
Key words : yuzu, hesperidin, platelet aggregation, cardiomyocyte, ischemic heart disease,
iii
TABLE OF CONTENTS
ABSTRACT ... i
TABLE OF CONTENTS ... iii
LIST OF FIGURES ... vii
LIST OF TABLES ... ix
ABBREVIATION ... x
I. INTRODUCTION ... 1
A. Antiplatelet therapy in cardiovascular diseases ... 1
B. Left ventricular (LV) remodeling and heart failure (HF) ... 1
C. Yuzu and its components ... 3
D. Aims of study ... 4
II. MATERIALS AND METHODS ... 5
A. Materials ... 5
1. Materials ... 5
B. Methods ... 5
1. Plant material and preparation of samples ... 5
iv
3. Animals ... 7
4. Preparation of platelets ... 7
5. In vitro platelet aggregation study ... 8
6. Determination of cytotoxicity... 8
7. Ex vivo platelet aggregation study ... 9
8. In vivo mice tail bleeding times ... 9
9. Measurement of thromboxane A2 (TXA2) formation ... 10
10. Echocardiography ... 10
11. Histological analysis ... 11
12. Immunohistochemistry ... 11
13. Gelatin zymography ... 11
14. Western blot analysis ... 12
15. Terminal dUTP nick end-labeling (TUNEL) staining ... 12
16. Statistical analysis ... 13
III. RESULTS ... 14
A. PART I. Anti-platelet effects of yuzu extract and its component ... 14
v
2. Contents of hesperidin and naringin in yuzu extracts ... 17
3. Effects of hesperidin and naringin on platelet aggregations in vitro ... 20
4. Cytotoxic effects of yuzu, hesperidin, and naringin on platelets ... 23
5. Effects of yuzu, hesperidin, and naringin on TXB2 formation ... 25
6. Effects of yuzu and hesperidin on platelet aggregation ex vivo ... 27
7. Effects of yuzu and hesperidin on tail bleeding times in mice ... 29
B. PART II. Preventive effect of yuzu and hesperidin on LV remodeling and dysfunction in rat permanent LAD occlusion model ... 31
1. Yuzu and hesperidin prevented LV remodeling and functional deterioration following MI ... 31
2. Yuzu and hesperidin prevented myocardial fibrosis during LV remodeling after chronic MI ... 35
3. Yuzu and hesperidin reduced the inflammatory reaction and MPO expression during LV remodeling after chronic MI ... 38
4. Yuzu and hesperidin block myocardial apoptosis through inhibition of caspase-3 activation during LV remodeling after chronic MI ... 41
5. Yuzu and hesperidin block LV wall thinning through inhibition of MMP-2 activation during LV remodeling after chronic MI ... 44
vi
IV. DISCUSSION ... 47
A. PART I. Anti-platelet effects of yuzu extract and its component ... 47
B. PART II. Preventive effect of yuzu and hesperidin on LV remodeling and dysfunction in rat permanent LAD occlusion model ... 51
V. CONCLUSION ... 55
REFERENCES ... 56
vii
LIST OF FIGURES
Fig. 1. Dose-dependent inhibitory effect of yuzu on in vitro platelet aggregation ... 15
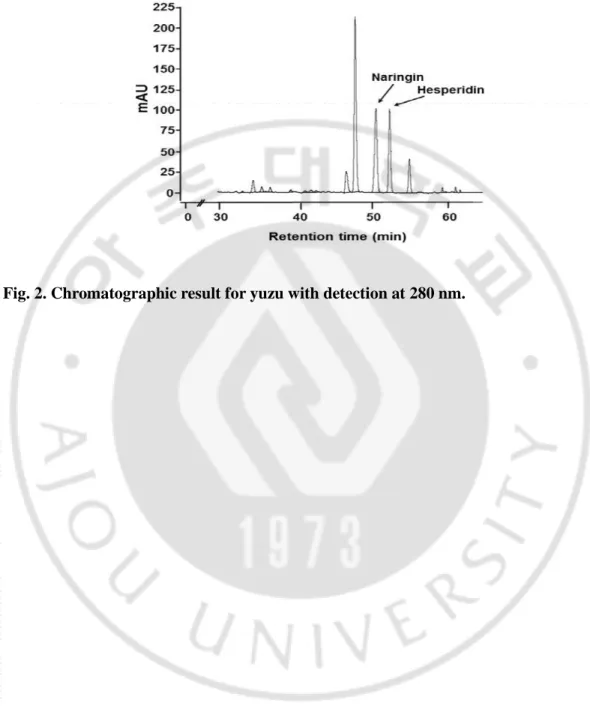
Fig. 2. Chromatographic result for yuzu with detection at 280 nm. ... 19
Fig. 3. Dose-dependent inhibitory effect of hesperidin on in vitro platelet aggregation ... 21
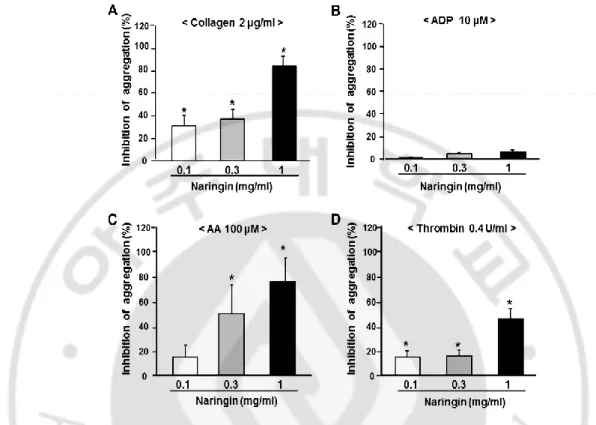
Fig. 4. Dose-dependent inhibitory effect of naringin on in vitro platelet aggregation ... 22
Fig. 5. Effect of yuzu, hesperidin, and naringin on collagen- and thrombin-induced TXB2 formation in rat platelets ... 26
Fig. 6. Effects of yuzu and hesperidin on mouse tail bleeding times ... 30
Fig. 7. Experimental schedule ... 32
Fig. 8. Effect of pre-treatment with yuzu or hesperidin on cardiac dysfunction ... 33
Fig. 9. Effect of pre-treatment with yuzu or hesperidin on LV dysfunction and LV remodeling ... 34
Fig. 10. Effect of pre-treatment with yuzu or hesperidin on cardiac fibrosis ... 36
Fig. 11. Effect of pre-treatment with yuzu or hesperidin on myofibroblast ... 37
Fig. 12. Effect of pre-treatment with yuzu or hesperidin on inflammatory reaction ... 39
viii
Fig. 14. Effect of pre-treatment with yuzu or hesperidin on cardiomyocyte apoptosis ... 42
Fig. 15. Effect of pre-treatment with yuzu or hesperidin on caspase-3 activity ... 43
Fig. 16. Effect of pre-treatment with yuzu or hesperidin on cardiomyocyte preservation ... 45
ix
LIST OF TABLES
Table 1. IC50 of yuzu, hesperidin, and naringin for platelet aggregation ... 16
Table 2. Contents of hesperidin and naringin in extract of yuzu ... 18
Table 3. Effects of yuzu, hesperidin, and naringin on LDH release by platelets ... 24
Table 4. Inhibitory effects (%) of orally administered yuzu and hesperidin on ex vivo platelet
x
ABBREVIATION
AA arachidonic acid
ADP adenosine diphosphate
BSA bovine serum albumin
cTnI cardiac troponin I
CVD cardiovascular disease
DMSO dimethylsulfoxide,
HF heart failure
LAD left anterior descending
LV left ventricular
MI myocardial infarction
MMP matrix metalloproteinase
MPO myeloperoxidase
PEG polyethylene glycol
PO permanent LAD occlusion
SMA smooth muscle actin
1
I. INTRODUCTION
A. Antiplatelet therapy in cardiovascular diseases
Platelets are essential for primary hemostasis and for repair of the endothelium, but they
are also responsible for the formation of pathogenic thrombi that cause cardiovascular
diseases (CVDs), such as acute coronary syndrome, ischemic stroke and symptomatic
peripheral artery disease. Platelets are also a source of inflammatory mediators, and their
activation by inflammatory triggers may be a critical component of atherothrombosis
(Jennings, 2009). Accordingly, the inhibition of platelet hyperactivation has been adopted as
a strategy to treat these disorders, and several anti-platelet drugs, including aspirin, are used
clinically. Since these anti-platelet drugs have all been reported to have adverse side effects,
the development of safe new therapeutic agents with anti-platelet activity remains a critical
issue. Interestingly, several natural compounds from plant sources, such as, spinach and
tomato, have been reported to have preventive effects against CVDs by inhibiting platelet
aggregation (O’Kennedy et al, 2006).
