국내 특수 생태환경의 탈질 저영양 세균의 종 다양성 및 생리적 특성 분석
이창묵1·원항연2·권순우2·강한철1·구본성1·윤상홍1*
1농촌진흥청 국립농업과학원 농업생명자원부, 2농촌진흥청 국립농업과학원 농업생물부 Received : July 18, 2011 / Revised : August 24, 2011 / Accepted : August 25, 2011
Analysis of Species Variety and Physiological Characteristics of Denitrifying Oligotrophic Bacteria Iso- lated from the Specific Environment in Korea. Lee, Chang-Muk1, Hang-Yeon Weon2, Soon-Wo Kwon2, Han-Chul Kang1, Bon-Sung Koo1, and Sang-Hong Yoon1*. 1Functional Biomaterial Division, 2Agricultural Microbiology Team, National Academy of Agricultural science, Rural Development Administration, Suwon 441- 707, Korea − In an effort to isolate novel bacteria for the bioremediation of over-fertilized soils, we identified 135 denitrifying cells out of 3,471 oligotrophic bacteria pools (3.9%) using a denitrification medium supple- mented with potassium nitrate as the sole nitrogen source. Soil samples were taken from ecologically well- conserved areas, including a mountain swamp around the demilitarized zone (Yongneup), two ecoparks (Upo and the Mujechi bog), and ten representative islands around the Korean peninsula (Jejudo, Daecheongdo, Socheongdo, Baekryeongdo, Ulrungdo, Dokdo, Geomundo, Hongdo, Huksando and Yeonpyeongdo). All of the 135 bacteria produced nitrogen gas from the denitrification medium, and were proved to be nitrate reduc- tase positive by API-BioLog tests. Phylogenetic analysis using 16S rDNA sequences revealed that the 135 bacteria consisted of 44 different genera. Along with the most prominent, Proteobacteria (87.4%), we identi- fied denitrifying bacteria from Firmicutes (9.4%), Actinobacteria (2.4%), and Bacteroidetes (0.8%). Physio- logical analyses of the 44 representative denitrifying bacteria, under various pH levels, growth temperatures and salt stresses, revealed 12 favorable denitrifying strains for soil bioremediation.
Key words: Oligotrophic bacteria, denitrifying bacteria, activity
서 론
토양 중에는 다양한 미생물이 존재하고 있지만, 아직 얼 마나 다양한 미생물이 특정 환경에 존재하는지 모르고 있다 . 그러나 생태계에서 물질순환이나 식물생육 등에 세균이 행 하고 있는 역할을 생각해 본다면 다양한 미생물이 존재하는 환경 시료를 대상으로 연구하는 것이 매우 중요하다고 할 수 있다[17]. 최근에는 생태 환경에 존재함에도 현재의 기술로 배양되지 못하거나 매우 까다로운 미생물이 전체 미생물의 99%에 이른다는 사실이 보고되어 있다[1, 18]. 이러한 난배 양 미생물이 새로운 기능성 물질 또는 유전자 탐색의 가장 풍부한 자원의 하나라는 사실이 알려져 있다[6, 13].
지금까지 분리되어 연구된 세균은 고농도 영양조건에서 급속히 증식하는 것이 대부분이다. 그러나 자연환경에 서식 하고 있는 난 배양 미생물에 대한 연구 결과를 살펴보면 저 농도 영양조건하에서 매우 느리게 증식하는 세균이 자연환 경 내에 다량 존재하고 이들이 미생물 생태 환경에서 차지
하는 역할에 대한 중요성이 부각되어 많은 연구가 진행되어 왔다[12, 15, 21]. 이렇게 영양분이 매우 적은 환경에서 자 랄 수 있는 세균을 저영양 세균(oligotrophic bacteria)이라고 하는데 저영양 환경이라는 개념이 매우 폭넓고 다양하기 때 문에 저영양 세균에 대한 정의는 모호한 상태이다. 또한, 이 들 저영양 세균은 분리하기가 대부분 매우 까다롭고 대량 배 양이 잘되지 않고, 배양 도중에 우점균에 의해 소실되는 경 우가 많기 때문에 분류학적 및 생리적 특성이 충분히 해명 되지 않은 채 난배양 미생물의 일부로 분류되고 있다[3, 4].
