Internal Transcribed Spacers (ITS)로 분석한 국내 Rubus 재배종의 계통학적 유연관계
유기석*,1·박명렬*,1·윤성중**,†
*전북대학교 농업생명과학대학 농업과학기술연구소
**전북대학교 농업생명과학대학 작물생명과학과, 농업과학기술연구소
Internal Transcribed Spacer (ITS) Regions Reveals Phylogenic Relationships of Rubus Species Cultivated in Korea
Gee Suck Eu*,1, Myoung Ryoul Park*,1, and Song Joong Yun**,†
*
Institute of Agricultural Science and Technology, Chonbuk National University, Jeonju 561-756, Korea.
**
Department of Crop Science, Institute of Agricultural Science and Technology, Chonbuk National University, Jeonju, 561-756, Korea.
ABSTRACT : The bramble cultivated widely in South Korea, which is known as Bokbunja-ddal-gi, is regarded as having originated from Korean native Rubus coreanus . This study was carried out to obtain basic phylogenetic information on Korean cultivated bramble (KCB) by comparing the internal transcribed spacer (ITS) regions with those of R. coreanus , blackberry ( R. lanciniatus ), black ( R. occidentalis ) and red ( R. idaeus ) raspberry. Sequences of the ITS 1 suggest that some KCB accessions share a significant similarity with both R. occidentalis and R. coreanus in the ITS 1 region. The ITS 2 sequences of the three KCB accessions clustered more closely to those of two R. occidentalis accessions than to those of R.
coreanus . These results suggest that there exist variations in the sequences of ITS among KCB accessions and KCB acces- sions are more closely related to black raspberry than R. coreanus in the ITS regions.
Key Words : Rubus Species, Phylogenetic Relationships, Internal Transcribed Spacer (ITS)
INTRODUCTION
Many species of the genus Rubus including raspberry, blackberry, and loganberries are collectively called bramble.
Korean endemic bramble have an excellent reputation for their medicinal effects. In Korea, R. coreanus , the Rubus species native to Korea and collectively called Bokbunja- ddal-gi or Bokbunja, has been used for various medicinal purposes for centuries. Dried fruit of R. coreanus has been used as remedies for palsy, imbecility, arthritis, stomach diseases, sexual disorders and cancer (Jeong and Sin, 1996;
Park et al. , 2004).
Recent interest on health foods and the scientific findings supporting the medicinal effects of bramble escalated demand for bramble in Korea. Accordingly, the production
and cultivation area of bramble has increased drastically in the last six years. However, though the commercial production of bramble in Korea is relatively new, as recent as the last five decades, little scientific documentation is available on its domestication or cultivation.
A majority of Korean cultivated bramble (KCB) is Bokbunja which has been known as a domesticated form of the Korea native R. coreanus. However, considerable morphological differences in flower and leaf are noticed between KCB and its claimed wild relative R. coreanus . A recent comparison of genetic background of Korea native and cultivated bramble with random amplified polymorphic DNA (RAPD) markers indicated that KCB share its genetic background more closely to black raspberry ( R. occidentalis ) than R. coreanus . (Eu et al ., 2008).
1
These two authors contributed equally to this study
†
Corresponding author: (Phone) +82-63-270-2508 (E-mail) [email protected]
It is imperative to develop new improved cultivars of bramble to satisfy customers' demands on bramble products at various forms and levels. Elucidation of genetic characteristics merits the first priority in genetic diversification and improvement of KCB. Though, we could get useful information on genomic feature of KCB from RAPD profiles (Eu et al ., 2008), more data are required for clear understanding on genetic features of KCB.
The nuclear ribosomal RNA genes (rDNA) which encode the 18S, 5.8S and 26S rRNA subunits are transcribed as a single unit along with the two spacers. The ITS regions are edited from the mature RNA transcript. Sequence variation in the ITS region is quite high and occurs even among closely related clones or species. Therefore, nucleotide sequences of the ITS of nuclear ribosomal DNA have been used as useful sources of molecular information for phylogenetic studies in many angiosperm families (Baldwin, 1992; Baldwin et al ., 1995), and have been applied in a large number of plant groups including grasses (Hsiao et al . 1994), cotton (Pillay and Meyers 1999), walnut (Stanford et al . 2000) and sunflower (Clevinger and Panero 2000). The phylogeny of Rubus was also generated based on ITS sequences (Alice and Campbell, 1999; Eriksson et al ., 1998). Therefore, the objective of this study was to obtain basic phylogenetic information about the origin of KCB by comparing ITS of KCB with those of R. coreanus , black and red raspberries and blackberries.