B. Left ventricular (LV) remodeling and heart failure (HF)
LV remodeling is a pathologic change in the architecture of the LV that occur due to
various CVDs including myocardial infarction (MI) and hypertension (Yi et al, 2012). Of
these, MI is caused by the partial interruption or occlusion of the blood supply to a part of
2
the rupture of a vulnerable atherosclerotic plaque (Gao et al, 2010) LV remodeling after MI
is associated with a combination of pathologic conditions, including myocyte hypertrophy,
myocyte apoptosis, myofibroblast proliferation, inflammatory reaction, and interstitial
fibrosis, which ultimately lead to the loss of systolic and diastolic function (Konstam et al,
2011). Cardiac hypertrophy is a compensatory process in response to increased
hemodynamic overload, characterized by an increase in the size of individual cardiac
myocytes and wall thickness. On the other hand, in chronic MI following left anterior
descending coronary artery (LAD) occlusion, a transition occurs from compensatory cardiac
hypertrophy to decompensatory hypertrophy, characterized by a chamber dilation and wall
thinning. In this chronic condition, processes such as extracellular matrix turnover, fibrosis,
inflammation and apoptosis are crucial determinants (Eapen and Rogers, 2009 ; . Yi et al,
2012)
LV remodeling after MI is a key contributor to HF, which is one of the most common
causes of cardiovascular morbidity and mortality worldwide (González et al, 2011). HF is
defined as a clinical syndrome because of its complex pathologic mechanism changes that
contribute to myocardial dysfunction following LV remodeling (Nagarajan and Tang, 2011).
Conventional HF therapy is still largely based on targeting the causes and neurohumoral
activation of HF, and includes agents such as angiotensin-converting enzyme inhibitors,
angiotensin-receptor antagonists, beta-blockers, and aldosterone antagonist (Jessup and
3
C. Yuzu and its components
Recently, natural products have become popular worldwide and have gained wide
acceptance as adjuncts to conventional therapy. Various studies have shown natural products
such as citrus fruits, grape, broccoli, and cacao are rich sources of phytochemicals such as
polyphenols that are well known for their cardiovascular protective effects (Akhlaghi and
Bandy, 2010; Yamazaki et al, 2010; Kim et al, 2014). Furthermore, numerous researches
indicate that the consumption of flavonoid-rich foods decrease the incidence of CVDs
(Yamazaki et al, 2008).
In particular, yuzu (Citrus junos sieb ex Tanaka) is one of the most famous natural
products. Yuzu is small tree that produces yellow-golden colored citrus fruits native to
northeast Asia, including Korea, China, and Japan. Yuzu is used in traditional Chinese
medicine and yuzu tea is known to improve a cold in Korea. Several studies have shown that
components of yuzu such as limonene, vitamin C, phenolic substances exhibit an antioxidant
and anti-inflammatory activities (Hirota et al, 2010).
Like other citrus fruits, yuzu contains many biofunctional components, such as,
flavonoids, carotenoids and ascorbic acid (Yoo et al., 2009). Furthermore, numerous
experimental studies have shown flavonoids inhibit primary hemostasis and many pathways
which are associated with platelet activation and aggregation (Rein et al., 2000). Flavonoids,
which occur in the free form and as glycosides, are a large group of low molecular weight
4
active compounds. Hesperidin and naringin are known to be major flavonoid components in
yuzu (Yoo et al, 2009). Hesperidin, a glycosidic form of hesperetin, is encountered
extensively in the plant kingdom, especially in citrus fruits, such as, grapefruits and oranges,
which are also commonly used in traditional medicines (Garg et al, 2001). Naringin is a
glycosidic form of naringenin, and the major flavanone found in citrus fruits. In animal
experiments, naringenin consumption was found to be associated with lipid-lowering effects,
reduced plasma markers of endothelial dysfunction, and improved insulin sensitivity (Chanet
et al, 2012). However, no report has been issued on its inhibitory effect on platelet
aggregation and LV remodeling.
D. Aims of study
1. In the present study, it was investigated the anti-platelet activities of the yuzu and its
major flavonoid components, in order to explain their possible contributions to the
attenuation of platelet hyperactivity.
2. The goal of the present study was to evaluate the effects of yuzu in a rat model of LV
remodeling induced by permanent LAD occlusion. Considering that hesperidin is
well-known major functional component of yuzu, it has also evaluated whether hesperidin
5
II. MATERIALS AND METHODS
A. Materials
1. Materials
Collagen, ADP, thrombin, arachidonic acid (AA), and luciferin/luciferase reagent were
purchased from Chrono-Log Co. (Harvertown, PA, USA). Dimethylsulfoxide (DMSO),
polyethylene glycol (PEG), bovine serum albumin (BSA), β-nicotinamide adenine dinucleotide (reduced disodium salt hydrate, β-NADH), pyruvic acid, and histone H1-IIIS
were purchased from Sigma (St. Louis, MO, USA). Thromboxane B2 (TXB2) enzyme
immunosaasy (EIA) kit was purchased from Cayman Chemical Company (Ann Arbor, MI,
USA).
B. Methods
1. Plant material and preparation of samples
Extract of yuzu were obtained from Konkuk University (Seoul, Republic of Korea).
Briefly, minced yuzu fruits were extracted with ethanol and lyophilized to remove solvent.
Yuzu extract was dissolved in saline (0.9% NaCl) for the in vitro, ex-vivo and in vivo study.
Yuzu major components, hesperidin and naringin are water-insoluble, they were dissolved in
DMSO to make final concentration of 0.1% DMSO for the in vitro platelet aggregation
6
a widely used solvent for water-insoluble compounds for in vivo study.
2. Experimental protocol
PART I: For platelet aggregation experiments, yuzu and its component, hesperidin and
narigin were administered with oral gavage at a volume of 0.5 ml/kg per mouse or rat 2 h
before experiments. Control animals received the same volume of saline or 70% PEG,
respectively. All agents were prepared just before use.
PART II: For LV remodeling experiments, all experimental procedures conformed to the
Guide for the Care and Use of Laboratory Animals published by the US National Institutes
of Health (NIH Publication No. 85-23, revised 1996), and the Committee on Animal
Research at Ajou Medical Center, Ajou University (Suwon, Republic of Korea), approved
the study. Male Sprague-Dawley rats (weight, 250–300 g) were anaesthetized with an
intraperitoneal injection of ketamine (100 mg/kg) and xylazine (10 mg/kg) before surgery.
The body temperature of the rats was maintained at 37 ± 0.5°C during surgery by using a
thermostatically controlled warming plate as described previously (Kim et al., 2010) In the
loss-of-function study, ischaemia-induced myocardial injury was induced by ligating the
LAD artery as described previously (Kim et al., 2010;
Yamazaki
et al. 2010).Sham-operated control group (sham) underwent the same surgical procedures except that the suture
placed under the left anterior descending was not tied. At 4 weeks after LAD occlusion, rats
were euthanized by CO2 inhalation for heart isolation. LV was used for staining experiments.
7
blot analysis. Peri-infarct zone was defined as the area within 2 mm of the visible edge of
infarction (Kido et al, 2005).
The rats were randomly distributed into experimental pre-treatment groups with similar
body weights. Yuzu (100 mg/kg/day, n=20), hesperidin (30 mg/kg/day, n=20), vehicle (PEG
0.3 ml/day, n=23) or sham (PEG 0.3 ml/day, n=20) was administered by oral gavage once
daily beginning 7 days before LAD occlusion and continuously administered until the time
of the terminal study, using oral gavage around 10:00 A.M. every morning (for total 5 weeks,
Fig. 7.).
