국내에서 수집된 자두의 품종식별을 위한 SSR Profile 데이터베이스 구축
홍지화1* ・ 심은조1・ 박원흠2・ 소은희1
1농림축산식품부 국립종자원 종자검정연구센터, 2경북농업기술원 청도복숭아시험장
Construction of SSR Profile Database for Variety Identification of Plum Collected in Korea
Jee-Hwa Hong
1*, Eun-Jo Shim
1, Won-Heum Park
2, and Eun-Hee Soh
11
Seed Testing & Research Center, Korea Seed & Variety Service, Ministry of Agriculture, Food and Rural Affairs, Gimcheon 740-220, Korea
2
Cheongdo Peach Experiment Station, Gyeongsangbuk-do Agricultural Technology Administration, Cheongdo, 714-851, Korea
Abstract : This study was conducted to construct a DNA marker database for 38 plum varieties collected in Korea using simple sequence repeat (SSR) markers. A set of 61 SSR primer pairs was tested to select polymorphic SSR markers between 8 varieties.
Among the 61 primer pairs, 21 showed polymorphism, reproducibility and easy scoring. The genetic relationship between the 21 SSR markers and 38 varieties was analyzed. A total of 210 polymorphic amplified fragments were obtained with the 21 SSR markers. Three to seventeen SSR alleles were detected for each locus, with an average of 10.0 alleles per locus. Average polymorphism information content (PIC) was 0.758, with a range from 0.549 to 0.870. A total of 210 SSR marker loci were used to calculate Jaccard’s distance coefficients for cluster analysis by an unweighted pair-group method with arithmetical average (UPGMA). The genetic distance ranged from 0.06 to 1.00 in 38 varieties. Out of 38 plum varieties, 32 were identified using the 21 SSR markers. Therefore, these SSR markers may be employed to complement distinctness, uniformity, and stability (DUS) tests or as potential tools to solve seed disputes regarding plums.
Keywords : Plum, SSR marker, Variety identification, Genetic relationship, Plant variety protection
*Corresponding author (E-mail: [email protected], Tel: +82-54- 912-0230, Fax: +82-54-912-0227)
(Received on March 21, 2015. Revised on March 25, 2015.
Accepted on April 23, 2015.)
97
http://dx.doi.org/10.9787/KJBS.2015.47.2.97 Print ISSN: 0250-3360
Copyright ⓒ 2015 by the Korean Society of Breeding Science
This is an Open-Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
서 언
자두는 장미과(Rosaceae), Prunus속의 Prunophora아속에 속하고 유럽, 아시아, 미국에서 재배가 되어왔다(Weinberger 1975). 자두는 8개의 염색체가 있고 2배체(2n = 2x = 16)에 서 6배체(2n = 6x = 48)가 있는 배수성 작물이다(Esmenjaud
& Dirlewanger 2007). 유럽계 자두에는 P. domestica, P.
insititia L., P. cerasifera Ehrh.이 있고 이중 P. domestica는
자두 품종의 중요한 자원으로 2,000년 이상 재배되어 온 것으 로 알려져 있다(Esmenjaud and Dirlewanger 2007). 아시아에는 중국에서 유래되어 일본에서 재배된 일본계 자두(P.
salicina Lindl.)가 있고(Hedrick 1911), 북미계 자두에는 P.
Americana Marsh., P. hortulana Bailey, P. munsoniana
Wight & Hedr., P. angustifolia Marsh., P. maritime Marsh.가 있는 것으로 알려져 있다(Okie 1987).
국내에서 자두(P. salicina Lindl.)는 2008년도 3월 1일부 터 품종보호 대상작물로 지정되어 현재까지 품종보호 출원된 품종은 11품종이고 이중 품종보호 등록된 품종은 5품종이다.
자두의 생산수입판매신고 건수는 395건(2015년 3월 기준)으 로 과수작물 중 6위를 차지하고 있어 향후 품종진위 확인 등 종자분쟁이 예상되는 작물 중 하나이다. 따라서 자두 품종보 호 출원품종과 가장 유사한 대조품종의 선정, 종자분쟁을 해 결을 위해서는 품종을 식별할 수 있는 분자표지의 선발과 이 를 이용한 품종의 데이터베이스 구축이 필요하다.
분자표지를 활용한 자두의 국외연구 동향을 살펴보면 미국 에서는 randomly amplified polymorphic DNA (RAPD) 마 커를 이용하여 자두에 대한 유전적 유연관계를 분석한 이래 (Boonprakob et al. 2001), 스페인에서는 CT/AG enriched genomic library를 구축하여 35개의 simple sequence repeat (SSR) 마커를 개발하였고(Mnejja et al. 2004), 이란에서는 8 개의 SSR 마커를 이용하여 40품종에 대한 품종식별 연구를 수행하였다(Etehadpoor et al. 2012). 또한 브라질에서는 8개 SSR 마커를 이용하여 일본계 자두 47품종에 대한 품종식별 연구 결과를 보고한바 있다(Klabunde et al. 2014). 국외에서 는 몇몇 연구자들에 의해 분자표지를 이용한 자두 품종식별 연구가 수행되어 왔으나 국내에서는 국제 식물 신품종 보호 동맹(UPOV, International Union for the Protection of New Varieties of Plants)이 제안하고(UPOV 2010), 국외에서 자두 품종식별에 많이 활용되고 있는 SSR 마커를 이용한 다형성 분석과 데이터베이스 구축 등 관련 연구는 미비한 실정이다.