MATERIALS AND METHODS
1. Plant materials
A total of thirteen accessions of Rubus species were used in this study. Three accessions of KCB were selected based on random amplified polymorphic DNA (RAPD) makers profiles from plants cultivated in the major cultivation areas in South Korea. Black raspberry (Roc, R. occidentalis L., two cultivars) was obtained from National Clonal Germ- plasm Repository, Corvallis, Oregon, USA. R. coreanus (Rco, three species), red raspberry (Rid, R. idaeus , two species) and blackberry (Rla, R. lanciniatus , three species) were obtained from the Korean Black Raspberry Experiment Station, Gochang, Korea (Table 1).
2. Chemicals
DNA extraction kits were from Gentra Systems (Minneapolis, MN, USA). The DNA plasmid vector pGEM- T-Easy was purchased from Promega (Madison, WI, USA).
All chemicals and enzymes were purchased from Sigma (St.
Louis, MO, USA), unless otherwise indicated.
3. Extraction of genomic DNA
DNA was isolated from fresh young leaves using the Puregene DNA purification kit (Gentra System, Minneapolis, MN, USA) following the instruction provided by the manufacturer. Fresh leaf samples (30 ㎎ ) were ground with
Table 1.Rubus
accessions used for the analysis of the internal transcribed spacer regions.
Entry Number Common Name Taxon Collection location Note (Variety)
KCB 1
Korea Cultivated bramble (KCB) Unknown
South Korea
Gochang, Jeonbuk
KCB 8 Sanchung, Gyeongnam
KCB 11 Jeongeup, Jeonbuk
Rco 25
Korean bramble (Rco)
R. coreanus MiqueDanyang, Chungnam
Rco 26 Okcheon, Chungbuk
Rco 23 Gochang, Jeonbuk
Rid 52 Red raspberry (Rid)
R. idaeusUSA
Golden Harvest
Rid 54 Canby
Rla 56
Blackberry (Rla)
R. lanciniatusThorny
Rla 57 Creeping
Rla 58 Ebano
Roc 39 Black raspberry (Roc)
R. occidentalis L.Shuttleworth
Roc 50 NC 98-8-1
a mortar and pestle in liquid nitrogen. Cells of the ground leaf tissue were lysed by incubating in lysis solution at 65
℃ for 60 min. Cell lysate was collected by centrifugation and treated with proteinase K (6 ㎎ / ㎖ ) for 60 min at 55 ℃ . RNA was degraded by adding RNase A (1.5 ㎎ / ㎖ ) in the cell lysate and incubating the lysate at 37 ℃ for 15 min.
Proteins were precipitated by adding the protein precipitation solution to the cell lysate followed by inverting racks containing the samples for about 2 min and centrifuging at 13,000~16,000 × g for 3 min. DNA in the supernatant was precipitated using isopropanol. Precipitated DNA was collected by centrifugation and DNA pellet was washed with 70% ethanol. DNA pellet was dried and hydrated in 50 ㎕ DNA hydration solution by incubating the DNA sample overnight at room temperature. DNA concentration was measured by both spectrophotometric assay and gel electrophoresis. DNA samples were used for PCR for ITS analysis.
4. The Polymerase Chain Reaction (PCR) Condition PCR for ITS regions were carried out with the DNA template prepared from leaf samples. PCR amplification was carried out in a 30 ㎕ reaction mixture, containing 0.01~0.1 ng of template DNA, 10 pmol each primer, dNTPs (0.2 mM each), and Taq polymerase (1 unit). The thermal cycles consisted of an initial denaturation (5 min at 94 ℃ ), 40 cycles of amplification (repeated cycles of denaturation (1 min at 94 ℃ ), annealing (1 min at 55 ℃ ) and extension (1 min at 72 ℃ ), and a last extension (10 min at 72 ℃ ). The amplified products were separated in 1% agarose gel at 100 V for 30 min and visualized by staining the gel in ethidium bromide solution and photographed under UV light.