3. Animals
Sprague-Dawley (SD) rats and ICR mice were purchased from the Samtako Laboratory
Animal Center (Republic of Korea), and housed in a conventional animal facility with free
access to food and water in a temperature and relative humidity monitored and controlled
environment under artificial lighting (12 h of light per day). Animals were allowed to
acclimatize for at least 7 days before experiments. All animals related study protocols were
conducted in accordance with the guidelines for the Care and Use of Laboratory Animals
published by the US National Institute of Health (NIH Publication No. 85-23, revised 1996),
and were approved by the Committee on Animal Research at Ajou Medical Center, Ajou
University.
4. Preparation of platelets
8
Briefly, Sprague–Dawley (SD) rats, weighing 200–250 g, were lightly anesthetized with
ethyl ether and 8–10 ml of blood was collected from abdominal aorta into sodium citrate
(3.8%, 1:9 v/v) containing tubes. After centrifugation at 150g for 10 min at room temperature,
supernatants (PRP) were used for the aggregation study. PRP was centrifuged at 1200g for
10 min at room temperature and supernatant was obtained as platelet poor plasma (PPP)
which was used to adjust PRP. All experiments were conducted at least four times.
5. In vitro platelet aggregation study
Platelet aggregation studies were performed under the experimental setting described by
previous studies using the turbidimetric method (Seo et al., 2011). Briefly, PRP was
stimulated with different aggregating agents at the following final concentrations; collagen 2
μg/mL, thrombin 0.4 U/mL, AA 100 μM, or ADP 10 μM. Platelet aggregation was recorded 5 min after platelet stimulation. Aggregations were measured by a Lumi-aggregometer
(Chrono-Log Co., Harvertown, PA, USA) connected to computer and expressed as percent
changes in light transmission, taking the value of a blank sample (buffer without platelets) to
be 100%. For in vitro studies, PRP was preincubated with different concentrations of the four
anthraquinone derivatives for 5 min in the cuvette of an aggregometer before being
stimulated with the aggregating agents described above.
6. Determination of cytotoxicity
The cytotoxic effects of samples on platelets were determined by measuring lactate
9
PRP was incubated at 37 °C for 5 min with vehicle or samples, and centrifuged at room
temperature for 1 min at 10,000 g. Aliquots of supernatant (25 μL) were then placed into a
96-well plate and mixed with 100 μL of NADH solution (0.03% β-NADH in phosphate
buffer) and 25 μL of pyruvate solution (22.7 mM pyruvic acid in phosphate buffer) at room temperature. Reductions in absorbance at 340 nm due to the conversion of NADH to NAD+
were used as measures of LDH activity. LDH leakages were expressed as percentages of
total enzyme activity measured in platelets completely lysed with 0.2% Triton X-100.
7. Ex vivo platelet aggregation study
Two hours after the oral administration of yuzu (100 mg/kg), hesperidin (10 mg/kg), or
aspirin (50 mg/kg), rat blood samples were collected and the platelet aggregation
experiments were performed as described above.
8. In vivo mice tail bleeding times
Bleeding times were determined as previously described (Cho et al, 2008). Male ICR
mice weighing 35–40 g were used in this experiment. Mice were fasted overnight
beforehand. Two hours after the oral administration of yuzu (3, 10, 30 mg/kg), hesperidin (1,
3, 10 mg/kg) or aspirin (50 mg/kg), mice were anesthetized with sodium pentobarbital (75
mg/kg, i.p.). Mice were then placed individually on a hotplate to control body temperature at
37 °C and tails were transected 3 mm from their tips with a razor blade and then immersed
in a 15 ml clear conical tube containing normal saline prewarmed to 37 °C. Times to blood
10
stopped for 15 s. Bleeding times exceeding 15 min were recorded as 15 min for the purposes
of statistical analysis.
9. Measurement of thromboxane A2 (TXA2) formation
The formation of TXA2 in platelets was measured by determining TXB2 using a TXB2
EIA kit (Cayman Chemical Co., Ann Arbor, MI, USA), because TXA2 is unstable and
quickly converted to TXB2. A suspension of rat PRP was preincubated for 5 min in the
presence or absence of DMSO, yuzu, hesperidin, naringin, or aspirin before adding collagen
(2 mg/ml) or thrombin (0.4 U/ml). After incubation at 37 °C for 5 min with collagen or
thrombin, EDTA (10 mM) was added to stop TXA2 formation. TXB2 in supernatant was
obtained by centrifuging at 12,000g for 1 min, and the amount of TXB2 in medium was
determined by using the TXB2 EIA kit, according to the procedure described by the
manufacturer (Cayman Chemical Co.).
10. Echocardiography
The rats were subjected to transthoracic echocardiography. In brief, the rats were
anaesthetized with an intraperitoneal injection of ketamine (100 mg/kg) and xylazine (10
mg/kg) and were examined with non-invasive echocardiography (echocardiograph IE33
ultrasound, S12-4 probe, Philips). Ventricular remodeling in the vehicle-, yuzu-, or
hesperidin-treated groups was assessed weekly with serial echocardiography (beginning 0
week prior to LAD occlusion until 4 weeks after LAD occlusion). Cardiac ventricular
11
of 5 consecutive cardiac cycles of each animal is reported.
11. Histological analysis
LV tissue was fixed with 4% paraformaldehyde for 24 hours, dehydrated with
increasing concentrations of ethanol, and then embedded in paraffin. LV sections (5 μm)
were stained with hematoxylin and eosin (H&E, Sigma-Aldrich) or Masson trichrome
(polysciences) as previously described (Akhlaghi and Bandy, 2010; Lal et al, 2012). Images
were captured by utilizing a Zeiss Axioplot Vision-series microscope and software (Carl
Zeiss, Oberkochen, Germany) and were quantified using the NIH Image J analysis program
(NIH, Bethesda, MD, USA).
12. Immunohistochemistry
Immunostaining was performed using a streptavidin–biotin-immunoperoxidase
complex method with 5 μm thick sections, which had been deparaffinized and heated in 0.01 M citrate buffer solution (pH ¼ 6.0) for 15 min for antigen retrieval. Rabbit polyclonal
antibody against, myeloperoxidase (MPO), or cardiac tronponin I (cTnI) purchased from
Abcam (Cambridge, UK) was used. The slides were examined using a light microscope
(Olympus CX21, Japan) and was evaluated with reference to the optical density of the stain
by using a computer-assisted image analysis system, ImageJ1.45F (NIH, USA).
13. Gelatin zymography
Gelatin zymography was performed utilizing the Novex In-gel Zymography System
12
were homogenized in 50 mM Tris-HCl (pH 7.5) containing 150 mM NaCl and 5 mM CaCl2.
After centrifugation, the supernatants were harvested and 25 µg of protein was mixed with
Tris-glycine sodium dodecyl sulfate (SDS) sample buffer. The samples were run on a Novex
10% Zymogram Gelatin Gel, followed by incubation with Zymogram Renaturing Buffer and
subsequent incubation with Zymogram Developing Buffer. After an overnight reaction, the
gel was stained with Simply Blue Safe Stain (Invitrogen).
14. Western blot analysis
Heart tissues were homogenized in a buffer containing 50 mmol/L Tris-HCl pH 7.4,
1% NP-40, 150 mmol/L NaCl, 0.25% Na-deoxycholate, 2 mmol/L EDTA, 1 mmol/L NaF, 1
mmol/L Na3VO4, 1 mmol/L PMSF, 10 μg/mL aprotinin, and 10 μmol/L leupeptin.
Homogenates were centrifuged at 14,681 g for 15 min and the supernatants were collected as
previously described (Kim et al, 2010). Equal amounts of protein were then separated by
SDS-polyacrylamide gel electrophoresis (PAGE) and reacted with antibodies specific for
caspase-3 (Cell Signaling, Danvers, MA, USA), α-SMA (Abcam) and α-tubulin
(Sigma-Aldrich). After probing with an HRP-conjugated secondary antibody, the proteins were
visualized using LAS 1000 (Fuji Photo Film, Tokyo, Japan). Densitometric analyses were
performed using Quantity One software,ImageJ1.45F (NIH, USA).