국내 토양 자원은 심각하게 산성화되어 있음이 알려져 있 다. 토양의 산성화 원인은 식물의 성장에 필요한 10여 가지 원소 중 질소, 인, 칼륨을 화학비료로 공급하고 이들이 흙 속에서 차츰 산화되어 질산, 인산, 황산 등으로 변환되는 것 에 기인한다[9]. 또한, 최근에는 산성비의 증가에 따라 토양 의 산성화가 가속화 되고 있다. 토양 산성도의 증가는 토양 미생물 종의 감소를 초래하며, 특히 토양 중에 존재하는 질 소 순환에 관여하는 미생물의 성장이 산성에 민감할 수 있 다. 따라서 토양 질소 순환에 문제가 생길 경우 화학비료의 계속적인 처리를 필요로 하는 악순환을 형성시키게 된다. 아 직 토양의 탈질소에 관한 난배양 미생물의 영향에 대한 연 구는 난배양 미생물 군이 있다는 정보 이외에 잘 알려져 있
*Corresponding author
Tel: +82-31-299-1691, Fax: +82-31-299-1672 E-mail: [email protected]
지 않다. 지금까지 알려진 질소 순환에 관여하는 미생물들 은 일련의 탈질소 관련 효소들을 가지고 있는 것이 알려져 있다[2, 8]. 따라서 난배양 미생물 유전자원 가운데에 전혀 연구되어 있지 못하면서 탈질소 관련 유용 유전자의 conserved domain이나 motif에 해당하는 신규 유전자가 대 량으로 있을 것이 추정되고 있다. 최근에는 생물정보학의 기 술적 개발을 바탕으로 질소순환 관련 유용 유전자군의 특징 을 포함하는 probe를 작성할 수 있으며[7], 이 probe를 microarray로 구성하면 질소순환(질소 고정 및 탈질소 과정 포함) 난배양 미생물 유전자원을 대량으로 검출 할 수 있을 것으로 예상하고 있다[5, 19].
따라서 본 연구에서는 국내 토양 가운데 생태환경이 잘 보존된 지역에서 분리된 저영양 세균 자원으로부터 탈질 세 균을 선발하여 이들의 종 다양성과 생태적 특성을 분석함 으로써 질소가 과다 축적된 생태환경을 선순환시키기 위한 새로운 미생물 및 유전자원 개발의 기초자료를 제공하고자 한다.
재료 및 방법 시료 채취와 미생물 배양 배지
시료는 한국의 북부지방, 도서지방, 특수 생태지역의 생태 가 잘 보존된 토양을 사용하였다. 각 시료의 채취 GPS; 용 늪(E:128o07'30'' N:38o12'53''), 백령도(E:124o53' N:37o52'), 대청도(E:124o53' N:37o53'), 소청도(E:124o55' N:37o52'), 홍 도(E:125o12' N:34o41'), 흑산도(E:125o26' N:34o41'), 연평도 (E:125o43' N:37o40'), 제주도(E:126o58' N:33o12'), 거문도 (E:127o11' N:34o00'), 우도(E:128o25' N:35o33'), 독도(E:131o52' N:37o14'), 울릉도(E:131o52' N:37o30'). 세균의 증식은 액체 영양(nutrient broth, NB) 배지와 NB배지를 희석한(diluted nutrient broth, DNB) 배지를 사용하였다. 표준 NB배지의 조 성은 beef extract, 10 g; peptone, 10 g; NaCl, 5 g; distilled water, 1000 mL이며 pH 7.0~7.2로 조정하였다. 그리고 NB 배지를 10-1배, 10-2배, 10-3,10-4배로 증류수에 희석한 4종류 의 희석 DNB 배지를 사용하였다.
저영양 세균의 분리
R2A 배지를 10-1배 희석한 배지 또는 10-4배 희석된 DNB 평판배지 위에 형성된 집락체로부터 토양세균을 각각 순수분 리 하여, 동일 농도의 DNB 10 mL 액체배지에 분리 균주를 전 배양한 후 통상농도의 10-1NB, 10-2NB, 10-3NB 및 10-4 NB 배지에 접종하여 28oC, 1주일간 배양 후 증식 능력을 판정하였다. 통상농도의 NB 배지에서 증식하지 못하는 세 균 중 10-1~10-4배로 희석한 배지에서 증식하는 세균을 저 영양 세균으로 순수분리 하였다. 순수 분리된 저영양세균은 20% 글리세롤이 첨가된 배지에 넣어 -80oC에서 장기 보존 하였다.
탈질소 세균의 선발
탈질소 세균은 nitrate 질소 배지에서 질소 기포를 형성하 며 동시에 nitrate reductase 활성이 양성일 경우를 최종 탈 질 균으로 판정하였다. 분리된 저영양 세균을 R2A 저영양 배지에서 이틀 동안 28oC 동안 충분히 배양한 뒤, 일정량을 Nitrate 질소 배지(Peptone 5 g/L, Meat extract 3 g/L, KNO3
1 g/L, pH 7.0)에 희석하여 nitrate를 제거하면서 동시에 질소 가스가 발생하는 과정을 관찰하여 탈질소 기능을 가진 세균 을 선별하였다. 먼저 nitrate 질소 배지를 멸균 처리 후 저영 양 세균을 접종하기 위하여 접종 전에 100oC 끓는 물에서 5분간 처리하여 용존 산소를 제거한 뒤, nitrate 배양액의 1%
(v/v) 되는 seed culture를 접종하였다. 산소와의 접촉을 피 하기 위하여 배지의 표면에 멸균한 mineral oil(1 mL)을 도 포하고, 발생한 질소 가스를 포획하기 위해 미리 멸균한 Durham tube를 배지에 뒤집어 seed culture와 같이 정치 배 양하였다. 이 상태에서 28oC에서 2~7일간 질소 기포 발생을 관찰하였다. 저영양 세균이 충분히 성장한 뒤 배지에 잔류 하는 산소가 소모된 뒤, 세균이 anaerobic 호흡을 수행하여 질소 가스가 기포로 보이는 균을 탈질소 기능을 가진 균으 로 1차 판정하였다.