5. Analysis of internal transcribed spacers (ITS) of nuclear ribosomal DNA
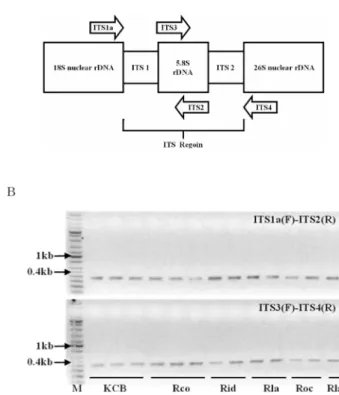
The ITS of nuclear ribosomal DNA were amplified using the specific primers targeting the ITS 1 and ITS 2 regions by PCR reactions (Fig. 1). The primers for the ampli-
fication of the ITS regions were designed as described by White et al . (1990, primers ITS1a (F) and ITS2(R)) and Kim and Kim (1999, primers ITS3 and ITS4) (Table 2).
The primer pair ITS1a (F) and ITS2 (R) or ITS3 (F) and ITS4 (R) was used to amplify the ITS 1 or ITS 2 region.
The amplification reaction was performed essentially the same as in PCR condition, except annealing at 55 ℃ for 1 min. The amplified products were separated in 1% agarose gel at 100 V for 30 min and visualized by staining the gel in ethidium bromide solution and photographed under UV light.
Genomic DNA fragments obtained by PCR amplification
Fig. 1
. Nuclear ribosomal DNA regions sampled (A) and DNA fragments amplified from the targeted internal transcribed sequences(ITS) (B). KCB; Korea cultivated bramble, Rco;
bramble native to Korea (
R. coreanusMique), Rid; red raspberry (
R. idaeus), Rla; Blackberry (
R. lanciniatus), Roc;
black raspberry (
R. occidentalisL.)
.Table 2.
The sequences of the primers used for amplification and sequencing of the internal transcribed sequence regions of nuclear ribosomal DNA.
Region Primer Direction Sequence (5'3') Reference
ITS 1 ITS1a Forward GGAAGGAGAAGTCGTAACAAGG Kim and Kim.(1999)
ITS2 Reverse GCTGCGTTCTTCATCGATGC Kim and Kim.(1999)
ITS 2 ITS3 Forward GCATCGATGAAGAACGCAGC Kim and Kim.(1999)
ITS4 Reverse TCCTCCGCTTATTGATATGC Kim and Kim.(1999)
were gel purified using AccuPrep Gel Purification Kit (BIONEER, Seoul, Korea). DNA fragments were cloned into the plasmid vector (pGEM-T-Easy, Madison, Promega).
The recombinant DNAs were transformed into E. coli strain JM109 by the standard CaCl
2method. Positive clones were grown in LB medium containing Ampicilin and plasmid DNAs were purified using the AccuPrep Pasmid Extraction Kit (BIONEER, Seoul, Korea). The insert DNA of the clones having about 0.37~0.39 kb (ITS 1) or 0.4~ 0.42 kb (ITS 2) insert were sequenced by the dideoxy chain termination method through commercial service.
6. Sequence and data analysis
The nucleotide sequences were blasted against the sequences in GenBank and annotated based on the sequence similarity. Sequence homology searches were performed using the program BLAST (Altschul et al ., 1997) against DNA and protein sequences in GenBank. Nucleotide and deduced amino acid sequence analyses were performed using DNASIS (Hitachi, USA), and the programs and databases offered by the National Center for Biotechnology Infor- mation (NIH, USA) and European Bioinformatics Institute (EBI, UK). Multiple sequence analysis was performed using the program AliBee (Brodsky et al ., 1995). The GC content
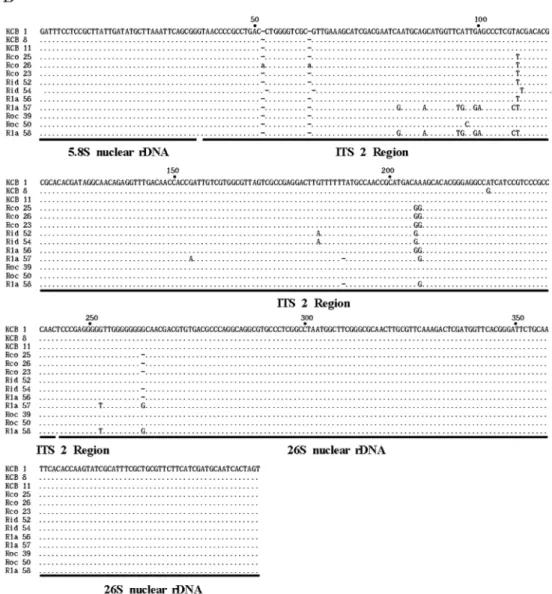
Fig. 2
. Aligned DNA sequence of the ITS 1 (A) and ITS 2 (B) regions in nuclear ribosomal DNA for thirteen accessions of Abbreviated names for each
Rubusaccession are the same as in Table 1.