따라서 본 연구에서는 자두 품종보호 출원품종의 대조품종 선정, 품종진위와 관련된 종자분쟁 해결 등에 SSR 마커의 활 용 가능성을 알아보기 위하여 국내에서 수집된 자두 품종들 을 대상으로 품종식별에 적합한 SSR 마커를 선발하고 이를 이용한 품종별 DNA 프로파일 데이터베이스 구축과 품종간 유연관계 분석을 실시하였다.
재료 및 방법
공시재료 및 DNA 추출
경북농업기술원 청도복숭아시험장에서 수집된 38품종의 잎을 채취하여 분석을 위한 재료로 활용하였다(Table 1). 공 시품종의 DNA는 NucleoSpin®PlantⅡ (Macherey-Nagel Cat.
740 770.250) 키트를 이용하여 추출하였다. 추출된 DNA는 분광광도계(NanoDrop2000, Thermo Scientific, USA)를 활 용하여 DNA 농도를 확인한 후 μL당 20 ng의 농도로 맞추어 PCR 분석에 사용하였다.
SSR 마커 선발 및 데이터베이스 구축
자두 품종식별을 위한 마커를 선발하기 위하여 국외에서 개발된 61개의 SSR 마커를 이용하여 ‘대석조생(Oishiwase)’,
‘포모사(Formosa)’, ‘자봉(Shihou)’, ‘귀양(Kiyou)’, ‘산타로 사(Santa Rosa)’, ‘퍼플퀸(Purple Queen)’, ‘하니레드(Honey Red)’, ‘홍료젠(Beniryozen)’ 8품종을 대상으로 다형성을 보
이는 마커를 선발하고자 하였다. PCR (Polymerase Chain Reaction) 반응산물은 자두 DNA 20 ng, 0.5 μM의 SSR primer, 2 ㎕ dNTP mixture (2.5 mM), Taq polymerase 1 units, 2.5 ㎕의 10 x PCR buffer (50 mM KCl, 20 mM Tris- HCl, pH 8.0, 2 mM MgCl2) (Genet Bio, Korea)에 증류수 를 첨가하여 총 반응액을 25 ㎕로 맞추었다. PCR (GeneAmp®
PCR System 9700)은 40회 실시하였고, pre-denaturation은 94℃에서 10분, denaturation은 94℃에서 30초, annealing은 50∼60℃에서 30초, extension은 72℃에서 45초, final-extension 은 72℃에서 10분간 수행하였다. PCR 증폭 산물은 QIAxcel Advanced System (QIAGEN)을 이용하여 전기영동 하였고 QIAxcel ScreenGel을 활용하여 각 품종별 대립유전자의 차 이를 분석하여 다형성을 보이는 마커를 선발하였다.
다형성을 나타내는 마커 중 SSR 프로파일 데이터베이스 구축에 활용할 마커를 선정한 후 프라이머의 정방향에 FAM, VIC, NED, PET 중 한가지로 형광 표지하여 자두 38품종을 PCR 하였다. 4 ㎕의 PCR 증폭산물을 2% 아가로스 젤에서 전기영동하여 증폭 여부를 확인한 후 증류수 220 ㎕에 PCR 산물을 증폭량에 따라 1∼3 ㎕씩 첨가하였다. 희석된 PCR 증폭 산물과 Hi-Di formamide와 size marker (LIZ500 size standard)를 첨가하여 94℃에서 2분간 denaturation 시킨 후 자동염기서열 분석장치(3130XL Genetic Analyzer, Applied Biosystems, USA)를 활용하여 전기영동 하였고, GeneMapper 3.7 프로그램(Applied Biosystems, USA)을 이용하여 대립유 전자 크기를 분석하였다.
다형성 지수 및 유사도 분석
자동염기서열 분석장치를 통해 분석된 대립유전자의 유무에 따라 피크가 있으면 ‘1’, 없으면 ‘0’으로 기록하여 엑셀파일에 품종별 SSR 프로파일을 대립유전자 크기 별로 정리하여 데이 터베이스를 구축하였다. SSR 마커의 다형성 지수를 조사하기 위하여 Anderson et al. (1993)이 보고한 공식을 이용하여 polymorphism information content (PIC) 값을 산출하였다.
NTSYS-pc (version 2.21 m)(Rohlf 2000) 프로그램의 Jaccard (Sneath & Sokal 1973)방법에 따라 유전적 유사도 값을 계산 한 후 unweighted pair-group method with arithmetical average (UPGMA)(Sneath and Sokal 1973) 방법을 통해 유 연관계를 분석하였다. 품종식별을 위한 최소 마커 선정을 위 하여 Mantel test (Mantel 1967) 분석을 통해 상관계수 값을 산출하였다.
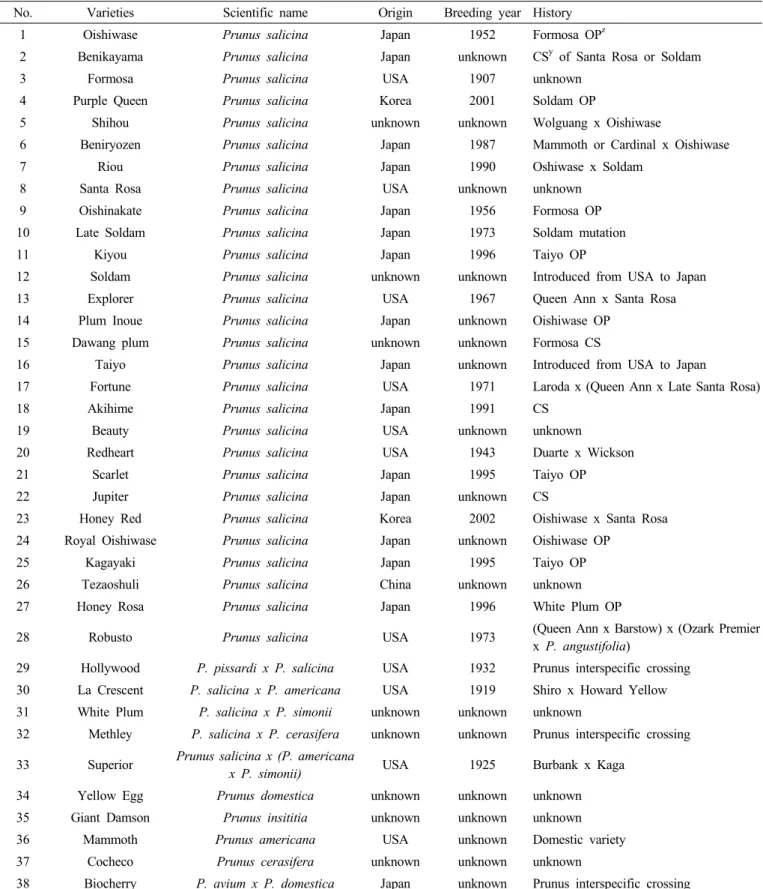
Table 1. The thirty-eight plum varieties used for construction of simple sequence repeat profile database.