15. Terminal dUTP nick end-labeling (TUNEL) staining
In situ labeling of fragmented DNA was performed using the Apop Taq Plus Kit
13
digoxigenin nucleotide and reacted with peroxidase-conjugated anti-digoxigenin antibody
and 3,3′-diaminobenzidine as previously described (Kim et al, 2010). The percent cell death
was calculated by expressing the number of TUNEL-positive cells as a percentage of total
cell counts.
16. Statistical analysis
All data are expressed as means ± SDs. The results were analyzed using 2-way ANOVA
and the differences between groups were compared by using the Student’s t-test. A p value of <0.05 was considered statistically significant. All experiments were repeated at least 4 times.
14
III. RESULTS
A. PART I. Anti-platelet effects of yuzu extract and its component
1. Effects of yuzu on platelet aggregations in vitro
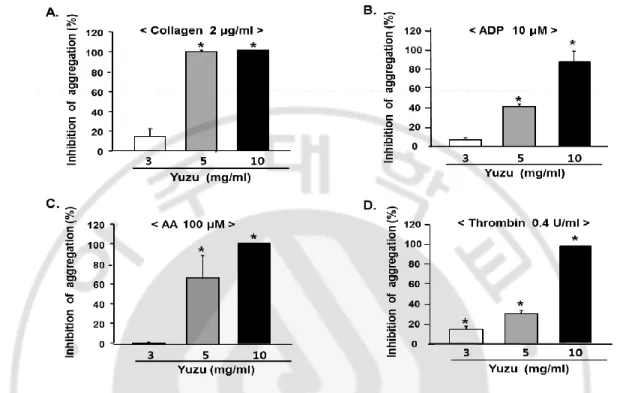
Yuzu (3–10 mg/ml) was found to inhibit collagen-, ADP-, AA- and thrombin-induced
aggregations in a dose-dependent manner (Fig. 1). Yuzu at 5 mg/ml inhibited collagen-,
ADP-, AA- and thrombin-induced platelet aggregations by 98.4 ± 1.6%, 41.5 ± 2.7%, 65.3 ±
21.7% and 31.1 ± 3.4% compared to vehicle, respectively. The IC50 values (half inhibitory
15
Fig. 1. Dose-dependent inhibitory effect of yuzu on in vitro platelet aggregation. Platelet
were preincubated for 5 min with various concentrations of yuzu at 37 °C before being aggregated with collagen 2 µg/ml (A), ADP 10 µM (B), or arachidonic acid (AA) 100µM (C) and thrombin 0.4 U/ml (D). Data were expressed as means ± SEMs (n = 4–5). * P < 0.05
16
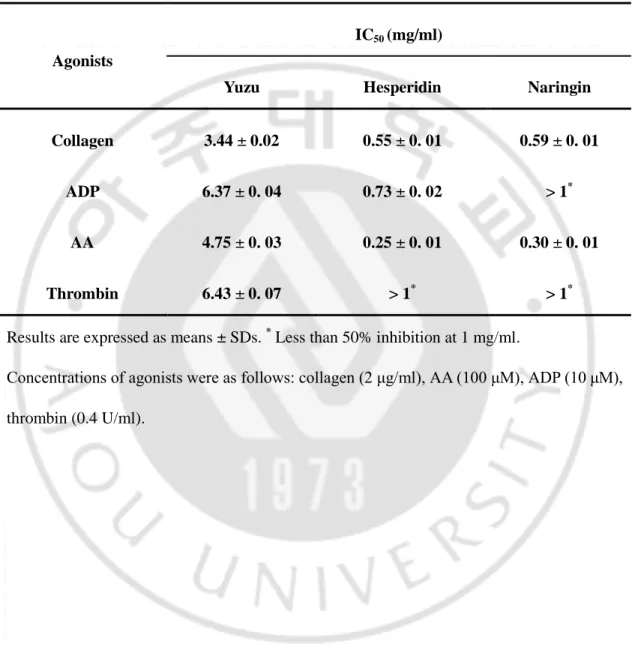
Table 1. IC50 of yuzu, hesperidin, and naringin for platelet aggregation.
Results are expressed as means ± SDs. * Less than 50% inhibition at 1 mg/ml.
Concentrations of agonists were as follows: collagen (2 μg/ml), AA (100 μM), ADP (10 μM), thrombin (0.4 U/ml).
Agonists
IC50 (mg/ml)
Yuzu Hesperidin Naringin
Collagen 3.44 ± 0.02 0.55 ± 0. 01 0.59 ± 0. 01
ADP 6.37 ± 0. 04 0.73 ± 0. 02 > 1*
AA 4.75 ± 0. 03 0.25 ± 0. 01 0.30 ± 0. 01
17
2. Contents of hesperidin and naringin in yuzu extracts
The high content of hesperidin and naringin in yuzu might be associated with significant
health benefits. The contents of hesperidin and naringin in yuzu extract are shown in Table 2.
Mean total amounts of hesperidin and naringin present in yuzu extract (mg/100 g of F.wt
yuzu) were 413 ± 15.06 and 1191.0 ± 17.19 mg/100 g fresh weight, respectively. HPLC
18
Table 2. Contents of hesperidin and naringin in extract of yuzu.
Hesperidin Naringin
Contents
(mg/100 g of F.wt yuzu )
413.79 ± 15.06 1191.0 ± 17.19
19
20
3. Effects of hesperidin and naringin on platelet aggregations in vitro
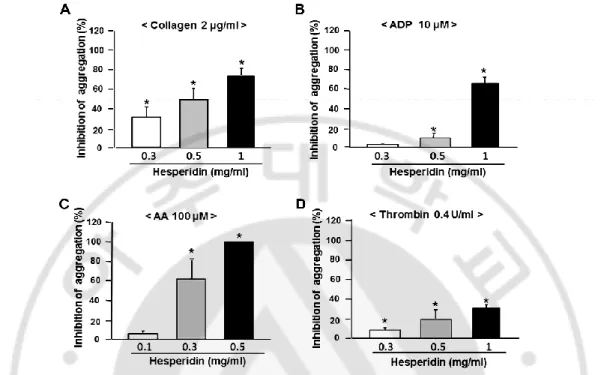
Hesperidin (0.1–1.0 mg/ml) was found to inhibit collagen-, ADP-, AA-, and
thrombin-induced aggregations in a dose-dependent manner (Fig. 3). As shown in Table 1, IC50 values
of hesperidin for collagen-, ADP-, AA- and thrombin-induced platelet aggregations were
0.55 ± 0.01, 0.73 ± 0.02, 0.25 ± 0.01, and >1 mg/ml, respectively.
Naringin at 1 mg/ml inhibited collagen-, AA-, and thrombin-induced platelet
aggregation by 82.4 ± 8.9%, 75.6 ± 24.4%, and 45.7 ± 8.9%, respectively (Fig. 4). However,
naringin showed little effect on ADP-induced platelet aggregation. IC50 values of naringin for
collagen-, ADP-, and AA- and thrombin-induced platelet aggregations were 0.59 ± 0.01, >1,
21
Fig. 3. Dose-dependent inhibitory effect of hesperidin on in vitro platelet aggregation.
Platelets were preincubated for 5 min with various concentrations of hesperidin at 37 °C before being aggregated with collagen 2 µg/ml (A), ADP 10 µM (B), or arachidonic acid (AA) 100 µM (C) and thrombin 0.4 U/ml (D). Data were expressed as means ± SEMs (n =
22
Fig. 4. Dose-dependent inhibitory effect of naringin on in vitro platelet aggregation.