Rubusspecies.
of sequence was analyzed using the SeqApp program (version 1.9a169, Gilbert, 1994).
RESULTS AND DISCUSSION
A total of thirteen accessions were selected from each Rubus species which formed a separate subclade by the RAPD markers (Eu et al ., 2008) for the analysis of the ITS of nuclear ribosomal DNA (Table 1). About 350~400 bp and 400~500 bp DNA fragments were amplified using the primer pairs targeted for the ITS 1 and 2, respectively (Fig. 1).
The amplified fragments for the ITS 1 and ITS 2 were cloned and their nucleotide sequences were determined and
their similarity to the known ITS sequences were calculated using the Basic Local Alignment Search Tool (BLAST) available at NCBI (http://www.ncbi.nlm.nih.gov/ BLAST/).
The nucleotide sequences showed over 98% similarity to those of the ITS 1 and 2 of the hybrid R. caesius x R.
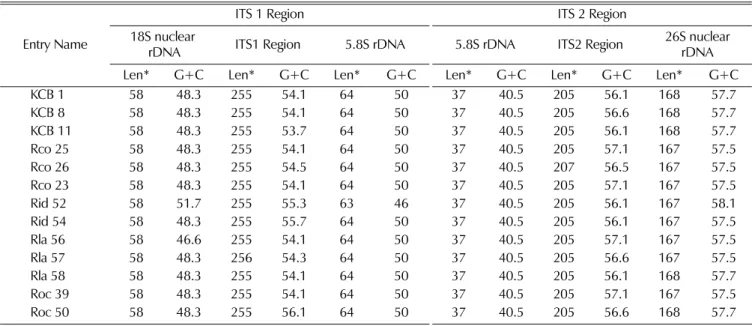
idaeus (Fig. 2). The nucleotide of the ITS 1 ranges from 376-378 bp in all Rubus species tested and the length of the ITS 2 varies from 409 to 411 bp in all Rubus species tested (Fig. 2, Table 3). In the DNA fragments amplified for the ITS 1, included also are a part of 18S and a part of 5.8S nuclear rDNAs. The partial 18S nuclear rDNA is 58 bp long and its average G+C content ranges from 46.6% to 51.7%. The ITS 1 region of all Rubus species is 255 bp except the accession Rla 57, which is 256 bp.
Fig. 2.
continued
Average G+C content of the ITS 1 region ranges from 53.1% to 56.1%. The partial 5.8S nuclear rDNA of all Rubus species is 64 bp long and its G+C content 50%.
Only one exception is Rid 52 whose partial 5.8S rDNA is 63 bp long and its G+C content 46% (Table 3).
In the DNA fragments amplified for the ITS 2, included also are a part of 5.8S and 26S nuclear rDNAs. The partial 5.8S nuclear rDNA is 37 bp long and its G+C content 40.5%. The length of the ITS 2 region of all Rubus species is 205 bp except the accession Rco 26 which is 207 bp. G+C content of the ITS 2 region ranges from 56.1% to 57.1%. The partial 26S nuclear rDNA of all Rubus species ranges from 167 to 168 bp long and its G+C content from 57.5% to 58.1% (Table 3).
The nucleotide sequences were closely examined using DNASIS program and aligned using the multiple alignment algorithms GeneBee (http://www.genebee.msu.su/genebee.html).