No. Varieties Scientific name Origin Breeding year History
1 Oishiwase Prunus salicina Japan 1952 Formosa OPz
2 Benikayama Prunus salicina Japan unknown CSy of Santa Rosa or Soldam
3 Formosa Prunus salicina USA 1907 unknown
4 Purple Queen Prunus salicina Korea 2001 Soldam OP
5 Shihou Prunus salicina unknown unknown Wolguang x Oishiwase
6 Beniryozen Prunus salicina Japan 1987 Mammoth or Cardinal x Oishiwase
7 Riou Prunus salicina Japan 1990 Oshiwase x Soldam
8 Santa Rosa Prunus salicina USA unknown unknown
9 Oishinakate Prunus salicina Japan 1956 Formosa OP
10 Late Soldam Prunus salicina Japan 1973 Soldam mutation
11 Kiyou Prunus salicina Japan 1996 Taiyo OP
12 Soldam Prunus salicina unknown unknown Introduced from USA to Japan
13 Explorer Prunus salicina USA 1967 Queen Ann x Santa Rosa
14 Plum Inoue Prunus salicina Japan unknown Oishiwase OP
15 Dawang plum Prunus salicina unknown unknown Formosa CS
16 Taiyo Prunus salicina Japan unknown Introduced from USA to Japan
17 Fortune Prunus salicina USA 1971 Laroda x (Queen Ann x Late Santa Rosa)
18 Akihime Prunus salicina Japan 1991 CS
19 Beauty Prunus salicina USA unknown unknown
20 Redheart Prunus salicina USA 1943 Duarte x Wickson
21 Scarlet Prunus salicina Japan 1995 Taiyo OP
22 Jupiter Prunus salicina Japan unknown CS
23 Honey Red Prunus salicina Korea 2002 Oishiwase x Santa Rosa
24 Royal Oishiwase Prunus salicina Japan unknown Oishiwase OP
25 Kagayaki Prunus salicina Japan 1995 Taiyo OP
26 Tezaoshuli Prunus salicina China unknown unknown
27 Honey Rosa Prunus salicina Japan 1996 White Plum OP
28 Robusto Prunus salicina USA 1973 (Queen Ann x Barstow) x (Ozark Premier
x P. angustifolia)
29 Hollywood P. pissardi x P. salicina USA 1932 Prunus interspecific crossing 30 La Crescent P. salicina x P. americana USA 1919 Shiro x Howard Yellow 31 White Plum P. salicina x P. simonii unknown unknown unknown
32 Methley P. salicina x P. cerasifera unknown unknown Prunus interspecific crossing 33 Superior Prunus salicina x (P. americana
x P. simonii) USA 1925 Burbank x Kaga
34 Yellow Egg Prunus domestica unknown unknown unknown
35 Giant Damson Prunus insititia unknown unknown unknown
36 Mammoth Prunus americana USA unknown Domestic variety
37 Cocheco Prunus cerasifera unknown unknown unknown
38 Biocherry P. avium x P. domestica Japan unknown Prunus interspecific crossing
zOP : Open pollination
yCS : Chance seedling
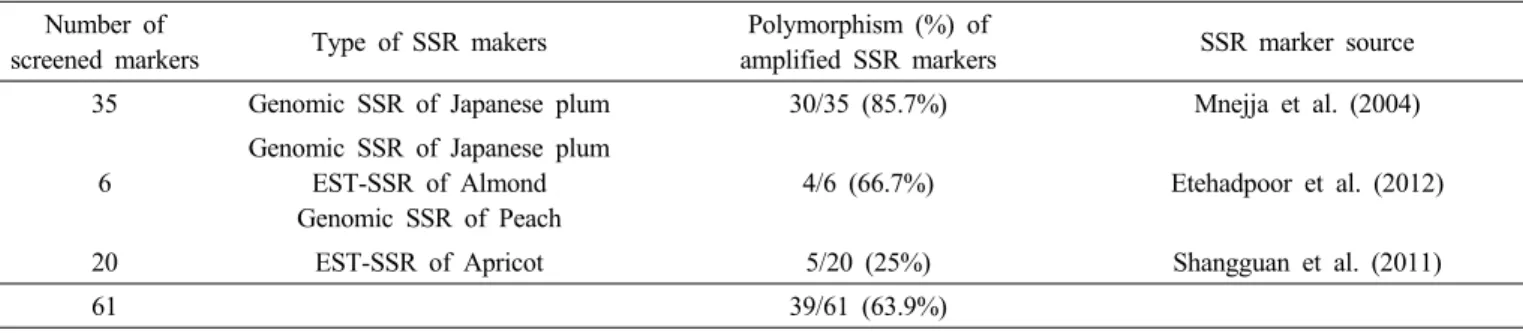
Table 2. Simple sequence repeat markers screened for identifying plum varieties and polymorphism of amplified markers.