Durham 튜브에 포집된 기체가 질산염 NO3−를 N2로 환원 시키는 대사과정 효소 중 하나인 nitrate reductase 효소에 기 인한다는 것을 확인하기 위하여 기포를 형성하는 저영양 세 균을 대상으로 각 액체 배지 시료를 일일 단위로 채취하여 nitrate reductase 활성을 검사하였다. 효소 활성은 8 g 설파 닐 산(sulfanilic acid, Fluka)을 5 N 아세트산 1리터에 녹여 서 상온에서 암 상태로 보존한 “시약 A”와 6 g의 N,N- Dimethyl-1-naphthylamine (Fluka)을 5 N 아세트산 1 리터 에 녹여 4oC 암 상태 보존한 “시약 B”를 사용하였다. 상기 추출한 액체 시료 10mL에 “시약 A”와 “시약 B”를 각 동일 한 양(5방울) 떨어뜨려 잘 혼합하였다. 약 1분 후 상온에서 배양액의 색깔이 붉은색으로 변화하는 것을 관찰하여 nitrate reductase 양성으로 판단하였다. 이 반응은 탈질소 배지에 포 함된 질산염(NO3−) 일부가 균의 nitrate reductase에의해서 아질산염(NO2−)으로 환원되었음을 나타낸다. 음성으로 나타 난 균들에 대해 배지 속의 아질산염이 균의 탈질소 과정에 서 완전히 소모되어 상기 방법으로 검출되지 않는지 재확인 하기 위하여, 색깔 변화가 없는 균에서 나온 액체배지시료 에 미량의 아연(Zn)을 촉매제로 더 첨가하였다. 이 촉매반응 은 배지에 잔존하는 질산염(NO3−)을 아질산염(NO2−)로 환원 시키는 역할을 한다. 따라서 아연 촉매하에서도 색깔의 변 화가 없는 경우는 탈질소 균에 의해 아질산염(NO2−)이 완전 히 소모되어 nitric oxide(NO) → nitrous oxide(N2O) → 질 소(N2)의 과정을 경유하여 탈질소 반응이 일어났음을 나타 낸다. 아연 촉매 반응에서 색깔의 변화가 없는 것을 또한 nitrate reductase 양성으로 판단하였다.
탈질소 세균의 16S rRNA 유전자 염기서열 분석
탈질소 기능을 가진 세균의 동정을 위해서 16S rRNA 유 전자 염기서열분석을 이용하였다. DNA extraction kit (Toyobo, Japan)로 분리균주의 DNA를 추출한 후 universal primer 27F(5'-AGA GTT TGA TCM TGG CTC AG-3') 와 1492R(5‘-TAC GGY TAC CTT GTT ACG ACT T-3’) 을 이용하여 16S rDNA 유전자를 증폭하였다[16]. PCR 산 물은 DNA sequencing kit(BigDye terminator Cycle Se- quencing Ready Reacions v. 3. 1.; Applied Biosystem)를 사용하여 반응시킨 후, Genetic Analyser 3100(Applied Bio- systems)로 염기서열을 분석하였다. 계통 분석도는 Kimura’s two-parameter model(1,000 replicate bootstrap value)에 따 른 Neighbor-joining과 Maximum-parsimony clustering 결 과를 MEGA 4 분석프로그램을 사용하여 작성하였다[20].
탈질소 세균의 생존 특성 분석
분리한 탈질소 균은 호기성 상태에서의 온도, pH, 염, 질 산염에 대한 생리적 적응능력이 조사되었다. 온도 조건은 4~50oC 범위에서, pH는 4~10, NaCl 염의 농도는 2~12%, 무기질소염 농도는 1~10% 에서 생존 여부를 R2A 배지를 기 본 배지로 하여 평판 배지와 액체 배지에서 각각 세 번 반복 실험을 통하여 생리적 생존 능력을 결정하였다. 온도에 따른 생존 특성을 측정할 때는 1% NaCl이 첨가된 R2A(pH 8.0)를 기본 배지조건으로 하였다. pH에 대한 생존 특성을 측정 할 경우에는 1% NaCl이 첨가된 R2A를 기본 배지조건으로 30oC에서 배양하였다. NaCl염과 무기질소염에 대한 저항성 은 R2A(pH 8.0)를 기본배지로 온도 30oC, pH 8.0하에서 24 시간 액체 배양하여 균 성장이 O.D. 600 nm에서 0.6 이상 일 경우를 생존한 것으로 판단하였다.