Platelets were preincubated for 5 min with various concentrations of naringin at 37 °C before being aggregated with collagen 2 µg/ml (A), ADP 10 µM (B), or arachidonic acid (AA) 100 µM (C) and thrombin 0.4 U/ml (D). Data were expressed as means ± SEMs (n =
23
4. Cytotoxic effects of yuzu, hesperidin, and naringin on platelets
To examine the cytotoxicities of yuzu, hesperidin, and naringin, LDH release from
platelets was measured. LDH released from platelets treated with DMSO, yuzu (10 mg/ml),
hesperidin (1 mg/ml), or naringin (1 mg/ml) were not significantly changed when compared
to non-treated controls, while that from platelets treated with digitonin (50 µM), used as a
positive control, were significantly increased. These results suggest that the anti-platelet
effects of yuzu, hesperidin, and naringin are unlikely to be associated with cytotoxicity
24
Table 3. Effects of yuzu, hesperidin, and naringin on LDH release by platelets.
LDH release was measured after incubating rat platelet-rich plasma (2×108 cells/ml) with
vehicle or samples for 5 min. Data represent means ± SEMs (n=4~6). * P<0.05 vs.
control(no- treatment)s.
Control DMSO Yuzu Hesperidin Naringin Digitonin LDH
release
(% of max)
25
5. Effects of yuzu, hesperidin, and naringin on TXB
2formation
To investigate the possible mechanism responsible for the antiplatelet effects of yuzu,
hesperidin, and naringin, their effects on TXB2 formation were evaluated. As shown in Fig. 5,
pretreatment of platelets with 1 mg/ml of hesperidin significantly inhibited collagen-induced
TXB2 formation, but had little effect on thrombin induced TXB2 formation. In the case of
yuzu (10 mg/ml) and naringin (1 mg/ml) treatment, collagen- and thrombin-induced TXB2
formation was inhibited. Aspirin, a positive control, completely blocked TXB2 formation at a
26
Fig. 5. Effect of yuzu, hesperidin, and naringin on collagen- and thrombin-induced
TXB2 formation in rat platelets. After preincubating platelet suspension with DMSO, yuzu (10 mg/ml) , hesperidin (1 mg/ml), naringin (1 mg/ml) or aspirin for 5 min, 2 µg/ml of
collagen or 0.4 U/ml of thrombin was added. After incubation at 37 °C for 5 min with collagen or thrombin, EDTA (10 mM) was added to stop TXB2 formation, and supernatant
was obtained by centrifugation at 12,000g for 1 min (A, B). TXB2 concentrations were
determined using an EIA kit. Data are expressed as means ± SEMs (n = 3–5). *P < 0.05 vs. controls. # P < 0.05 vs. vehicle.
27
6. Effects of yuzu and hesperidin on platelet aggregation ex vivo
To determine the concentration required for the inhibitory effects of yuzu and hesperidin
on ex vivo platelet aggregation, the effects of various concentrations of yuzu (10, 30, 100
mg/kg) and hesperidin (1, 3, 10 mg/kg) were evaluated in a preliminary experiment (data not
shown); 100 mg/kg and 10 mg/kg were chosen as optimum concentrations for yuzu and
hesperidin, respectively, for the ex vivo platelet aggregation experiments. Hesperidin has
more inhibitory effects on 4 agonist-induced platelet aggregation than naringin, and thereby
it was focused on hesperidin in later experiments.
As shown in Table 4, yuzu administered at 100 mg/kg (p.o.) 2 h before blood collection
significantly inhibited collagen-, ADP-, AA-, and thrombin-induced platelet aggregations, by
32.0 ± 8.6%, 17.3 ± 2.5%, 37.5 ± 10.6%, 18.4 ± 4.8% compared to control, respectively.
Similarly, hesperidin administered at 10 mg/kg (p.o.) significantly inhibited collagen-, ADP-,
AA-, and thrombin-induced platelet aggregations by 23.6 ± 6.5%, 19.9 ± 4.2%, 41.0 ± 13.8%,
13.5 ± 2.1% compared to control, respectively. Aspirin (50 mg/kg), a reference anti-platelet
drug, also significantly inhibited collagen-, AA-, ADP-, and thrombin-induced platelet
28
Table 4. Inhibitory effects (%) of orally administered yuzu and hesperidin on ex vivo
platelet aggregation.
Inhibition of platelet aggregation (% of control)
Agonists Yuzu (100 mg/kg, P.O.) Hesperidin (10 mg/kg, P.O.) Aspirin (50 mg/kg, P.O.) Collagen 32.0 ± 8.6%* 23.6 ± 6.5%* 32.5 ± 10.4%* AA 37.5 ± 10.6%* 41.0 ± 13.8%* 99.6 ± 0.3%* ADP 17.3 ± 2.5%* 19.9 ± 4.2%* 21.6 ± 6.6%* Thrombin 18.4 ± 4.8%* 13.5 ± 2.1%* 11.2 ± 3.0%*
Concentrations of agonists were as follows: collagen (2 μg/ml), AA (100 μM), ADP (10 μM),
and thrombin (0.4 U/ml). Data are expressed as means ± SEMs (n = 5~7). *P<0.05 vs.
29
7. Effects of yuzu and hesperidin on tail bleeding times in mice
Because the mouse tail bleeding time assay is an easily accessed, reliable measure of
platelet function, it has been widely used to evaluate anti-platelet effects (Gadi et al., 2009).
Therefore, it was performed bleeding time assays to examine the effects of yuzu and
hesperidin on platelet function in vivo. As shown in Fig. 6, the mean bleeding time of
untreated control mice were 92.8 ± 13.4 sec, and this was significantly prolonged by 30
mg/kg (p.o.) of yuzu or 10 mg/kg (p.o.) of hesperidin to 442.3 ± 95.6 sec and 422.2 ± 94.9
sec, respectively. Aspirin at 50 mg/kg (p.o.) also significantly prolonged bleeding time (to
30
Fig. 6. Effects of yuzu and hesperidin on mouse tail bleeding times. Yuzu (3, 10, or 30
mg/kg), hesperidin (1, 3, or 10 mg/kg) or aspirin (50 mg/kg) were orally administered to mice 2 h before the experiments. Data were expressed as means ± SEMs (n = 7–9). *P < 0.05 vs. control (no-treatment).
31
B. PART II. Preventive effect of yuzu and hesperidin on LV remodeling and
dysfunction in rat permanent LAD occlusion model
1. Yuzu and hesperidin prevented LV remodeling and functional
deterioration following MI
In this study, it was evaluated the effects of yuzu and hesperidin on LV remodeling in a
rat model of permanent LAD occlusion (PO). As shown in Fig. 7, cardiac function
measurement time
was examined by non-invasive echocardiography.
Ventricular
remodeling in the vehicle-, yuzu-, or hesperidin-treated groups was assessed weekly
with serial M-mode echocardiogram (Fig. 8).
The vehicle-treated group showed asignificantly greater increase in end-diastolic and end-systolic dimensions (LVIDd & LVIDs)
compared to sham group. This was associated with marked LV dysfunction as reflected by
reduced LV ejection fraction (EF) (Fig. 9A-C). LV function was preserved more markedly in
the yuzu- or hesperidin-treated groups up to the termination of the study. However, yuzu and
hesperidin treatments for 4 week after LAD occlusion did not significantly attenuated
cardiac hypertrophy, as measured by the heart weight to body weight (HW/BW) ratio. (Fig.