Number of
screened markers Type of SSR makers Polymorphism (%) of
amplified SSR markers SSR marker source
35 Genomic SSR of Japanese plum 30/35 (85.7%) Mnejja et al. (2004)
6
Genomic SSR of Japanese plum EST-SSR of Almond Genomic SSR of Peach
4/6 (66.7%) Etehadpoor et al. (2012)
20 EST-SSR of Apricot 5/20 (25%) Shangguan et al. (2011)
61 39/61 (63.9%)
Fig. 1. Four simple sequence repeat markers showing polymorphism among 8 plum varieties. The PCR products were separated using a QIAxcel Advanced System (QIAGEN). Lane 1 : Oishiwase, 2 : Formosa, 3 : Shihou, 4 : Kiyou, 5 : Santa Rosa, 6 : Purple Queen, 7 : Honey Red, 8 : Beniryozen.
결과 및 고찰
SSR 마커 선발 및 데이터베이스 구축
자두 품종식별에 적합한 SSR 마커를 선발하기 위해서 자 두뿐만 아니라 Prunus속의 아몬드, 복숭아, 살구로부터 개발 된 SSR 마커를 분석에 활용하였다. 자두에서 유래된 35개 SSR (Mnejja et al. 2004), 자두, 아몬드, 복숭아로부터 유래 된 6개 SSR (Etehadpoor et al. 2012), 살구에서 유래된 20개 SSR 마커(Shangguan et al. 2011), 총 61개 SSR 마커를 이용하 여 ‘대석조생(Oishiwase)’, ‘포모사(Formosa)’, ‘자봉(Shihou)’,
‘귀양(Kiyou)’, ‘산타로사(Santa Rosa)’, ‘퍼플퀸(Purple Queen)’,
‘하니레드(Honey Red)’, ‘홍료젠(Beniryozen)’ 8품종을 대상 으로 다형성을 분석 한 결과 스페인에서 개발된 자두 35개의 SSR 마커 중 30개 마커, 이란에서 선발된 총 6개의 SSR 마 커 중 4개 마커, 중국에서 개발된 살구 20개의 SSR 마커 중 5개 마커가 공시품종에서 다형성을 나타내었다. 총 61개 마 커 중 39개 마커가 다형성을 보여 다형성 비율은 63.9%였고 마커 유래별 다형성 비율은 25%에서 85.7%까지 나타났다 (Table 2). QIAxcel ScreenGel 분석에서 다형성을 보이는 39 개 SSR 마커 중 반복 실험간 뚜렷한 재현성을 보이며 대립유전 자의 판독이 용이한 ‘CPSCT022’, ‘CPSCT041’, ‘BPPCT001’,
‘EST-SSR2’ 등 21개의 마커를 최종 선발하였다(Fig. 1). 21 개 마커 중 17개는 자두에서 개발된 SSR 마커(Mnejja et al.
2004)이고, 2개는 각각 아몬드 및 복숭아로부터 유래된 마커 (Etehadpoor et al. 2012)이며, 나머지 2개는 살구에서 유래 된 마커(Shangguan et al. 2011)인바 자두와 동일한 Prunus 속이 주류이나 종이 다른 아몬드, 복숭아, 살구에서 유래된 SSR 마커가 본 연구에서 공시한 자두 품종에서 안정적으로 증폭되고 품종간 다형성을 나타냄을 확인할 수 있었다(Table 3). 따라서 최종적으로 선발된 21개의 SSR 마커를 이용하여 국내에서 수집된 자두 38품종의 DNA 프로파일 데이터베이
스를 구축하였다. 21개 SSR 마커에 의해 분석된 대립유전자 의 수는 총 210개였고, 평균 대립유전자의 수는 10개로 나타 났으며, 마커 당 대립유전자수는 ‘EST-SSR26’ 마커가 3개로 가장 적었고 ‘CPSCT027’ 마커가 17개로 가장 많았다(Table 3). 스페인에서는 자두 8품종에 대하여 27개의 SSR 마커로 다형성 분석하였을 때 마커 당 대립유전자 수는 2∼9개이고, 평균 대립유전자의 개수는 5.7개(Mnejja et al. 2004)임을 보 고하였으나, 브라질에서는 8개 SSR 마커로 47품종을 분석하 였을 때 대립유전자 수는 8∼21개이고, 평균 13개의 대립유 전자가 나타났고(Klabunde et al. 2014), 이란에서는 8개 SSR 마커로 4품종을 분석하였을 때 대립유전자 수가 4∼12 개임(Etehadpoor et al. 2012)을 보고한 바 있어 마커에 따른 대립유전자 수는 공시품종의 수와 품종의 다양성 정도에 따 라 차이가 남을 알 수 있었다. 본 연구에 활용된 자두 품종 중 일본계 자두(P. salicina Lindl.)는 2배체이고, 유럽계 자두 중