결과 및 고찰 탈질소 균의 선발
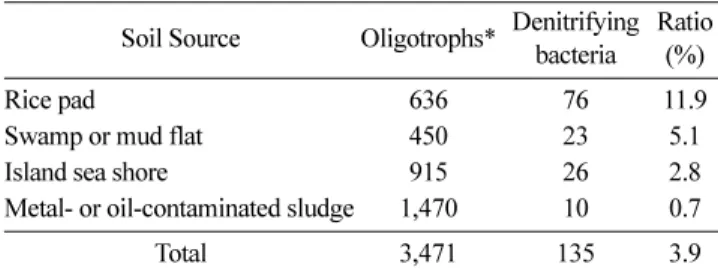
국내 생태환경이 잘 보존된 다양한 지역의 토양에서 분리한 총 3,471 개주의 저영양 미생물 자원으로부터 탈질소 능력을 가진 135균주를 선발하였다(Table 1). 저영양 세균을 수집한 여 러 토양 시료 중 논 토양은 저영양 배지에서 배양 가능한 균 의 11.9%가 탈질균으로 선발되어 가장 높은 빈도를 나타내었 다. 이는 질소시비가 많은 작물 경작지 토양에서 다른 토양보 다 탈질균이 우점하리라는 예상과 일치한 결과였다. 반면 비교 적 생태환경이 잘 보존된 늪이나 섬에서는 약 5.1%의 선발률 을 보였고 중금속 오염 토양에서는 0.7%의 가장 낮은 선발률 을 보였다. 선발된 135개 균주는 모두 질산염 첨가배지의 혐 기성 조건 배양에서 질소 기포를 형성하였고, 질산염(NO3−)을 아질산염(NO2−)으로 환원하는 nitrate reduction 활성이 있음을 확인하였다(Fig. 1). 이는 질소가스 발생에 의한 탈질 과정이 일련의 질산염 환원반응 효소들이 관여하고 있음을 시사해준다.
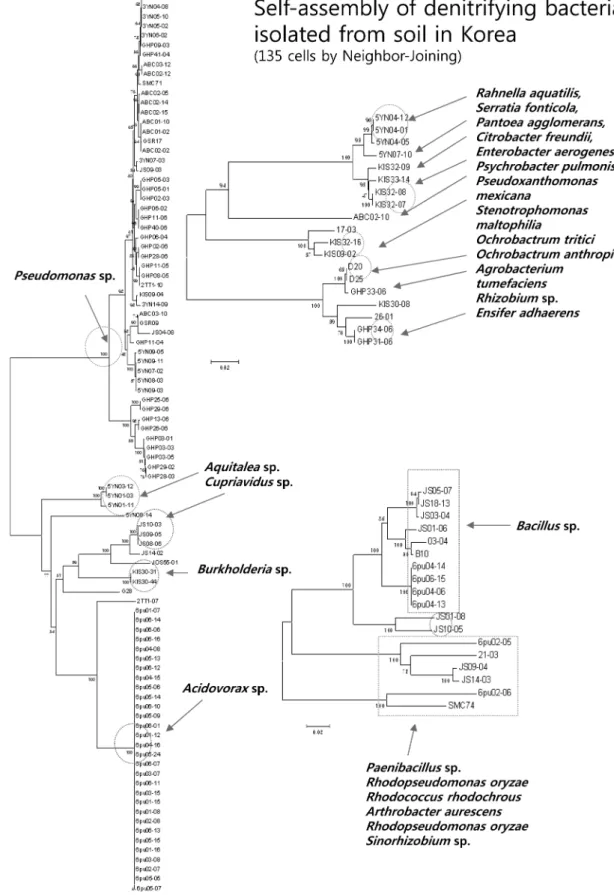
탈질소 균의 종 다양성 분석
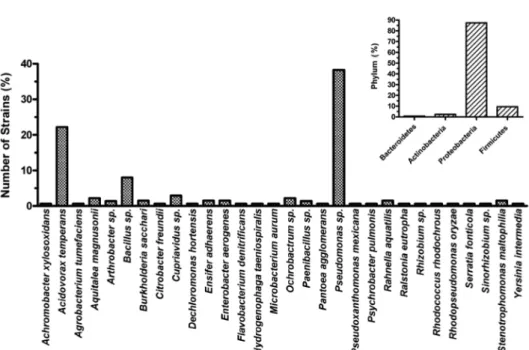
선발된 135 주의 탈질소 균의 종(Species)별 다양성 분석 을 위해 이들 균의 genomic DNA를 분리하여 상기 설명한 16S rRNA 프라이머를 사용하여 PCR로 증폭한 산물을 염기 서열 분석하여 균 동정에 사용하였다. GenBank와 Ribosome Database Project(http://rdp.cme.msu.edu)의 정보를 이용하여 계통분류를 통한 종 분석을 수행하였다. 그 결과, 이미 선발 된 135 주의 탈질소 균은 크게 44개의 속(Genus)에 대부분 포함되었으며(Table 2), 특히 Pseudomonas sp., Acidovorax sp., Bacillus sp.속이 전체의 90% 이상을 차지하였다(Fig.