Alignment of the ITS 1 sequences required several one- to five-base gaps due to insertion/deletion (indel) mutations among the accessions of Rubus species (Fig. 2). There are 12 indel mutations in the ITS 1 region. There are five, three, one, two and one 1-, 2-, 3-, 4-, and 5-base indels, respectively. Also, there are six informative sites in the ITS 1 region. Contrastingly, alignment of the ITS 2 sequences required no gaps, indicating that there are no indel mutations among the Rubus species investigated in this study. There are three informative sites in the ITS 2 region
(Fig. 3).
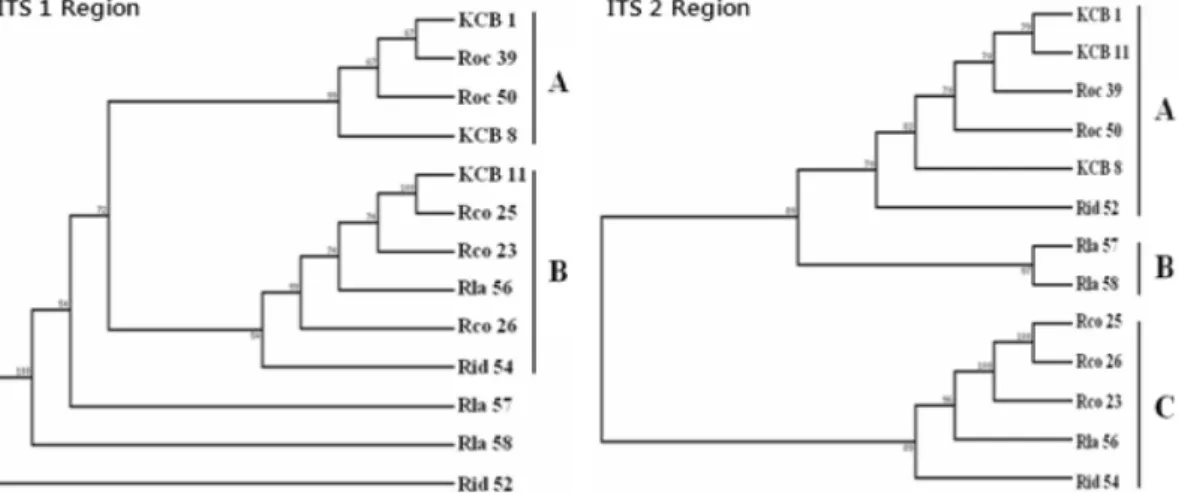
Phylogenetic trees were constructed using the ITS 1 and 2 nucleotide sequences. A phylogenetic tree based on the ITS 1 sequences contains a strongly supported clade and three outgroups. The clade incudes two subclades. The two KCB accessions, KCB 1 and KCB 8, are found nested within subclade A containing the two black raspberry accessions, Roc 39 and Roc 50. However, KCB 11 is found included in subclade B containing three R. coreanus, one red raspberry and one R. lanciniatus accessions. Two blackberry and one red raspberry accessions are found as outgroups (Fig. 3).
The phylogenetic tree based on the ITS 2 sequences contains a clade with three subclades. The three KCB accessions, KCB 1, KCB 8 and KCB 11 are found in subclade A containing the two black raspberry accessions, Roc 39 and Roc 50, and a red raspberry accession, Rid 52. Two blackberry accessions form subclade B. Subclade C includes all the three R. coreanus, and each one of red raspberry and blackberry accessions (Fig. 3). Nucleotide sequences of ITS 1 and 2 generated informative characters for phylogenetic relationship among a total of thirteen representative Rubus accessions selected from each subclade differenciated by RAPD markers.
The length of ITS 1 and ITS 2 of Rubus accessions used in this study ranges from 255 to 256 bp and from 205 to 207 bp, respectively. The length of the ITS 1 is
Table 3.
Size and base composition of the ITS 1 and ITS 2 regions of nuclear ribosomal DNA of
Rubusspecies.
Entry Name
ITS 1 Region ITS 2 Region
18S nuclear
rDNA ITS1 Region 5.8S rDNA 5.8S rDNA ITS2 Region 26S nuclear
Len* G+C Len* G+C Len* G+C Len* G+C Len* G+C Len* rDNA G+C
KCB 1 58 48.3 255 54.1 64 50 37 40.5 205 56.1 168 57.7
KCB 8 58 48.3 255 54.1 64 50 37 40.5 205 56.6 168 57.7
KCB 11 58 48.3 255 53.7 64 50 37 40.5 205 56.1 168 57.7
Rco 25 58 48.3 255 54.1 64 50 37 40.5 205 57.1 167 57.5
Rco 26 58 48.3 255 54.5 64 50 37 40.5 207 56.5 167 57.5
Rco 23 58 48.3 255 54.1 64 50 37 40.5 205 57.1 167 57.5
Rid 52 58 51.7 255 55.3 63 46 37 40.5 205 56.1 167 58.1
Rid 54 58 48.3 255 55.7 64 50 37 40.5 205 56.1 167 57.5
Rla 56 58 46.6 255 54.1 64 50 37 40.5 205 57.1 167 57.5
Rla 57 58 48.3 256 54.3 64 50 37 40.5 205 56.6 167 57.5
Rla 58 58 48.3 255 54.1 64 50 37 40.5 205 56.1 168 57.7
Roc 39 58 48.3 255 54.1 64 50 37 40.5 205 57.1 167 57.5
Roc 50 58 48.3 255 56.1 64 50 37 40.5 205 56.6 168 57.7
Len*; Length
similar to that reported for Rubus by Alice and Campbell (1999). The length of the ITS 2 is also similar to that reported for the five different Rubus species, including R.
deliciosus , R. rosifolius, R. parvus, R. caesius, and R.
macraei , reported by Alice and Campbell (1999) which ranges form 207 to 212 bp. This result indicates that the length of the ITS 1 and 2 of KCB and R. coreanus is similar to that of other Rubus species. It is known that there is a considerable variation in the length of the ITS 1 and 2 sequences among angiosperm taxa (Baldwin et al ., 1995).
Phylogenetic relationships inferred from ITS 1 and 2 sequences share a significant similarity with those from RAPD analysis (Eu et al ., 2008). The ITS 1 sequence of two KCB accessions, KCB 1 and KCB 8, cluster more closely to the two black raspberry accessions, Roc 39 and Roc 50. However, that of KCB 11 is closely related to that of the three R. coreanus, one red raspberry and one blackberry accessions. The sequence of two blackberry and one red raspberry accessions is most distantly related to each of them and to the rest of species. This result indicates that there are considerable variations in ITS 1 nucleotide sequence both within and among Rubus species used in this study. The result also indicates that some KCB accessions share a significant similarity either with black raspberry or R. coreanus in the ITS 1 region.
The ITS 2 sequences of three KCB accessions, KBC 1, KBC 8 and KBC 11, cluster more closely to two black raspberry accessions, Roc 39 and Roc 50, and a red raspberry accession, Rid 52 than to those of R. coreanus . This result implies that there are considerable variations in
the ITS 2 nucleotide sequences both within and among Rubus species used in this study. The result also indicates that some KCB accessions share a significant similarity with black raspberry in the ITS 2 region.
In summary, phylogentic relationships inferred from the molecular features of ITS DNA of Rubus species suggest that KCB accessions shows relatively closer relatedness to that of black raspberry than to that of R. coreanus . This result does not support the widely accepted ancestry of KCB to R. coreanus.
ACKNOWLEDGEMENT
This study was supported by Technology Development Program for Agriculture and Forestry, Ministry of Agriculture and Forestry, Republic of Korea. The authors express their thanks to Dr. Nahla Bassil (USDA ARS National Clonal Germplasm Repository, 33447 Peoria Road, Corvallis, OR 97333-2521, USA) for providing the protocol for genomic DNA preparation from the Rubus leaf samples.
LITERATURE CITED
Alice LA and Campbell CS. (1999). Phylogeny of
Rubus(Rosaceae) based on nuclear ribosomal DNA. American Journal of Botany. 86:81-97.
Altschul SF, Madden TL, Schaffer AA , Zhang J, Zang ZW, Miller W and Lipman DJ. (1997). Gapped BLAST and PSI- BLAST: a new generation of protein database search programs.
Nucleic Acids Research. 25:3389-3402.
Fig. 3.