P. cerasifera는 2배체이나 P. domestica, P. insititia은 6배
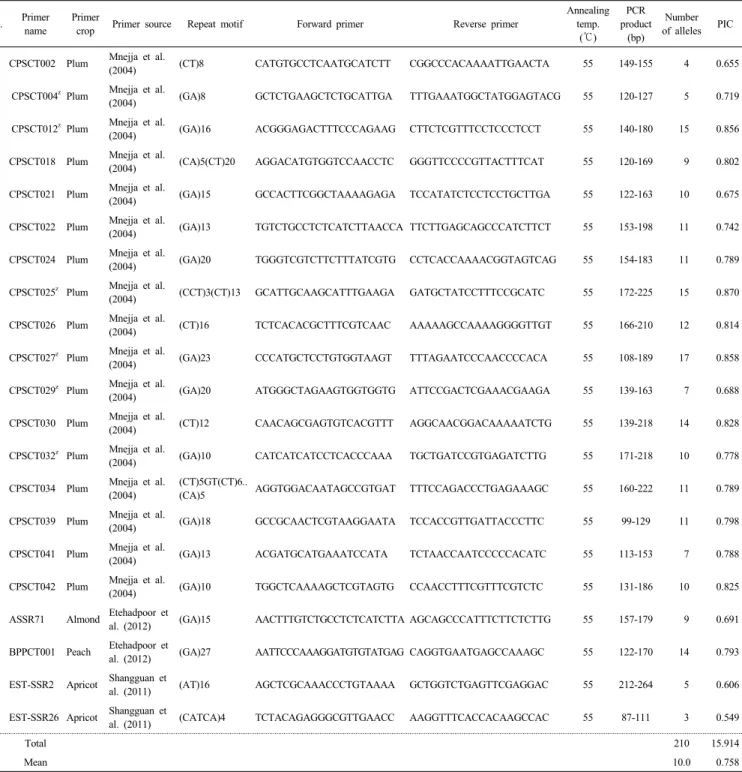
체로 알려져 있어(Esmenjaud & Dirlewanger 2007) 공시품 종 마다 배수성이 달라서 대립유전자 수가 많이 나타난 것으 로 추정된다(Silva et al. 2012). Ming et al. (2006)은 배수성 작물에서 종간교잡에 의해 품종이 육성될 경우 배수체로 인Table 3. The effective 21 simple sequence repeat markers selected for identification of plum varieties.
No. Primer name
Primer
crop Primer source Repeat motif Forward primer Reverse primer
Annealing temp.
(℃)
PCR product
(bp)
Number of alleles PIC 1 CPSCT002 Plum Mnejja et al.
(2004) (CT)8 CATGTGCCTCAATGCATCTT CGGCCCACAAAATTGAACTA 55 149-155 4 0.655
2 CPSCT004zPlum Mnejja et al.
(2004) (GA)8 GCTCTGAAGCTCTGCATTGA TTTGAAATGGCTATGGAGTACG 55 120-127 5 0.719 3 CPSCT012zPlum Mnejja et al.
(2004) (GA)16 ACGGGAGACTTTCCCAGAAG CTTCTCGTTTCCTCCCTCCT 55 140-180 15 0.856 4 CPSCT018 Plum Mnejja et al.
(2004) (CA)5(CT)20 AGGACATGTGGTCCAACCTC GGGTTCCCCGTTACTTTCAT 55 120-169 9 0.802 5 CPSCT021 Plum Mnejja et al.
(2004) (GA)15 GCCACTTCGGCTAAAAGAGA TCCATATCTCCTCCTGCTTGA 55 122-163 10 0.675 6 CPSCT022 Plum Mnejja et al.
(2004) (GA)13 TGTCTGCCTCTCATCTTAACCA TTCTTGAGCAGCCCATCTTCT 55 153-198 11 0.742 7 CPSCT024 Plum Mnejja et al.
(2004) (GA)20 TGGGTCGTCTTCTTTATCGTG CCTCACCAAAACGGTAGTCAG 55 154-183 11 0.789 8 CPSCT025z Plum Mnejja et al.
(2004) (CCT)3(CT)13 GCATTGCAAGCATTTGAAGA GATGCTATCCTTTCCGCATC 55 172-225 15 0.870 9 CPSCT026 Plum Mnejja et al.
(2004) (CT)16 TCTCACACGCTTTCGTCAAC AAAAAGCCAAAAGGGGTTGT 55 166-210 12 0.814 10 CPSCT027z Plum Mnejja et al.
(2004) (GA)23 CCCATGCTCCTGTGGTAAGT TTTAGAATCCCAACCCCACA 55 108-189 17 0.858 11 CPSCT029z Plum Mnejja et al.
(2004) (GA)20 ATGGGCTAGAAGTGGTGGTG ATTCCGACTCGAAACGAAGA 55 139-163 7 0.688 12 CPSCT030 Plum Mnejja et al.
(2004) (CT)12 CAACAGCGAGTGTCACGTTT AGGCAACGGACAAAAATCTG 55 139-218 14 0.828 13 CPSCT032z Plum Mnejja et al.
(2004) (GA)10 CATCATCATCCTCACCCAAA TGCTGATCCGTGAGATCTTG 55 171-218 10 0.778 14 CPSCT034 Plum Mnejja et al.
(2004)
(CT)5GT(CT)6..
(CA)5 AGGTGGACAATAGCCGTGAT TTTCCAGACCCTGAGAAAGC 55 160-222 11 0.789 15 CPSCT039 Plum Mnejja et al.
(2004) (GA)18 GCCGCAACTCGTAAGGAATA TCCACCGTTGATTACCCTTC 55 99-129 11 0.798 16 CPSCT041 Plum Mnejja et al.