2). 이들 속은 Proteobacteria에 속하는 문(phylum) 인 것으 로 알려져 있다(Fig. 3).
탈질소 균 속(Genus) 별 스트레스에 대한 생존 특성 분석 분리 동정된 44개 속에 속하는 탈질 미생물 그룹에서 한 개 Table 1. Screening of denitrifying cells from various soils in Korea.
Soil Source Oligotrophs* Denitrifying bacteria
Ratio (%)
Rice pad 636 76 11.9
Swamp or mud flat 450 23 5.1
Island sea shore 915 26 2.8
Metal- or oil-contaminated sludge 1,470 10 0.7
Total 3,471 135 3.9
*Soil samples (20 g) were processed with the 2 mm mesh before dilution into 0.85% NaCl. The resuspended soils were centrifuged for 30 min at 150 rpm. The supernatant was diluted serially (102~105) to spread on R2A minimal agar plates. The plates were incubated at 28oC for 5 days to isolated colonies. Well-isolated single colony was subject to denitrifying test as shown in Materi- als and Methods.
Fig. 1. Detection of denitrifying soil bacteria. (A) Nitrogen gas was collected from soil bacteria grown at 28oC for 7 days in nitrate broth (1% KNO3). To enhance anaerobic respiration, boiled nitrate broth was covered with sterile mineral oil after cell inoculation.
Nitrogen gas entrapped in a Durham tube is shown (red circle). (B) Nitrate reduction test from the nitrogen gas-producing cells. Sul- fanic acid was added to the nitrate culture broth to generate nitrate- sulfanic acid, followed by adding α-naphthylamine to precipitate red prontosil. From left; G27, 5-23, 4-8, 1B10, D25, 5-23, 14-9, negative control without inoculation.
의 균을 선택하여 개별적으로 여러 종류의 스트레스(온도, pH, 염농도, 질소 성분)에서의 생존 능력을 비교하여 환경적응성 이 우수한 탈질소 균을 선발하였다(Fig. 4). 이들 탈질 균들은 환경 적응성이 뛰어나 질소로 과다 오염된 토양의 친환경적 복구에 사용 시 적응 능력이 우수할 것으로 예상하고 있다.
분리된 탈질소 균들의 생존 온도영역은 대부분 10~30oC
에서 자랄 수 있지만 D20(Ochrobactrum tritici), JS14-O2 (Ralstonia eutropha), SMC74(Sinorhizobium sp.)는 최적 성장온도인 30oC의 매우 좁은 영역에서(28-30oC) 생존하는 특성을 가져 실용성이 떨어지는 것으로 생각된다. 그러나 6PU04-06(Bacillus pumilus) 균주는 10~50oC의 매우 넓은 영역에서도 생존이 가능하였다. 저항성을 가지는 pH 영역의 Table 2. Growth evaluation of the denitrifying soil bacteria.
Strain IDa Primary Identification
Growth rangeb Temp (oC)
(Min.~Max.)
pH (Min.~Max.)