32
33
Fig. 8. Effect of pre-treatment with yuzu (100 mg/kg/day) or hesperidin (HSP, 30
34
Fig. 9. Effect of pre-treatment with yuzu (100 mg/kg/day) or hesperidin (HSP, 30
mg/kg/day) on LV dysfunction and LV remodeling Echocardiographic measurement of
left ventricular (LV) internal dimensions at both systole and diastole (LVIDs and LVIDd,
respectively), LV ejection fraction, andheart weight (HW) to body weight (BW) ratio. Data
are expressed as mean ± SEM; *p<0.05 vs. sham; #p<0.05 vs. vehicle (permanent LAD
35
2. Yuzu and hesperidin prevented myocardial fibrosis during LV remodeling after
chronic MI
To determine whether in vivo findings have pathological relevance, it was evaluated the
effects of yuzu and hesperidin on cardiac fibrosis in a rat MI model of permanent LAD
occlusion. The rats were treated with yuzu, hesperidin, or vehicle 1 week before LAD
occlusion. As observed upon Masson’s trichrome staining, increased interstitial fibrosis was observed in the rats treated with the vehicle after 4 weeks of LAD occlusion, which was
significantly reduced in the group that was pre-treated with yuzu or hesperidin (Fig. 10A and
B). Western blot analysis revealed upregulation of α-SMA expression in hearts from the vehicle group, which was reversed to control levels by pre-treatment with yuzu or hesperidin
36
Fig. 10. Effect of pre-treatment with yuzu (100 mg/kg/day) or hesperidin (HSP, 30 g/kg/day) on cardiac fibrosis. (A) Representative Masson’s trichrome sections of the left
ventricle. Scale bar, 100 μm. n=6-7. (B) Quantitative analysis of fibrosis. Data are expressed as mean ± SEM; *p<0.05 vs. sham; #p<0.05 vs. vehicle. n=6-7. PO, permanent LAD occlusion.
37
Fig. 11. Effect of pre-treatment with yuzu (100 mg/kg/day) or hesperidin (HSP, 30
g/kg/day) on myofibroblast. Expression of myocardial α-smooth muscle actin (α-SMA)
protein. Representative image (upper part) and quantitative analysis (bottom part). PO, permanent LAD occlusion. Data are expressed as mean ± SEM; *p<0.05 vs. sham; #p<0.05
38
3. Yuzu and hesperidin reduced the inflammatory reaction and MPO
expression during LV remodeling after chronic MI
To histologically confirm that myocardial injury reflected inflammation, it was
performed H&E staining and
myeloperoxidase
(MPO) immunohistochemical analysis todetect neutrophil activity. H&E staining showed that the strongest inflammatory reactions
were observed in the vehicle group. Examination of heart sections of the yuzu- and
hesperidin-treated groups showed nearly normal cardiac cells with a well-preserved
cytoplasm and prominent nucleolus (Fig. 12). The number of MPO-expressing cells was
significantly higher in the vehicle groups than in the yuzu- and hesperidin-treated groups
39
Fig. 12. Effect of pre-treatment with yuzu (100 mg/kg/day) or hesperidin (HSP, 30
mg/kg/day) on inflammatory reaction Representative H & E staining sections of the left
ventricle (LV). PO, permanent LAD occlusion, H & E, hematoxylin & eosin Scale bar, 100
40
Fig. 13. Effect of pre-treatment with yuzu (100 mg/kg/day) or hesperidin (HSP, 30 mg/kg/day) on MPO expression (A) Representative H & E staining sections of the left
ventricle (LV). Scale bar, 100 μm. n=6-7. (B) Representative examples of neutrophil activity expressed as myeloperoxidase (MPO) antibody activity (arrow) in the LV. Scale bar, 100 μm. n=6-7. (B) Quantitative analysis of MPO positive cells. Data are expressed as mean ± SEM;
41
4. Yuzu and hesperidin block myocardial apoptosis through inhibition of
caspase-3 activation during LV remodeling after chronic MI
It was next investigated whether adverse LV remodeling contributed to increased cardiac
cell death in the rats with LV remodeling after MI. Apoptosis was evaluated using the
TUNEL assay. After 4 weeks of LAD occlusion, the number of TUNEL-positive myocytes
was significantly higher in the vehicle groups than in the yuzu- and hesperidin-treated groups
(Fig. 14A-B). Thus, yuzu and hesperidin attenuated MI-induced myocardial apoptosis. Next,
we compared the caspase-3 activity of the yuzu- and hesperidin-treated groups with the
vehicle group to determine whether caspase-3 is involved in myocardial apoptosis.
Treatment with yuzu or hesperidin significantly reduced caspase-3 activity to levels similar
42
Fig. 14. Effect of pre-treatment with yuzu (100 mg/kg/day) or hesperidin (HSP, 30 mg/kg/day) on cardiomyocyte apoptosis. (A) Representative TUNEL staining and (B)
quantitative analysis of TUNEL positive cells. Data are expressed as mean ± SEM; *p<0.05
43
Fig. 15. Effect of pre-treatment with yuzu (100 mg/kg/day) or hesperidin (HSP, 30
mg/kg/day) on caspase-3 activity. The activity of myocardial caspase-3 protein.
Representative image (upper part) and quantitative analysis (bottom part). n=7-8. PO, permanent LAD occlusion
44
5. Yuzu and hesperidin block LV wall thinning through inhibition of MMP-2
activation during LV remodeling after chronic MI
LAD occlusion for 4 weeks induced prominent cardiomyocyte loss in the LV mass, as demonstrated by decreased cardiac troponin I (cTnI) staining. Compared to vehicle treatment,
yuzu or hesperidin treatment significantly inhibited cardiomyocyte loss and expression of cTnI (Fig. 16). To gain a better understanding of the mechanisms leading to the prevention of
LV wall thinning in yuzu- and hesperidin-treated groups, it was examined matrix metalloproteinases-2 (MMP-2) activity and resident cardiomyocytes in the LV myocardium.
MMP-2 activity increased in the left ventricles of rats subjected to LAD occlusion, whereas treatment with yuzu significantly reduced MMP-2 activity to levels similar to those observed in sham-operated animals (Fig. 17). These effects were associated with a significant
45
Fig. 16. Effect of pre-treatment with yuzu (100 mg/kg/day) or hesperidin (HSP, 30 mg/kg/day) on cardiomyocyte preservation. Representative cardiac Troponin I (cTnI, red
florescence, blue, neuclei) staining. Scale bar, 100 μm. n = 6-7. PO, permanent LAD
46
Fig. 17. Effect of pre-treatment with yuzu (100 mg/kg/day) or hesperidin (HSP, 30
mg/kg/day) on MMP-2 activation. The activity of MMP-2. Representative image by
zymography (upper part) and quantitative analysis (bottom part). n=7-8. PO, permanent LAD occlusion
47
IV. DISCUSSION
A. PART I. Anti-platelet effects of yuzu extract and its component
Under pathologic conditions, vascular injury results in the rapid generation of thrombin
at sites of injury and the exposure of extracellular matrix, such as, collagen in vessel walls,
thereby triggering platelet aggregation and thrombus formation via collagen and thrombin
dependent mechanisms (Nieswandt et al, 2005). Collagen- and thrombin-induced platelet
aggregations are ADP and TXA2-dependent, because both agonists can induce the release of
ADP and TXA2 (Farndale et al, 2004). There are, however, differences between the signaling
pathways of collagen- and thrombin-induced platelet activation. Platelet responses to
collagen are mediated via glycoprotein VI (GPVI) and integrin 21 (Farndale et al, 2004),
while those to thrombin are largely mediated through G-protein coupled protease-activated
receptors (PARs) (Brass, 2003). Collagen induces platelet activation through a tyrosine
kinase-based signaling pathway that involves Syk kinase and PLC-2, which results in cytosolic Ca2+ increase, shape change, and the releases of ADP and TXA2. Thrombin bound
to PAR-1 on platelet surfaces causes the activation of the Gq/PLC-2pathways (Brass, 2003), which leads to generation of IP3, the mobilization of intracellular Ca
2+
, an increase in Src
48
upon exposure to activating agonists, such as collagen and thrombin, platelets liberate AA,
which is then converted by cyclooxygenase and TXA2 synthase into TXA2 (Jennings, 2009).
ADP secreted from dense granules in activated platelets induces platelet aggregation via the
G protein-coupled purinergic receptors, P2Y1 and P2Y12, in an autocrine manner and this
promotes stable platelet aggregation. The current view of the relationship between these two
platelet ADP receptors is that P2Y1 initiates aggregation and that this is reinforced by P2Y12
(Davì et al, 2007). P2Y1 coupled to Gq regulates Ca 2+
-dependent signaling events, and
P2Y12 is Gi-linked and activates integrin IIb3 by a mechanism involving the inhibition of
cAMP production by adenyl cyclase (Hardy et al, 2004). Taken together, platelets are
activated by multiple physiological agonists that interact with their specific receptors and
trigger different signaling pathways, and thus, antiplatelet agents that act only at one site are
likely to be limited in terms of preventing the formation of pathogenic thrombi. Indeed, dual
anti-platelet therapy with aspirin plus clopidogrel, a P2Y12-receptor antagonist, has been
shown to be more effective at reducing ischemic events in patients with atherothrombotic
cardiovascular diseases than aspirin alone (Chen et al, 2005). Furthermore, previous
observations support the notion that a combination of antiplatelet drugs and preparations of
herbal food supplements may be beneficial in some clinical states and that a synergism exists
between flavonoids and aspirin in terms of their inhibitory effects on platelet function.