(2004) (GA)13 ACGATGCATGAAATCCATA TCTAACCAATCCCCCACATC 55 113-153 7 0.788
17 CPSCT042 Plum Mnejja et al.
(2004) (GA)10 TGGCTCAAAAGCTCGTAGTG CCAACCTTTCGTTTCGTCTC 55 131-186 10 0.825 18 ASSR71 Almond Etehadpoor et
al. (2012) (GA)15 AACTTTGTCTGCCTCTCATCTTA AGCAGCCCATTTCTTCTCTTG 55 157-179 9 0.691 19 BPPCT001 Peach Etehadpoor et
al. (2012) (GA)27 AATTCCCAAAGGATGTGTATGAG CAGGTGAATGAGCCAAAGC 55 122-170 14 0.793 20 EST-SSR2 Apricot Shangguan et
al. (2011) (AT)16 AGCTCGCAAACCCTGTAAAA GCTGGTCTGAGTTCGAGGAC 55 212-264 5 0.606 21 EST-SSR26 Apricot Shangguan et
al. (2011) (CATCA)4 TCTACAGAGGGCGTTGAACC AAGGTTTCACCACAAGCCAC 55 87-111 3 0.549
Total 210 15.914
Mean 10.0 0.758
zMinimum markers
하여 동일한 유전자좌에 서로 다른 대립유전자를 가지는 이 형접합체(heterozygote)를 나타낸다고 보고한 바 있다.
한편, 공시품종 간의 다형성 정도를 나타내는 PIC 값은 0.549∼0.870이였고 평균 0.758을 나타내었다. ‘EST-SSR26’
마커에서 0.549를 나타내었고, ‘CPSCT025’ 마커에서 0.870 으로 나타나 대립유전자가 많이 분석된 마커에서 PIC 값이 높은 경향을 나타내었다. Mnejja et al. (2004)에 의하면 자두 8품종을 27개의 SSR 마커로 분석하였을 때 다형성 정도를
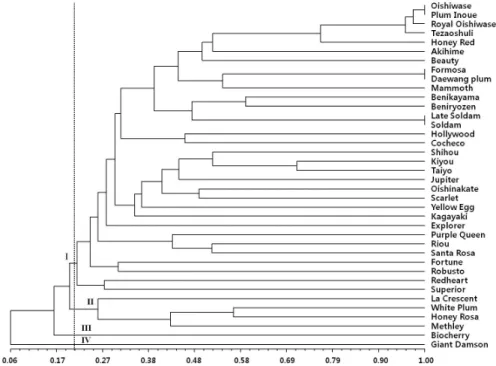
Fig. 2. Genetic relationship of 38 plum varieties using 21 simple sequence repeat markers. The scale at the bottom is Jaccard’s coefficient of similarity. The dotted line refer to a similarity level of 0.21.
나타내는 Discrimination Power (DP) 값은 0.74이였고, Etehadpoor et al. (2012)는 PIC값이 0.69이였음을 보고하였으 며, Klabunde et al. (2014)는 0.803으로 보고한 바 있어 본 연구의 PIC 값은 Mnejja et al. (2004) 및 Etehadpoor et al.
(2012)의 결과보다 높은 값을 나타내었으나, Klabunde et al.
(2014)의 결과보다는 낮게 나타났다.
자두의 품종식별을 위한 최소 마커를 선정하기 위하여 21 개 마커 중 PIC 값이 높으면서 공시품종이 식별 가능한 6개 SSR 마커(CPSCT004, CPSCT012, CPSCT025, CPSCT027, CPSCT029, CPSCT032)를 선정하였다. 최종 선발된 21개 마커 조합과 6개 마커 조합의 상관관계(Mantel test) 분석 결 과 상관계수(r) 값이 0.821로 높게 나타나 향후 자두 품종식 별에 6개의 최소 마커 조합을 우선 적용한다면 분석에 소요되 는 시간과 비용을 절감할 수 있을 것으로 사료된다.
품종간 유연관계 분석
최종 선발된 21개 SSR 마커를 이용하여 자두 38품종에 대한 유연관계를 분석하였다(Fig. 2). 공시품종의 유사도 지 수는 0.06∼1.00의 범위로 나타났고, 38품종 중 ‘대석조생 (Oishiwase)’과 ‘플럼정상(Plum Inoue)’, ‘포모사(Formosa)’
와 ‘대왕자두(Daewang plum)’, ‘솔담(Soldam)’과 ‘레이트
솔담(Late Soldam)’ 을 제외한 32품종이 21개 마커에 의해 식별되는 것으로 나타났다. 마커로 식별되지 않은 품종들의 육성내역을 살펴보면 1) ‘플럼정상(Plum Inoue)’은 ‘대석조 생(Oishiwase)’의 자연교잡 실생으로 육성되었고, 2) ‘대왕자 두(Daewang plum)’는 ‘포모사(Formosa)’의 우연 실생으로 육 성되었으며, 3) ‘레이트 솔담(Late Soldam)’은 ‘솔담(Soldam)’
의 돌연변이로 육성된 것으로 알려져 있다. 1) ‘플럼정상 (Plum Inoue)’은 ‘대석조생(Oishiwase)’을 품종 육성 시에 모본으로 활용하였기 때문에 유사도가 높게 나타난 것으로 추정되고 2) ‘대왕자두(Daewang plum)’도 ‘포모사(Formosa)’
를 품종 육성 시에 모본으로 활용하였기 때문에 두 품종간 유 사도가 높게 나타난 것으로 추정된다. Yamamoto et al.
(2003)는 복숭아에서 우연 실생으로 육성된 품종에 대하여 SSR 마커로 분석하였을 때 모본과 육성된 품종간에 대립유 전자의 차이가 없음을 보고하였다. Hong et al. (2015)은 분 자표지로 식별되지 않는 품종에 대하여 형태적 특성조사를 수행하여 분자표지와 형태적 특성의 상관관계를 분석한 바 있는데, 앞으로 분자표지로 식별이 되지 않은 자두 품종들에 대해서는 형태적 특성검정을 통하여 적정 분자표지의 수를 검 토해야 할 것으로 사료된다. 3) ‘레이트 솔담(Late Soldam)’은
‘솔담(Soldam)’의 돌연변이로 육성되었기 때문에 두 품종간 유
전적 유사도가 100%로 나타난 것으로 추정된다. Yamamoto et al. (2003)는 아조변이로 육성된 품종의 경우 동일한 대립유전 자형을 나타낸다고 보고하였고, Hong et al. (2013)은 사과에 서 원품종(Initial variety)과 돌연변이로 육성된 품종 간에 SSR 마커로 식별이 가능하다고 하였으며, 복숭아의 경우 원 품종과 돌연변이 품종간에 식별되는 경우와 식별이 되지 않는 경우가 있다고 보고한 바 있다(Hong et al. 2013). 기존의 연 구 결과를 살펴볼 때 원품종과 원품종으로부터 유래된 품종간 의 품종식별은 품종 육성 시 변이의 정도에 따라 달라질 것으 로 추정되나, 품종간의 육성내역이 명확하다면 유래관계를 증 명할 수 있는 DNA 마커 개발과 정확한 유전적 양상을 알아보 기 위한 연구가 추가적으로 수행 되어야 할 것으로 사료된다.
공시품종간의 유연관계를 살펴보면 유사도 지수 0.21을 기 준으로 할 때 4개의 그룹으로 구분되었다. 4개의 그룹이 자두 의 유래된 지역별 계통과 정확하게 일치되지는 않았으나 대 체적으로 그룹 I에는 일본계 자두가 주로 속하였고 그룹 II에 는 일본계 자두와 교배된 교잡종 품종들이 주로 분포하였으 며, 그룹 III에는 유럽계 자두와 교잡된 교잡종, 그룹 IV에는 유럽계 자두가 분포하였다. Etehadpoor et al. (2012)이 보고 한 연구 결과에 따르면, 자두 품종이 수집된 지역의 품종별로 그룹화가 되지 않았다고 보고한 바 있는데 본 연구에서도 비 슷한 양상을 나타내었다. 일본계 자두들이 속해있는 그룹 I의 품종 중 ‘할리우드(Hollywood)’는 일본계 자두와 교잡종으로 육성된 품종이고, ‘맘모스(Mammoth)’는 미국계 자두이며,
‘코체코(Cocheco)’ 및 ‘옐로우에그(Yellow Egg)’는 유럽계 자두, ‘슈페리어(Superior)’는 일본계 자두와 미국계 자두의 교배로 육성된 품종으로 알려져 있는데 향후 분자 마커의 수 를 증가하거나, 육성내역이나 유래관계에 대한 조사, 형태적 특성검정을 통한 품종간의 거리를 면밀히 조사한다면 계통별 불일치가 되는 부분을 이해할 수 있을 것으로 사료된다. ‘하 니로사(Honey Rosa)’는 일본계 자두로 알려져 있지만 일본 계 자두와 교잡으로 육성된 품종들이 분포하는 그룹 II에 속하 였는데 ‘하니로사(Honey Rosa)’가 ‘화이트플럼(White Plum)’
의 자연 교잡실생으로 육성되었기에 ‘화이트플럼(White Plum)’
과 같은 그룹에 속한 것으로 추정된다. 그룹 III에 속하는 ‘바 이오체리(Biocherry)’는 유럽계 자두와 교잡으로 육성된 품종 이고, 그룹 IV에 속하는 ‘자이언트댐슨(Giant Damson)’은 유 럽계 자두로 알려져 있어 다른 품종들과 유전적 거리가 멀게 나타난 것으로 추정된다. 그룹 I의 공시품종 중 ‘하니레드 (Honey Red)’와 ‘대석조생(Oishiwase)’의 유사도는 0.77로
다른 품종에 비해 유사도가 높게 나타났는데 ‘대석조생 (Oishiwase)’이 ‘하니레드(Honey Red)’의 모친으로 활용되 었기 때문인 것으로 추정된다.
UPOV의 실무기술위원회 중의 하나인 분자생물학 및 생화 학 실무작업반회의(BMT, Working Group on Biochemical and Molecular Techniques, and DNA-Profiling in Particular) 에서는 품종보호 제도에 분자표지 기술의 개발과 활용을 위 한 논의를 통해 분자표지를 구별성(Distinctness, D), 균일성 (Uniformity, U), 안정성(Stability, S) 심사에 직접적으로 활 용하는 것은 인정하지 않고 있으나 DNA 마커 선발 및 데이 터베이스 구축에 대한 지침서 작성(UPOV 2010) 및 DUS 심 사에 DNA 마커의 활용 방안을 제시한 바 있고(UPOV 2011), 최근에는 TGP (Technical Guidelines Protocol) 15 문서를 추가 제정함으로써 형질 연관 마커의 활용과 기존품종 관리 에 분자표지와 형태적 특성의 병합에 관한 적용 사례를 구체 적으로 소개하고 있는 실정이다(UPOV 2013).
본 연구에서는 UPOV 유전자 분석 지침서를 기반으로 자 두의 SSR 마커를 선발하고 품종별 DNA 프로파일 데이터베 이스를 구축하였다. 향후 공시품종에 대한 특성조사를 통해 SSR 마커와 형태적 특성의 상관관계가 구체적으로 밝혀진다 면 본 연구결과가 자두 품종보호 출원품종의 대조품종 선정 과 구별성, 균일성, 안정성 검정, 분쟁종자에 대한 품종진위 확인 등에 유용하게 활용될 것으로 사료된다.
적 요
국내에서 수집된 자두 38품종의 식별을 위하여 SSR 마커 를 이용하여 품종별 SSR 프로파일 데이터베이스를 구축하였 다. 자두 품종의 식별에 적합한 마커를 선정하기 위하여 8개 품종을 대상으로 총 61개의 마커를 분석하였다. 8개 품종간 에 높은 다형성과 재현성을 나타내고, 밴드패턴이 선명한 21 개의 마커를 선발하여 공시된 38품종을 분석하였을 때 총 210개의 대립유전자가 분석되었다. 대립유전자의 수의 분포 는 3∼17개를 나타내었고, 마커당 평균 대립유전자의 수는 10개로 분석되었다. PIC 값은 0.549∼0.870의 범위에 속하 였으며 평균값은 0.758로 나타났다. 210개의 대립유전자를 Jaccard 방법에 의해 유사도를 산출하고 비가중 산술방식에 의해 집괴 분석한 결과 공시품종의 유전적 거리는 0.06∼
1.00의 범위를 나타내었고 21개 SSR 마커에 의해 38품종 중 32품종이 식별되었다. 본 결과는 자두 출원품종의 신품종 보
호 심사를 위한 유전자 분석 자료, 종자분쟁 발생 시 해결도 구로 유용하게 활용될 것으로 사료된다.
REFERENCES
1. Anderson JA, Churchill GA, Autrigue JE, Tanksley SD.
1993. Optimizing parental selection for genetic linkage maps. Genome 36: 181-186.
2. Boonprakob U, Byrne DH, Graham CJ, Okie WR, Beckman T, Smith BR. 2001. Genetic relationships among cultivated diploid plums and their progenitors as determined by RAPD markers. J. Am. Soc. Hortic. Sci. 126: 451-461.
3. Esmenjaud D, Dirlewanger E. 2007. Genome mapping and molecular breeding in plants. Volume 4, Fruit and Nuts, C. Kole (Ed.). pp.119-135.
4. Etehadpoor M, Fatahi Moghadam MR, Zamani Z. 2012.
Genetic diversity of wild and commercial genotypes of plum using SSR molecular marker. Iranian J. Horticul.
Sci. 43: 199-210.
5. Hedrick UP. 1911. The Plums of New York. Rep. NY State Agri. Exp. Stn. pp.611.
6. Hong JH, Kwon YS, Choi KJ. 2013. Analysis of genetic relationship of apple varieties using microsatellite markers.
J. Life Sci. 23: 721-727.
7. Hong JH, Yi SI, Kwon YS, Kim Y, Choi KJ. 2013.
Genetic diversity analysis of peach [Prunus persica (L.) Batsch] varieties using SSR markers. Korean J. Breed.
Sci. 45: 201-211.
8. Hong JH, Kwon YS, Mishra RK, Kim DH. 2015.
Construction of EST-SSR databases for effective cultivar identification and their applicability to complement for lettuce (Lactuca sativa L.) distinctness test. Amer. J.
Plant Sci. 6: 113-125.
9. Klabunde GHF, Dalbó MA, Nodari RO. 2014. DNA fingerprinting of Japanese plum (Prunus salicina) cultivars based on microsatellite markers. Crop Breed. Appl.
Biotechnol. 14: 139-145.
10. Mantel N. 1967. The detection of disease clustering and a generalized regression approach. Cancer Res. 27: 209-220.
11. Ming R, Moore PH, Wu K, D’Hont A, Glazmann JC, Tew TL, Mirkov TE, da Silva J, Jifon J, Rai M. Schnell
RJ, Brumbley SM, Lakshmanan P, Comstock JC, Paterson AH. 2006. Sugarcane improvement through breeding and biotechnology. Plant Breed. Rev. 27: 15-118.
12. Mnejja M, Garcia-Mas J, Howad W, Badenes ML, ArÚs P. 2004. Simple-sequence repeat (SSR) markers of Japanese plum (Prunus salicina Lindl.) are highly polymorphic and transferable to peach and almond. Mol. Ecol. Notes. 4:
163-166.
13. Okie WR. 1987. Plum rootstocks. In: Rom RC, Carlson RF (eds) Rootstocks for Fruit Crops. John Wiley and sons, New-York, USA, pp.321-360.
14. Rohlf FJ. 2000. NTSYSpc. Numerical taxonomy and multivariate analysis system-version 2.10b. Applied Biostatistics Inc, New York.
15. Shangguan LF, Li XY, Ning N, Wang YZ, Zhang Z, Fang JG. 2011. Development of EST-SSR markers in apricot. Acta Horticulturae Sinica. 38: 43-54.
16. Silva DC, Duarte-Filho LSC, Santos JM, Barbosa GVS.
2012. DNA fingerprinting based on simple sequence repeat (SSR) markers in sugarcane clones from the breeding program RIDESA. Afr. J. Biotechnol. 11:
4722-4728.
17. Sneath PHA, Sokal RR. 1973. Numerical taxonomy: The principles and practice of numerical classification. W. H.
Freeman, San Francisco.
18. UPOV. 2010. INF/17/1 Guidelines for DNA-profiling:
Molecular marker selection and database construction (“BMT guidelines”), Geneva, Switzerland.
19. UPOV. 2011. INF/18/1 Possible use of molecular markers in the examination of distinctness, uniformity and stability (DUS), Geneva, Switzerland.
20. UPOV. 2013. TGP/15/1 Guidance on the use of bio- chemical and molecular markers in the examination of distinctness, uniformity and stability (DUS), Geneva, Switzerland.
21. Weinberger JH. 1975. Plums. In: J.Janick and J.N.
Moore(eds.). Advances in fruit breeding. Purdue Univ.
Press, West Lafayette, Ind. pp.336-347.
22. Yamamoto T, Mochida K, Imai T, Haji T, Yaegaki H, Yamaguchi M, Matsuta N, Ogiwara I, Hayashi T. 2003.
Parentage analysis in Japanese peaches using SSR markers.
Breed. Sci. 53: 35-40.