NaCl (Max. %)
NaNO2 (Max. %)
KNO3 (Max. %)
03-04 Bacillus niacini 10~30 5~9 2.9 0.70 1.5
17-01 Pseudoxanthomonas mex 10~30 6~9 2.9 0.035 3.0
26-1 Rhizobium sp. 4~42 5~9 2.9 1.05 4.0
2TT1-07 Hydrogenophaga taeniosp 10~42 8~9 2.9 0.035 1.0
3YN07-03 Pseudomonas chlororaphis 10~42 4~10 5.8 2.10 9.5
3YN14-09 Pseudomonas syringae 4~30 5~9 2.9 0.035 1.5
5YN01-03 Aquitalea magnusonii 10~40 5~9 1 0.70 9.5
5YN04-01 Rahnella aquatilis 10~30 5~9 2.9 0.35 4.0
5YN07-10 Serratia fonticola 10~30 5~9 5.8 0.70 7.5
5YN08-14 Dechloromonas hortensis 4~30 5~9 2.9 0.35 2.0
5YN09-11 Pseudomonas fluorescens 4~30 5~9 2.9 0.35 2.0
6PU02-05 Microbacterium aurum 10~42 4~9 5.8 0.70 5.5
6PU04-06 Bacillus pumilus 10~50 4~10 11.6 0.70 3.0
6PU05-07 Acidovorax temperans 10~30 6~9 2.9 0.35 1.0
ABC02-05 Pseudomonas grimontii 4~30 5~9 2.9 0.70 3.0
ABC02-10 Pychrobacter pulmonis 4~30 8~10 5.8 2.10 3.5
B6 Bacillus casamancensis 10~42 6~9 5.8 0.35 8.0
D20 Ochrobactrum tritici 28~30 5~9 2.9 1.40 6.0
G28 Achromobacter xylosoxida 4~42 5~9 2.9 0.70 5.5
GHP03-03 Pseudomonas citronellolis 4~42 5~9 2.9 1.05 5.5
GHP05-01 Pseudomonas putida 4~42 5~9 5.8 1.75 7.5
GHP09-03 Pseudomonas veronii 4~30 5~9 2.9 0.35 2.5
GHP25-06 Pseudomonas azelaica 10~30 5~9 2.9 0.70 5.5
GHP26-06 Pseudomonas nitroreduce 10~42 5~9 2.9 1.05 4.0
GHP33-06 Ochrobactrum anthropi 10~42 5~9 2.9 1.05 4.0
GHP34-06 Ensifer adhaerens 10~30 6~9 2.9 0.35 2.5
JS01-06 Bacillus djibelorensis 4~30 8~9 5.8 1.75 3.0
JS01-08 Paenibacillus telluris 10~30 6~10 5.8 0.35 2.0
JS03-04 Bacillus mycoides 10~42 6~9 2.9 0.35 5.5
JS04-08 Pseudomonas fulva 10~42 5~9 5.8 2.10 6.5
JS05-07 Bacillus weihenstephanens 10~30 5~9 2.9 1.05 5.5
JS08-06 Cupriavidus basilensis 10~30 5~9 2.9 0.70 2.0
JS09-03 Pseudomonas rhodesiae 4~30 5~9 2.9 0.035 2.5
JS09-04 Arthrobacter ramosus 4~30 5~9 5.8 0.35 9.0
JS10-05 Paenibacillus terrae 10~30 5~9 2.9 0.35 3.0
JS14-02 Ralstonia eutropha 28~30 5~9 5.8 0.35 2.5
JS14-03 Arthrobacter aurescens 4~30 5~9 2.9 0.35 6.5
KIS09-02 Stenotrophomonas maltop 4~42 5~9 2.9 0.35 4.0
KIS30-08 Agrobacterium tumefacien 10~30 6~9 2.9 0.35 2.5
KIS30-31 Burkholderia sacchari 10~42 5~10 2.9 0.35 9.5
KIS32-09 Pantoea agglomerans 10~42 5~9 5.8 1.05 6.0
KIS33-14 Citrobacter freundii 4~42 5~9 5.8 2.10 5.5
SMC71 Pseudomonas lurida 4~30 5~9 5.8 0.70 3.0
SMC74 Sinorhizobium sp. 28~30 8~9 2.9 0.35 2.5
aThe applicable 12 denitrifying strains are underscored.
bPositive growth is determined if OD600 value reached more than 0.6 in R2A broth for 24 hrs under given stress conditions.
Fig. 2. Neighbor-joining tree constructed based on the partial 16S rDNA gene sequences of the strains isolated in this study. The strains represented the three major groups consist of each sharing the same genus; Pseudomonas sp., Acidovorax sp. and Bacillus sp. The bootstrap values (>50%) for 500 replicates are indicated next to the branches.
경우, 대부분의 균주가 pH 5~9 범위에서는 생존에 이상이 없었으나 2TT1-07(Hydrogenophaga taeniosp), JS01-06 (Bacillus djibelorensis), SMC74(Sinorhizobium sp.)는 pH 8-9의 매우 좁은 영역에서만 생존하는 특성을 보여 산성토 양이 대부분인 한국토양에서는 적합하지 않다고 할 수 있다.
그러나 JS01-06의 경우, 염에 대한 저항성이 다른 균들보다 우수한 특성을 보여주었다. 대부분의 균은 염(NaCl)농도 6%
이상에서 삼투압을 견디지 못하고 성장이 중지되었다. 그러 나 6PU04-06(Bacillus pumilus)는 10% 이상의 소금에서도 잘 자라는 내염성을 가짐으로써 이 균주가 상기에서 설명한 넓은 온도와 pH 범위에서도 생존하는 탈질균임을 감안할 때
실제 현장에 적용할 때 탈질소 능력은 강하지 않아도 많은 장점이 있을 수 있음을 보여준다. 반면에 무기 질산염의 농 도에 따른 균의 생존 능력을 조사하였을 때, 균에 따라 큰 차이가 나타나는 것을 알 수 있었다. 특히 JS04-08, KIS30- 31, JS09-04, 3YN07-03 같은 균은 매우 높은 질소 스트레 스 하에서도 생존할 수 있다는 것을 확인하였다(Table 2). 이 는 균들의 탈질 능력과 더불어 대사과정 차이에 의한 것으 로 판단되었는데, 미생물은 질산염을 사용하여 단백질 합성 같은 대사과정에 질소 성분으로 이용하는 것이 잘 알려져 있 기 때문이다. 본 연구의 결과에서 높은 농도에서 잘 생존한 다는 것은 질소 과다 축적된 토양에서 실제로 이용할 수 있 Fig. 3. Taxonomic identification of the 135 denitrifying soil bacteria strains. Full-length 16S rDNA sequences were compared to RDP database (http://rdp.cme.msu.edu) to identify the closest genus for each strain. The most representative phylum was proteobacteria.
Fig. 4. Stress response of the denitrifying bacteria to pH, temperature, nitrite and nitrate. With 135 denitrifying soil bacteria, 44 genus groups were assembled by 16S rDNA clustering. Those 44 representatives from each genus were examined for the growth ability under various stress conditions. Each spot denotes the maximum survival at the specific temperature, pH, and either nitrite (A) or nitrate (B). Cir- cles represent top 6 denitrifying strains selected.
다는 것을 의미할 뿐이지 이들이 반드시 탈질소 활성이 높 다는 것을 의미한다고 볼 수 없다[14]. 그럼에도 이들 질산 염에 저항성을 보이는 균들은 유기 질소 성분이 과다하게 오 염된 토양에서도 생존능력이 높을 것이 예상되므로 토양 정 화 과정에 매우 유리하게 사용할 수 있다.
전체적으로 종합하면 본 연구에서 선별한 탈질 균들 가운 데 간척지토양에서 적용될 탈질 미생물로는 6PU04-06 (Bacillus pumilus)이 가장 적합한 것으로 생각된다. 이외에 한 국의 기후(10~42oC)나 토성(pH 5 이상)에 비교적 적합한 탈 질 균으로는 총 12 균주가 선발되었다. 질산염(NO3−)을 분해 하며 높은 염 농도에서도 강한 저항성을 보이는 균주는 JS14- 03(Arthrobacter aurescens), JS09-04(Arthrobacter ramosus), B6(Bacillus casamancensis), KIS30-31(Burkholderia sacchari), 3YN07-03(Pseudomonas chlororaphis), JS04-08(Pseudomo- nas fulva), GHP05-01(Pseudomonas putida), 5YN07-10 (Serratia fonticola)의 8균주가 있었다. 아질산염(NO2−)에 강 한 저항성을 보이는 균주로는 JS01-06(Bacillus djibelor- ensis), KIS33-14(Citrobacter freundii), ABC02-10(Pychro- bacter pulmonis)가 포함되었다. 특히 Pseudomonadaceae에 속하는 3YN07-03, GHP05-01, JS04-08은 nitrate과 nitrite 모두에서 강한 저항성을 보여주었다(Table 2). 분리한 135종 의 탈질소 균 가운데 기존 학계에 보고되지 않은 신종 균을 파악하기 위하여 16S rRNA 염기서열을 사용하여 계통분석 을 수행하였다. 그 결과 5YN1-3(Aquitalea sp.)의 새로운 신 종 균을 발견하였다. 특히 Aquitalea 속에 속하는 5YN1-3균 의 경우, 지금까지 단 한 종의 Aquitalea 균만이 학계에 보 고되어 있었는데, 본 연구에서 탈질소 능력을 가진 신종 Aquitalea 균주를 확보할 수 있었다[10, 11].
요 약
국내의 대표적 섬이나 생태환경이 잘 보존된 토양으로부 터 저영양세균 3,471주를 분리하였고 이로부터 질소발생 분 석법에 따라 탈질균 135주를 최종 선발하였다. 이들 균주의 16S rDNA를 염기서열 분석한 결과 이들의 90% 가량이 Proteobacterium 문에 속하였으며 다른 44속에 대부분 포함 되었다. 대표적인 44속에 대해 분자생물학적 동정을 한 후 다양한 외부환경(온도, pH, 염, 무기질소염) 조건에 대한 이 들의 생존성 범위를 조사한 결과, 넓은 온도(4oC~ 42oC)와 pH(4~10)범위에서 자랄 수 있는 탈질균 12 종을 최종 선발 하였다.
Acknowledgement
Authors thank Dr. Jin-Ho Kim and Dr. Chul-Man Choi for the ion chromatography analysis. This work is sup- ported by the grant from National Academy of Agricul-
tural Science, Rural Development Administration project PJ0067032011.
REFERENCES
1. Amann, R. I., W. Ludwig, and K. H. Schleifer. 1995.
Phylogenetic identification and in situ detection of individual microbial cells without cultivation. Microbiol. Rev. 59: 143- 169.
2. Cheneby, D., L. Philippot, A. Hartmann, C. Henault, and J.
Germon. 2000. 16S rDNA analysis for characterization of denitrifying bacteria isolated from three agricultural soils.
FEMS Microbiol. Ecol. 34: 121-128.
3. Colwell, R. K., E. A. Norse, D. Pimentel, F. E. Sharples, and D. Simberloff. 1985. Genetic engineering in agriculture.
Science 229: 111-112.
4. Colwell, R. R. and A. Huq. 1994. Environmental reservoir of Vibrio cholerae. The causative agent of cholera. Annals of the New York Academy of Sciences 740: 44-54.
5. Feng, S. and E. R. Tillier. 2007. A fast and flexible approach to oligonucleotide probe design for genomes and gene families. Bioinformatics 23: 1195-1202.
6. Fernandez-Arrojo, L., M. E. Guazzaroni, N. Lopez-Cortes, A. Beloqui, and M. Ferrer. 2010. Metagenomic era for bio- catalyst identification. Curr. Opin. Biotechnol. 21: 725-733.
7. Gao, H., Z. K. Yang, T. J. Gentry, L. Wu, C. W. Schadt, and J. Zhou. 2007. Microarray-based analysis of microbial com- munity RNAs by whole-community RNA amplification.
Appl. Environ. Microbiol. 73: 563-571.
8. Jones, C. M., B. Stres, M. Rosenquist, and S. Hallin. 2008.
Phylogenetic analysis of nitrite, nitric oxide, and nitrous oxide respiratory enzymes reveal a complex evolutionary history for denitrification. Mol. Biol. Evol. 25: 1955-1966.
9. Kim, H. and H. Kang. 2011. The Impacts of Excessive Nitrogen Additions on Enzyme Activities and Nutrient Leaching in Two Contrasting Forest Soils. J. Microbiol. 49:
369-375.
10. Lau, H. T., J. Faryna, and E. W. Triplett. 2006. Aquitalea magnusonii gen. nov., sp. nov., a novel Gram-negative bacterium isolated from a humic lake. Int. J. Syst. Evol.
Microbiol. 56: 867-871.
11. Lee, C. M., H. Y. Weon, Y. J. Kim, J. A. Son, S. H. Yoon, B.
S. Koo, and S. W. Kwon. 2009. Aquitalea denitrificans sp.
nov., isolated from a Korean wetland. Int. J. Syst. Evol.
Microbiol. 59: 1045-1048.
12. Ohta, H. and T. Hattori. 1983. Agromonas oligotrophica gen.
nov., sp. nov., a nitrogen-fixing oligotrophic bacterium.
Antonie Van Leeuwenhoek 49: 429-446.
13. Pace, N. R. 1997. A molecular view of microbial diversity and the biosphere. Science 276: 734-740.
14. Philippot, L., M. Cregut, D. Cheneby, M. Bressan, S. Dequiet, F. Martin-Laurent, L. Ranjard, and P. Lemanceau. 2008.
Effect of primary mild stresses on resilience and resistance of the nitrate reducer community to a subsequent severe
stress. FEMS Microbiol. Lett. 285: 51-57.
15. Poindexter, J. S. 1981. The caulobacters: ubiquitous unusual bacteria. Microbiol. Rev. 45: 123-179.
16. Schmidt, T. M., E. F. DeLong, and N. R. Pace. 1991.
Analysis of a marine picoplankton community by 16S rRNA gene cloning and sequencing. J. Bacteriol. 173: 4371-4378.
17. Singh, A., N. Singh Chauhan, H. V. Thulasiram, V. Taneja, and R. Sharma. 2010. Identification of two flavin monooxy- genases from an effluent treatment plant sludge metageno- mic library. Bioresour. Technol. 101: 8481-8484.
18. Staley, J. T. and A. Konopka. 1985. Measurement of in situ
activities of nonphotosynthetic microorganisms in aquatic and terrestrial habitats. Annu. Rev. Microbiol. 39: 321-346.
19. Steward, G. F., B. D. Jenkins, B. B. Ward, and J. P. Zehr.
2004. Development and testing of a DNA macroarray to assess nitrogenase (nifH) gene diversity. Appl. Environ.
Microbiol. 70: 1455-1465.
20. Tamura, K., J. Dudley, M. Nei, and S. Kumar. 2007. MEGA4:
Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol. Biol. Evol. 24: 1596-1599.
21. Whang, K. and T. Hattori. 1988. Oligotrophic bacteria from rendzina forest soil. Antonie Van Leeuwenhoek 54: 19-36.