(Navarro-Núñez et al, 2008). Taken together, antiplatelet agents that act at multiple sites are
49
has significant inhibitory effects on platelet aggregations induced by multiple agonists in in
vitro PRP and ex vivo models. However, it is important to note that after oral ingestion, the
bioavailability of active compounds present in the yuzu and hesperidin depended on their
gastrointestinal metabolization and absorption degree. Nevertheless, this result indicated that
yuzu may have therapeutic benefit in terms of preventing atherothrombotic disease due to its
potential to inhibit multiple sites of platelet aggregation.
Consistent with previous reports, the present study shows that hesperidin and naringin
are major flavonoid components in yuzu. Regarding their effects on platelet aggregation,
hesperidin and naringin showed limited inhibitory effect on thrombin-induced aggregation,
and naringin, in particular, was found to have no effect on ADP-induced aggregation,
whereas yuzu showed multiple inhibitory effects on collagen-, ADP, AA, and
thrombin-induced platelet effects. These results suggest that the anti-platelet activity of yuzu may be
attributable to the combined effects of its components, including those of hesperidin and
naringin.
Regarding platelet activation in response to various stimuli, TXA2 is an important
mediator of platelet activation and aggregation (Jennings, 2009). Accordingly, agents that
inhibit TXA2 formation are expected to play important roles in the pathogenesis of
platelet-linked cardiovascular disease. In this study, it was examined the effects of yuzu, hesperidin,
and naringin on TXA2 formation induced by collagen and thrombin. It was found that yuzu,
50
However, when platelets were stimulated by thrombin, yuzu and naringin (but not
hesperidin) significantly inhibited TXB2 formation. In terms of collagen-induced platelet
aggregation, it has been reported that hesperetin inhibits platelet aggregation by inhibiting
PLC-γ2 phosphorylation (Jin et al, 2007). However, in case of thrombin-mediated platelet
aggregation, both Gαq and PLC-β2 play major roles in responses to activation by PAR1 or PAR4, and that PLC-β2 is required for the sustained Ca2+ increase that occurs after thrombin
activation (Vaidyula and Rao, 2003). Based on these results, it is likely that hesperidin has a
limited effect on the PLC-β2 signaling pathway, and thus, a limited inhibitory effect on
thrombin-induced platelet aggregation. The inhibitions of cyclooxygenase and TXA2
synthase are well known inhibitory mechanisms of TXA2 formation. Therefore, further study
is required to elucidate whether the anti-platelet effects of yuzu and hesperidin are associated
with the inhibitions of cyclooxygenase and/or TXA2 synthase.
In this study, yuzu and hesperidin increased bleeding time more than aspirin in in vivo
study. Actually, there are many reports showing that the administration of anti-platelet drug
causes an increase in the bleeding time as a side effect (Kim et al, 2004). However, even now,
no drug is used by a greater number of people worldwide than aspirin. Furthermore, its
combination therapy, such as aspirin-clopidogrel-cilostazol, provides incremental benefit,
becausemultiple pathways activate platelets (Goto, 2005; Han et al, 2009; Jennings, 2009).
In case of aspirin, given the chronicity of aspirin therapy used to treat CVD, optimal dosing
51
Sweeny, 2011). Therefore, the use of aspirin for either primary or secondary prevention of
coronary artery disease is largely a risk-benefit calculation. Likewise, these results suggest
that optimal intake of yuzu to minimize its adverse effect should be emphasized and that
development of a novel combination approach with yuzu remains to be investigated.
B. PART II. Preventive effect of yuzu and hesperidin on LV remodeling and
dysfunction in rat permanent LAD occlusion model
Herein, this study provide the first evidence for the cardioprotective role of yuzu and its
major component, hesperidin in rat chronic MI model induced by permanent LAD occlusion,
by demonstrating that yuzu and hesperidin prevented heart from MI-induced dysfunction, LV
fibrosis, inflammatory reaction and cardiomyocyte apoptosis.
Chronic MI results in complex architectural alterations such as dilatation of the LV and
infarct thinning, which is called LV remodeling, in both infarct and non-infarct region.
Patients exhibiting extensive LV remodeling after MI are more likely to experience
complications such as HF and myocardial rupture, leading to an elevated risk of mortality
(Creemers et al, 2001). Although modern cardiology has made substantial advances in the
diagnosis and management of MI, it is necessary to design therapeutic strategies to attenuate
LV remodeling after MI by modulation of the molecular and cellular factors involved in the
52
After chronic MI, LV fibrosis contributes to adverse structural remodeling, leading to
impaired contractile properties of the LV as well as deteriorated electrical conduction system
(van den Borne et al, 2010). LV fibrosis occurs as a result of the imbalance between
enhanced synthesis reduced degradation of collagen (González et al, 2011). As a critical
step in response to myocardial injury, fibroblasts are activated into α-SMA-positive myofibroblasts which can generate extracellular matrix proteins such as type I collagen.
Therefore, the degree of fibroblast activation is a significant predictor of HF progression in
both experimental animal models and in human patients (Pchejetski et al, 2012). The
present study have demonstrated that yuzu and hesperidin exert anti-fibrotic remodeling
effects via the inhibition of excess collagen deposition and conversion of fibroblasts into
α-SMA-positive myofibroblasts during LV remodeling process following permanent LAD
occlusion. These results suggest that yuzu and hesperidin represent novel preventive natural
products against LV remodeling and cardiac dysfunction induced by chronic MI.
Regarding cardiac structure remodeling process after chronic MI, recent studies have
identified the importance of several inflammatory mediators that are released during this
process, such as neutrophils and various cytokines, and inflammatory cells are attracted to
the myocardial injury site (Paulus, 2000; Diwan et al, 2003; Campian et al, 2010). MPO is a
well-known enzyme that is released by activated neutrophils and has powerful pro-oxidative
and pro-inflammatory properties. Recent studies have suggested that regional MPO activity
53
2010; Tang et al, 2011). Subsequent studies have provided quantitative support for this
observation by showing significant elevations in the systemic levels of MPO in a wide
spectrum of CVD scenarios, with chronic MI and HF being the most frequently studied
(Anatoliotakis et al, 2013). Accordingly, pre-clinical studies in experimental models suggest
a possible therapeutic role of MPO inhibition in HF. In this study, it was found that yuzu and
hesperidin have anti-inflammatory properties and participate in the control of the
inflammatory response through the inhibition of MPO expression during chronic MI.
Experimental models and clinical studies have shown that loss of functional
cardiomyocytes contributes to structural changes that underlie progressive LV remodeling
during chronic MI (González et al, 2011; Konstantinidis et al, 2012). Although the
significance of apoptosis in LV remodeling still remains debatable, cardiomyocyte apoptosis
leads to the loss of cardiomyocyte mass and the reduction of myocardial contractile function
(Dom, 2009). Therefore, elimination of pro-apoptotic signals may prevent the progression of
LV remodeling during chronic MI. Since the main apoptotic death pathways converge on
caspases, the most efficient approach for interrupting cardiomyocyte apoptosis might be the
targeting of these enzymes. Many animal studies have confirmed that the inhibition of
caspases, such as caspase-3, mitigates LV dysfunction and enables survival during the
progression to end-stage HF. Although specific caspase inhibitors are being developed and a
few have shown promising results for clinical therapy by targeting cell death in LV
54
until very recently (Yang et al, 2013). The present study have demonstrated that yuzu and
hesperidin have anti-apoptotic properties and participate in the control of cell death by
inhibiting caspase-3 activity and expression.
Cardiomyocyte loss during ischemic damage can be replaced by non-contractile fibrotic
cells rather than by new cardiomyocytes (van Wijk et al, 2012). In humans, it is known that
extensive LV dilatation after chronic MI increases the risk of complications such as the HF,
aneurysm formation and cardiac rupture. In addition, the positive effects of MMP inhibition
on LV dilatation in animal models have led to the proposed use of MMP inhibitors as
potential therapies in patients at risk for the development of HF after MI (Creemers et al,
2001). MMP-2 is abundant and ubiquitously expressed in almost all of the cells that
comprise the heart. Activated MMP-2 degrades susceptible sarcomeric and cytoskeletal
proteins including troponin I (TnI), myosin light chain-1 (MLC-1), and α-actin, leading to
the acute contractile dysfunction observed in ischemia/reperfusion injury (Kandasamy et al,
2010). Interestingly, the present study have demonstrated that yuzu and hesperidin
effectively preserved cardiomyocyte mass during chronic MI, possibly through the inhibition
of MMP-2-induced wall thinning.
LV remodeling after chronic MI remains a major cause of morbidity and mortality
worldwide, leading to a dramatical increase of health care costs (Strauer et al, 2010).Despite
a number of pharmacological advances, mortality following MI remains still high. The
55
prevented LV remodelling and LV dysfunction following chronic MI, and suggested a
potential use of yuzu or hesperidin as a cardioprotective strategy.
V. CONCLUSION
In conclusion, yuzu and its major compounds hesperidin and naringin were found to
have significant anti-platelet activity, possibly via the inhibition of TXA2 formation. These
findings suggest that yuzu-based foods may be especially beneficial in the prevention of
platelet-associated atherothrombotic disease
In addition, the present study has demonstrated that pre-treatment of yuzu or hesperidin
significantly prevented LV remodeling following chronic MI in rat LAD occlusion model.
56
REFERENCES
1. Akhlaghi M, Bandy B: Dietary broccoli sprouts protect against myocardial oxidative
damage and cell death during ischemia-reperfusion. Plant Foods Hum Nutr. 65:
193-199, 2010
2. Anatoliotakis N, Deftereos S, Bouras G, Giannopoulos G, Tsounis D, Angelidis C,
Kaoukis A, Stefanadis C: Myeloperoxidase: expressing inflammation and oxidative
stress in cardiovascular disease. Curr Top Med Chem 13: 115-138, 2013
3. Benavente-García O, Castillo J : Update on uses and properties of citrus flavonoids:
new findings in anticancer, cardiovascular, and anti-inflammatory activity. J Agri.
Food Chem. 56: 6185-6205, 2008
4. Brass LF: Thrombin and platelet activation. Chest 124: 18S-25S, 2003
5. Campian ME, Hardziyenka M, de Bruin K, van Eck-Smit BL, de Bakker JM,
Verberne HJ, Tan HL: Early inflammatory response during the development of right
ventricular heart failure in a rat model. Eur J Heart Fail 12: 653-658, 2010
6. Cazenave JP, Ohlmann P, Cassel D, Eckly A, Hechler B, Gachet C : Preparation of
washed platelet suspensions from human and rodent blood. Methods Mol Biol 272:
13-28, 2004
Bennetau-57
Pelissero C, Morand C, Bérard AM: Naringin, the major grapefruit flavonoid,
specifically affects atherosclerosis development in diet-induced
hypercholesterolemia in mice. J Nutr Biochem J Nutr Biochem 23(5): 469-477, 2012
8. Chen ZM, Jiang LX, Chen YP, Xie JX, Pan HC, Peto R, Collins R, Liu LS,
COMMIT (ClOpidogrel and Metoprolol in Myocardial Infarction Trial)
collaborative group: Addition of clopidogrel to aspirin in 45,852 patients with acute
myocardial infarction: randomized placebo-controlled trial. Lancet 366: 1607-1621,
2005
9. Cho J, Furie BC, Coughlin SR, Furie B: Acritical role for extracellular protein
disulfide isomerase during thrombus formation in mice. J Clin Invest 118:
1123-1131. 2008
10. Creemers EE, Cleutjens JP, Smits JF, Daemen MJ: Matrix metalloproteinase
inhibition after myocardial infarction: a new approach to prevent heart failure?. Circ
Res 89: 201-210, 2001
11. Davì G, Patrono C: Platelet activation and atherothrombosis. N Engl J Med
357,:2482-2494, 2007
12. Diwan A, Tran T, Misra A, Mann DL: Inflammatory mediators and the failing
heart: a translational approach. Curr Mol Med 3: 161-182, 2003
13. Dorn GW 2nd, Apoptotic and non-apoptotic programmed cardiomyocyte death in
58
14. Eapen Z, Rogers JG: Strategies to attenuate pathological remodeling in heart failure.
Curr Opin Cardiol 24: 223-229, 2009
15. Farndale RW, Sixma JJ, Barnes MJ, de Groot PG: The role of collagen in
thrombosis and hemostasis. J Thromb Haemost 2: 561-573, 2004
16. Fuster V, Sweeny JM: Aspirin: a historical and contemporary therapeutic overview.
Circulation 123: 768-778, 2011
17. Gadi D, Bnouham M, Aziz M, Ziyyat A, Legssyer A, Legrand C, Lafeve FF, Mekhfi
H: Parsley extract inhibits in vitro and ex vivo platelet aggregation and prolongs
bleeding time in rats. J Ethnopharmacol 125: 170-174, 2009
18. Gao E1, Lei YH, Shang X, Huang ZM, Zuo L, Boucher M, Fan Q, Chuprun JK, Ma
XL, Koch WJ: A novel and efficient model of coronary artery ligation and
myocardial infarction in the mouse. Circ Res 107: 1445-1453, 2010
19. Garg A, Garg S, Zaneveld LJ, Singla AK: Chemistry and pharmacology of the
Citrus bioflavonoid hesperidin. Phytother Res 15: 655-669, 2001
20. González A, Ravassa S, Beaumont J, López B, Díez J: New targets to treat the
structural remodeling of the myocardium. J Am Coll Cardiol 58: 1833-1843. 2011
21. Goto S: Cilostazol: potential mechanism of action for antithrombotic effects
accompanied by a low rate of bleeding. Atheroscler Suppl 6: 3-11, 2005
22. Han Y, Li Y, Wang S, Jing Q, Wang Z, Wang D, Shu Q, Tang X: Cilostazol in
59
coronary intervention in patients with acute coronary syndromes: a randomized,
controlled study. Am Heart J 157: 733-739, 2009
23. Hardy AR, Jones ML, Mundell SJ, Poole AW: Reciprocal cross-talk between P2Y1
and P2Y12 receptors at the level of calcium signaling in human platelets. Blood
104: 1745-1752, 2004
24. Hirota R, Roger NN, Nakamura H, Song HS, Sawamura M, Suganuma N:
Anti-inflammatory effects of limonene from yuzu (Citrus junos Tanaka) essential oil on
eosinophils. J Food Sci 75: H87-H92, 2010
25. Jennings LK: Mechanisms of platelet activation: need for new strategies to protect
against platelet-mediated atherothrombosis. Thromb Haemost, 102: 248-257. 2009
26. Jessup M, Brozena: Heart failure. N Engl J Med 348: 2007-2018, 2003
27. Jin YR, Han XH, Zhang YH, Lee JJ, Lim Y, Chung JH, Yun YP: Antiplatelet
activity of hesperetin, a bioflavonoid, is mainly mediated by inhibition of
PLC-gamma2 phosphorylation and cyclooxygenase-1 activity. Atherosclerosis 194:
144-152, 2007
28. Kandasamy AD, Chow AK, Ali MA, Schulz R: Matrix metalloproteinase-2 and
myocardial oxidative stress injury: beyond the matrix. Cardiovasc Res 85: 413-423,
2010
29. Kido M, Du L, Sullivan CC, Li X, Deutsch R, Jamieson SW, Thistlethwaite PA: