낙동정맥 애미랑재 지역의 식생구조 분석
이수동∙이승주*
경남과학기술대학교 조경학과∙*한국농어촌공사
Analysing Vegetation Structure of Aemirang Hill in the Nakdongjeongmaek, Korea
Lee, Soodong∙Lee, Seungjoo*
Dept. of Landscape Architecture, Gyeongnam National University Science and Technology
*
Korea Rural Community Corporation
ABSTRACT : This paper is to contribute for management and restoration as a basic study of vegetation structure in Aemirang Hill, Nakdongjeongmaek. To verify the characteristics of vegetation structure, we set up 25 plots(100㎡). The result of community analysis was used by TWINSPAN classification. The communities were classified 7 types i.e. Quercus variabilis community, Quercus mongolica community, Pinus densiflora community, Pinus densiflora-Quercus mongolica community, Q. mongolica-Quercus variabilis community, Quercus mongolica-Betula davurica community, Larix leptolepis community. Pinus densiflora community and Pinus densiflora-Quercus mongolica community are progressively transforming a Quercus mongolica community that is called Pinus densiflora next succession stage. Whereas the communities that has dominant Quercus variabilis and Quercus mongolica did not appear a next generation succession tree such as Carpinus laxiflora, Carpinus cordata in the under story and shrub layer. Therefore, Aemirang Hill's ecological succession was to maintain the status condition. Aemirang Hill's diversity index is 0.5738 to 0.8906. It were analyzed that diversity index was lower than other places.
Key words : vegetation structure, basic study, aemirang hill, ecological succession
1)I. 서 론
한반도의 산맥체계는 일제강점기때 한반도 지하자원 을 조사하는 과정에서 생겨난 산물로 1900년대 초 일본 지질학자인 고토 분지로(小藤文次郞)가 조사한 지질구 조에 바탕을 두고 ‘창지개명(倉地改名)’한 산맥이름을 그대로 쓰고 있다. 하지만 1980년대 초부터 일반인에게 조선후기에 편찬된 지리서인 ‘산경표(山經表)’의 존재가 알려지면서 백두대간을 중심으로 한 전통적 지리 인식체 계에 대한 국민적 관심이 높아지기 시작하였다(국토연구 원, 2004). 이를 계기로 1990년대에 정부, 시민단체, 학계
Corresponding author : Lee, Seungjoo Tel: 031-420-3273
E-mail: [email protected]
를 중심으로 우리나라 산줄기 및 지리 인식체계의 정당 성을 확립하는 차원에서 백두대간의 개념 형성과정에 대 한 연구가 시작되었고(산림청, 1996; 산림청, 대한지리학 회, 1997; 양보경, 1997; 국토연구원, 녹색연합, 1999) 이 후 연구는 생태계 현황 파악을 통한 자료축적과 이를 활 용한 관리 및 복원 측면으로 진행되었다(조현서, 이수동, 2010). 백두대간의 개념과 관리의 중요성이 알려진 1990 년대 후반부터 생태계 연구는 활성화되었는데 재와 재구 간의 식물종리스트 파악(박상곤 등, 2009; 임동옥 등, 2004, 오구균, 이정은, 2003), 구간별 현존식생과 식물군 집구조를 파악하는 연구들이 수행되었다(오구균, 박석곤, 2002; 최송현, 오구균, 2003; 김갑태, 엄태원, 2004; 추갑 철, 김갑태, 2004; 2005). 2000년대에 들어서는 식물생태 계, 동물생태계, 토양생태계, 이용환경 등에 대한 개별적
인 연구가 진행되기도 하였으나 1990년대부터 쌓아온 분야별 연구결과를 바탕으로 복원 또는 관리 개념이 제 시되기에 이르렀다. 결국 이후 연구는 훼손경향분석(이 동근 등, 2007), 훼손지 복원을 위한 모델 및 수종 선정 (김남춘 등, 2008; 이지혜 등, 2008), 관리범위 및 방향설 정(신준환, 2004; 유기준, 2002; 권태호 등, 2002; 2004;
산림청, 한국환경생태학회, 2002; 2003; 임업연구원, 2003), 인위적인 훼손실태 및 효율적인 관리방안 및 복 원시스템 제안(권태호 등, 2004; 유기준 등, 2004; 산림 청, 한국환경생태학회, 2001; 국토연구원, 2001; 2002a;
2002b) 등 현황조사 및 분석, 관리 및 복원 방안 연구 등 생물다양성 증진과 지속가능한 발전을 목표로 하는 유연하고 다양한 결과들이 도출되었다.
이러한 연구로 인하여 백두대간이라는 용어는 익숙해 졌으나 지맥인 정맥에 대한 현황연구는 현재 미비한 상 태이며, 특히 낙동정맥에 대한 연구는 가지산도립공원과 특정구간에 대해 수행된 몇몇 연구결과(김인택, 1998; 김 인택 등, 1996; 1997; 송유성, 1997; 백재봉 등, 2000; 김 갑태 등, 2004; 신경옥, 2004; 조현서, 이수동, 2010)가 보 고된 이외에는 최근에 수행된 연구결과가 거의 없는 실 정이다. 따라서 본 연구는 낙동정맥 구간 중 애미랑재를 중심으로 현황조사를 실시하여 식생구조를 밝히고 그 결 과를 토대로 향후 정맥관리 및 복원방안 제안을 위한 기 초자료를 제공하고자 한다.
II. 연구내용 및 방법
1. 연구대상지개황 및 조사구 설정
백두대간은 백두산에서 시작되어 동쪽 해안선을 끼고 남쪽으로 흐르다가 내리뻗어 지리산에 이르고 있으며 태 백산에서 서남쪽 소백산으로 이어지는 맥은 태백산 북쪽 에서 벗어나 경북 울진의 백병산과 평해의 백암산, 영덕 의 용두산, 청송의 주왕산을 지나고, 줄기차게 남쪽으로 만 달려 경주의 단석산, 청도의 운문산, 언양의 가지산, 양산의 취서산, 동래의 금정산을 지나 몰운대에 이어지 는데, 이것이 낙동정맥이다(송유성, 2007). 즉, 태백산 지 역의 매봉산에서 남쪽으로 뻗어 경상남북도를 거쳐 부산 다대포 물운대(沒芸臺)에 이르는 약 359㎞에 이르는 백 두대간의 정맥 중 하나를 일컫는다(김동필 등, 2004).
본 연구대상지는 백두대간의 정맥중 하나인 낙동정맥 능선부 구간중 영양군과 울진군의 경계부 백암산의 능선 부와 울진군 남서쪽 구간 사이에 입지한 애미랑재 구간 을 연구대상지으로 설정하였다. 식물군집구조는 능선부
및 계곡부의 대표적인 식생 및 식생군락, 입지환경의 변 화가 있는 지역에 조사구를 설정하여, 군집의 유형에 따 라 총 25개소를 설정하였다. 각 조사구는 10m×10m(100
㎡) 방형구를 기본으로 하여 1∼2개소를 함께 설치하였 으며 각 조사구의 주요 환경인자와 식생구조를 조사하였 다.
Figure 1 The survey site and plots of vegetation structure in Aemirang hill, Nakdongjeongmaek.
2. 식생조사 및 군집구조분석
식생구조는 방형구법(Quadrat Method)으로 조사하였으 며 각 조사구내에 출현하는 수종을 대상으로 수관의 위 치에 따라 교목, 아교목, 관목으로 구분하여 수종, 개체 수 및 흉고직경을 측정하여 기록하였다. 교목 및 아교목 층은 흉고직경(DBH) 2cm 이상인 수목을 대상으로 수종, 수고, 지하고, 수관폭을 조사하였고 관목층은 수고, 지하 고, 수관폭을 조사하였다.
조사자료를 토대로 각 조사구의 수종별 상대적 우세 를 비교하기 위하여 Curtis and McIntosh(1951)의 중요치 (Importance Percentage: I.P.)를 통합하여 백분율로 나타낸 상대우점치를 수관층위별로 분석하였다(Brower and Zar, 1977). 상대우점치(Importance Percentage: I.P.)는 각 종별 (상대밀도+상대피도)/2 로 계산하였으며 개체들의 크기를 고려하여 수관층위별로 가중치를 부여한((교목층 I.P.×3 + 아교목층 I.P.×2 + 관목층 I.P.×1)/6) 평균상대우점치 (M.I.P.)를 구하였다(박인협, 1986). 식생조사자료를 바탕 으로 TWINSPAN classification 분석을 실시하였다(Hill, 1979). 종 구성의 다양한 정도를 나타내는 척도인 종다 양도는 Shannon의 수식(Pielou, 1975)을 적용하여 종다양
Community Pd Pd-Qm Qv Qm Qm-Bd Qm-Qv Ll
Altitude(m) 925 720 660 935 800 720 900 940 920 700 780 910 750 680
Aspect W10S W10S E25S S25W S20E E20S S30W S35W E10S N10W E30N S30E S15W E05S
Slope(°) 26 31 23 25 30 26 23 23 23 21 24 25 26 25
Canopy
Height(m) 17 12 18 16 17 18 17 13 15 18 13 17 15 22
Mean DBH(cm) 35 15 20 23 30 25 27 13 12 35 11 35 12 16
Coverage(%) 60 70 70 80 40 70 80 80 80 60 80 60 80 70
Understory
Height(m) 7 3 6 8 6 6 4 4 8 10 4 7 6 7
Mean DBH(cm) 5 5 5 6 6 5 4 4 6 6 4 5 5 4
Coverage(%) 10 40 10 40 30 10 10 20 15 15 20 10 15 10
Shrub Height(m) <2.0 <2.0 <2.5 <2.0 <2.0 <2.0 <2.0 <2.0 <1.5 <2.0 <2.0 <2.0 <2.0
Coverage(%) 30 25 30 15 15 60 50 40 20 5 30 10 40
Area(㎡) 500 500 400 300 200 500 100
* Pd: Pinus densiflora, Qv: Quercus variabilis, Qm: Quercus mongolica, Qs: Quercus Serrata, Ud: Ulmus davidiana var. japonica, Bs:
Betula scgmidtii, Ll: Larix leptolepis
Table 1 General description of the surveyed site
Ⅰ
Ⅱ Ⅳ
Ⅲ Ⅴ
18, 19, 20, 21, 22
9, 10, 12,
13, 17 7, 8, 24, 25 5, 6, 11, 23 1, 2, 3, 4,
14 15, 16
Figure 2 TWINSPAN classificatio ordination of twenty-five plots in Yongso Valley.
도(Species diversity, H'), 균재도(Evenness, J'), 우점도 (Dominance, D')를 종합적으로 비교하였으며 Whittaker (1956)의 수식을 이용하여 유사도지수(Similarity index)를 분석하였다.
III. 결과 및 고찰
1. 군집별 개황
대상지 25개 조사구별 군집유형은 TWINSPAN 분석결 과에 의해 Figure 2와 같이 6개의 유형으로 구분되었다.
제ⅠDivision은 쇠물푸레, 소나무, 철쭉나무 등이 출현하 는 군과 조록싸리가 출현하는 군으로 구분되었다. 쇠물 푸레, 소나무, 철쭉나무가 출현하는 군은 제ⅡDivision에 서 소나무, 굴참나무가 출현하는 군과 쇠물푸레, 물박달 나무가 출현하는 군으로 나누어졌고 소나무, 굴참나무가 출현하는 군은 제ⅢDivision에서 노린재나무와 굴참나무 가 출현하는 소나무군집과 그렇지 않은 소나무-신갈나무 군집으로 최종 분류되었다. 제ⅡDivision에서 쇠물푸레와 물박달나무가 출현하는 군은 신갈나무-물푸레나무군집으 로 명명되었다. 제ⅠDivision에서 조록싸리가 출현하는 군은 제ⅣDivision에서 물박달나무가 출현하는 신갈나무-
물박달나무군집과 출현하지 않는 군으로 구분되었다. 출 현하지 않는 군은 제ⅤDivision에서 굴참나무가 출현하 는 신갈나무-굴참나무군집과 출현하지 않는 신갈나무군 집, 일본잎갈나무군집으로 최종 구분되었다. 이상의 TWINSPAN에 의한 군집분류 결과 소나무군집, 소나무- 신갈나무군집, 굴참나무군집, 신갈나무군집, 신갈나무-물 박달나무무군집, 신갈나무-굴참나무군집의 6개로 구분되 었고, 일본잎갈나무가 교목층에서 우점하는 조사구는 따 로 분류하여 최종 7개 유형으로 구분되었다.
낙동정맥 애미랑재 지역 각 조사구의 주요인자 및 출 현종수를 Table 1에 나타내었다. 조사구는 영양군과 울 진군의 경계부 백암산 능선부와 울진군 남서쪽 구간 사 이에 위치한 애미랑재에 걸쳐있는 천연림으로 현존식생 을 감안하여 설정되었으며, 조사구들은 해발고 660~
940m, 경사도 20~31°로 다소 급경사지에 위치하고 있 었다. 교목층 수관 울폐도는 50~80%의 범위이며, 교목 층과 아교목층 조사구당 목본식물 출현종 수는 7~15종 으로 나타났다. 식물군집구조 군집을 구분한 결과 소나 무군집, 소나무-신갈나무군집, 굴참나무군집, 신갈나무군 집, 신갈나무-물박달나무무군집, 신갈나무-굴참나무, 일본 잎갈나무군집 등의 7개 유형으로 구분되었다.
Community P. densiflora P. densiflora-Q. mongolica Q. variabilis
scientific name c u s m c u s m c u s m
Pinus densiflora 88.95 4.33 - 45.92 54.15 16.23 - 32.49 4.14 - - 2.07
Quercus variabilis 1.73 20.09 - 7.56 - - - - 93.24 29.38 1 56.58
Quercus mongolica 6.26 32.22 23.38 17.77 45.85 79.3 8.54 50.78 - 54.38 6.58 19.22
Populus maximowiczii - - - - - - - - 2.62 - - 1.31
Acer pseudo-sieboldianum - - - - - - 8.86 1.48 - - - -
Lespedeza cyrtobotrya - 2.19 3.74 1.35 - - - - - - - -
Rhododendron mucronulatum - - 10.29 1.72 - - 9.68 1.61 - - - -
Rhododendron schlippenbachii - 25.71 24.58 12.67 - 4.47 53.73 10.45 - 1.99 - 0.66
Gleditsia japonica var. koraiensis - - - - - - - - - - 37.95 6.33
Tripterygium regelii - - - - - - - - - - 10.58 1.76
Fraxinus rhynchophylla 3.07 - 0.77 1.66 - - 2.87 0.48 - 1.65 12.01 2.55
Fraxinus sieboldiana - 13.27 30.99 9.59 - - 6.82 1.14 - 4.41 10.93 3.29
Symplocos chinensis for. pilosa - - - - - - - - - 4.87 8.87 3.1
etc. MIP〈 1.0 4species 1.78 3species 1.59 5species 3.12
Table 2 Importance percentage of the woody plants by the stratum in seven communities
community Q. mongolica Q. mongolica-B. davurica Q. mongolica-Q. variabilis Larix leptolepis
scientific name c u s m c u s m c u s m c u s m
Quercus mongolica 94.28 60.52 6.43 68.39 69.66 33.56 14.23 48.39 44.35 73.48 21.78 50.3 - 26.9 4.58 9.73
Quercus variabilis 3.31 1.42 - 2.13 3.27 - - 1.64 55.66 19.31 - 34.27 - 19.07 - 6.36
Betula davurica - 1.26 - 0.42 27.07 7.08 - 15.9 - - - - - - - -
Larix leptolepis 1.12 4.51 - 2.06 - - - - - - - - 100 44.18 - 64.73
Corylus sieboldiana - 0.91 - 0.3 - 16.34 5.01 6.28 - - - - - - - -
Tripterygium regelii - - - - - - 19.88 3.31 - - - - - - - -
Stephanandra incisa - - - - - - - - - - 18.62 3.1 - - - -
Lindera obtusiloba - - - - - - - - - - 18.31 3.05
Lespedeza maximowiczii - - 28.28 4.71 - - - - - - 8.81 1.47 - - 65.86 10.98
Lespedeza cyrtobotrya - - 1.98 0.33 - - - - - - 8.1 1.35 - - - -
Rhododendron mucronulatum - 12.46 - 4.15 - - 9.31 1.55 - - - -
Maackia amurensis - - 1.1 0.18 - - - - - 7.22 - 2.41 - - - -
R. yedoense var. poukha-nense - - - - - - 33.17 5.53 - - - - - - - -
Fraxinus sieboldiana - 12.14 2.92 4.53 - 14.97 16.61 7.76 - - - - - 9.84 9.16 4.81
Rhododendron schlippenbachii - 3.42 27.91 5.79 - 5.81 5.55 2.86 - - - - - - - -
Tripterygium regelii - - - - - - 6.68 1.11 - - - -
Symplocos chinensis for. pilosa - - - - - 5.38 5.55 2.72 - - - - - - - -
Fraxinus rhynchophylla - 2.08 2.93 1.18 - 4.41 - 1.47 - - - - - - - -
Fraxinus sieboldiana - - - - - - - - - - 21.7 3.62 - - - -
Tripterygium regelii - - 16.71 2.79 - - - - - - - - - - - -
Acer mono - 4.71 - 1.57 - - - - - - - - - - - -
Symplocos chinensis for. pilosa - 3.49 4.15 1.86 - - - - - - - - - - - -
etc. MIP〈 1.0 8species 3.76 - 2species 0.83 1species 0.35
2. 상대우점치
Table 2는 군집별 주요 출현종의 상대우점치 및 평균 상대우점치를 나타낸 것이다. 능선부와 급경사지에 분포 하는 소나무군집의 교목층에서는 소나무가 상대우점치 (I.P.) 88.95%로 우점종이었고 신갈나무(I.P.: 6.26%), 물푸 레나무(I.P.: 3.07%)가 출현하였다. 아교목층은 교목성상 인 신갈나무(I.P.: 32.22%), 굴참나무(I.P.: 20.09%) 등과 관목성상인 철쭉꽃(I.P.: 25.71%)이 경쟁상태이었고 쇠물 푸레나무(13.27%), 소나무(4.33%) 등이 동반 출현하였다.
관목층은 쇠물푸레나무(I.P.: 30.99%), 철쭉꽃(I.P.:
24.58%), 신갈나무(I.P.: 23.38%)의 세력이 우세하였다. 본
군집은 교목층 소나무 세력이 우세하여 당분간은 소나무 군집을 유지하겠으나, 아교목층에 신갈나무, 굴참나무 등 교목성상의 낙엽성참나무류의 세력이 확대되고 있어 장 차 이들 수종과의 경쟁이 예상되었다. 소나무-신갈나무 군집의 교목층은 소나무(I.P.: 54.15%)와 신갈나무(I.P.:
45.85%)가 경쟁하고 있는 상태이었고, 아교목층에서는 소나무(I.P.: 16.23%) 세력에 비해 신갈나무(I.P.: 79.30%) 세력이 우세하였다. 관목층은 철쭉꽃(I.P.: 53.73%)이 우 점종이었고 진달래(I.P.: 9.68%), 당단풍(I.P.: 8.86%), 신갈 나무(I.P.: 8.54%), 쇠물푸레나무(I.P.: 6.82%) 등이 주요 동반종이었다. 본 군집은 교목층에서 소나무와 신갈나무
1
1 1
1 1
1
11 1 2
2 2
2 2
2 22 3
3 3
3 3
3 4
4 4
4 4
4 4
4 4
4 10m
10m
0 10m
0 10m 1
1
1 1
1 1 1
1 1 11
1
1 2
2 2
2 2 2
2
2 2
2
2 2 22 2
2 2 22 0m
10m
0 10m
0 10m
1. P. densiflora 2. Q. mongolica 3. F. sieboldiana 4.
Rhododendron schlippenbachii 1. P. densiflora 2. Q. mongolica
Pinus densiflora community's Pinus densiflora-Quercus mongolica community's
1 1
1
1 11
1 1
1 1
1 1
1 1
1
11 1 1 11
11 11 11 11
10m 0
10m
10m
0 10m
1. Q. variabilis 2. P. densiflora 3.Q. mongolica 4. R.
schlippenbachii 1. Q. mongolica
Quercus variabilis community's Quercus mongolica community's
1
1
1 1
1
1 1
1
2 2 2
2 2
2
12 11
22
2 11
11 2 1
2 3
4 10m
10m
0 10m
0 10m
1 1 1
1
21 1
1 1 2
2
2 2
2 2 2 2 3
3 4
664 5
5 5
5 5 5
6 6
5 55 5 10m
10m
0 10m
10m 0
1. Q. mongolica 2. Q. mongolica 3. Maackia amurensis 1. Q. mongolica 2. B. davurica 3. Q. mongolica 4. F.
rhynchophylla 5. C. sieboldiana 6. S. chinensis for. pilosa
Quercus mongolica-Q. variabilis community's Quercus mongolica-Betula davurica community's
Figure 3 The diagram of major community's vegetation structure in Aemirang hill.가 경쟁하고 있으나 아교목층에서는 신갈나무의 세력이 우세한 것으로 분석되었다. 한편, 우리나라의 식생천이는 소나무에서 참나무류로 진행되는 것이 일반적이고(박인 협 등, 1988) 특히, 대부분 신갈나무, 졸참나무 등으로의
천이를 예측하고 있어(이경재 등, 1991; 1994; 1997; 박인 협 등, 1989) 신갈나무로 천이가 진행될 것으로 판단되 었다.
굴참무군집의 교목층에서는 굴참나무(I.P.: 93.24%)가
우점종이었고 소나무(I.P.: 4.14%), 황철나무(I.P.: 2.62%)가 출현하였다. 아교목층은 신갈나무(I.P.: 54.38%)와 굴참나 무(I.P.: 29.38%) 등의 교목성상인 참나무류가, 관목층은 조록싸리(I.P.: 37.95%)가 우점하였으며 쇠물푸레나무, 미 역줄나무, 신갈나무, 주엽나무, 다릅나무 등 낙엽활엽수가 주요 출현종이었다. 본 군집은 굴참나무의 세력이 우세 한 반면, 경쟁종인 낙엽활엽수의 출현이 없어 당분간 현 상태를 유지할 것으로 판단되었다. 신갈나무군집은 교목 층과 아교목층에서 신갈나무가 상대우점치(I.P.) 94.28%, 60.52%로 우점종이었고 교목층에서는 굴참나무(I.P.:
3.31%)가, 아교목층에서는 쇠물푸레나무(I.P.: 12.14%)가 주요 출현종이었다. 관목층은 조록싸리(I.P.: 28.28%), 철 쭉꽃(I.P.: 27.91%)이 우점하였으며 미역줄나무(I.P.:
16.71%), 신갈나무(I.P.: 6.43%) 등 다양한 수종이 출현하 였다. 본 군집은 교목층 신갈나무의 우점도가 높고 아교 목층에서도 비교적 세력이 강하여 지속적으로 현 상태를 유지할 것으로 판단되었다. 신갈나무-물박달나무군집의 교목층과 아교목층은 신갈나무가 상대우점치(I.P.) 69.66%, 33.56%로 우점종이었고 교목층에서는 물박달나 무(I.P.: 27.07%)가, 아교목층에서는 참개암나무(I.P.:
16.34%), 쇠물푸레나무(I.P.: 14.97%), 진달래(I.P.: 12.46%) 가 동반 출현하였다. 관목층은 산철쭉(I.P.: 33.17%), 미역 줄나무(I.P.: 19.88%), 쇠물푸레나무(I.P.: 16.61%), 신갈나 무(I.P.: 14.23%) 등 다양한 종이 분포하였다. 본 군집은 교목층에서 신갈나무와 물박달나무가 경쟁하고 있었으나 하층에 차대를 형성할 수 있는 우점종이 출현하지 않아 당분간 현상태를 유지할 것으로 판단된다. 신갈나무-굴참 나무군집의 교목층은 굴참나무(I.P.: 55.66%)와 신갈나무 (I.P.: 44.35%)가 경쟁하고 있는 상태이었고 아교목층에서 는 신갈나무(I.P.: 73.48%)가 경쟁종인 굴참나무(I.P.:
19.31%), 다릅나무(I.P.: 7.22%)에 비해 세력이 월등히 우 세하였다. 관목층에서는 신갈나무(I.P.: 21.78%), 쇠물푸 레나무(I.P.: 21.70%), 국수나무(I.P.: 18.62%)가 주요 출현 종이었다. 따라서 본 군집은 낙엽성참나무인 신갈나무 와 굴참나무가 경쟁하고 있으나 하층으로 갈수록 신갈 나무의 세력이 우세해져 장차 신갈나무로의 천이가 예 측되었다.
일본잎갈나무군집의 교목층에서는 일본잎갈나무(I.P.:
100.00%) 1종만이 출현하였고 아교목층은 일본잎갈나무 (I.P.: 44.18%)와 낙엽성 참나무류인 신갈나무(I.P.:
26.90%)와 굴참나무(I.P.: 19.07%) 등이 경쟁하고 있었다.
관목층은 조록싸리(I.P.: 65.86%)의 우점도가 높았고 생강 나무(I.P.: 18.31%), 쇠물푸레나무(I.P.: 9.16%), 신갈나무 (I.P.: 4.58%) 등이 함께 출현하였다. 본 군집은 교목층과 아교목층에 식재된 일본잎갈나무의 세력이 당분간은 유
지되겠으나 하층에 신갈나무, 굴참나무 등 낙엽성 참나 무류의 세력이 확장되고 있어 향후 이들 군집으로의 천 이가 예측되었다.
이상을 종합해보면 소나무군집, 소나무-신갈나무군집 은 교목층에서 소나무가 상대우점치(I.P.) 88.95%, 54.15%
로 우점하고 있으나 평균상대우점치(M.I.P)는 45.92%, 32.48%로 낮아지고 있는 바, 이는 하층에 출현하는 신갈 나무, 굴참나무 등과의 경쟁에 의한 것으로 분석되었다.
기존 식물군집구조 연구에서 우리나라의 천이는 소나무
→신갈나무, 졸참나무 등 참나무류→서어나무, 까치박달 등으로의 진행을 예측하고 있어(이경재 등, 1992; 최송 현, 이경재, 1993) 본 대상지에서도 소나무가 우점하는 군집은 장차 하층의 우점종인 신갈나무로의 천이가 예상 되었다. 굴참나무와 신갈나무가 우점하는 군집은 교목층 우점종인 낙엽성 참나무류의 세력이 우세하고 서어나무, 까치박달 등 차대를 형성할 수 있는 수종이 하층에 출현 하지 않아 현상태를 유지할 것으로 판단되었다. 일본잎 갈나무군집은 조림된 것으로 당분간은 현상태를 유지하 겠으나 인공림인 아까시나무가 참나무류로의 천이를 예 측한 연구결과(최송현, 이경재, 1993)가 있는바 장기적으 로는 아교목층 우점종인 신갈나무로의 천이 진행을 기대 할 수 있다.
3. 흉고직경급별
7개 군집의 흉고직경급별(DBH) 분포현황을 살펴보면 (Table 9), 소나무군집의 경우 DBH 7~32cm 구간에 소 나무 38주가 집중 분포하였고 DBH >=52인 소나무 3주 가 출현하였다. 경쟁종에 해당되는 교목성상 신갈나무와 굴참나무는 중소경목으로 DBH 2~22cm 구간에 1~6주 씩 관찰되었으며, 관목층에는 72주가 출현하였다. 하층에 쇠물푸레와 철쭉꽃 등의 개체수가 많았으나 아교목․관 목성상으로 천이에 영향은 없을 것이다. 따라서 본 군집 은 소나무가 우점하는 현상태를 유지겠으나 하층에 경쟁 종인 교목성상의 신갈나무, 굴참나무가 출현하고 있어 장기적으로는 낙엽성 참나무류로의 천이가 예측되었다.
소나무-신갈나무군집은 소나무가 DBH 7~37cm구간에 14주, 신갈나무가 DBH 2~27㎝ 구간에 28주 분포하여 경쟁상태인 것으로 판단되었으며 하층으로 갈수록 신갈 나무의 세력은 커지고 있었다. 교목층에서 소나무가 신 갈나무와 경쟁하고 있으나 소나무→낙엽성참나무류로의 천이가 일반적인 우리나라 산림에서 하층으로 갈수록 우 세해지는 신갈나무로 세력으로 보아 신갈나무로의 천이 가 진행되고 있는 것으로 판단되었다.
굴참나무군집은 굴참나무 DBH 7~74cm 구간에 30주
Commuties Species name SH D2 D3 D4 D5 D6 D7 D8 D9 D10 D11 D12
P. densiflora
Pinus densiflora - - 9 19 6 2 2 - - 1 - 3
Quercus variabilis - 1 4 - 1 - - - - - - -
Quercus mongolica 72 6 1 1 3 - - - - - - -
Rhododendron schlippenbachii 56 6 2 - - - - - - - - -
Fraxinus sieboldiana 136 3 1 - - - - - - - - -
P. densiflora Q. mongolica -
Pinus densiflora - - 1 5 - 4 3 1 - - - -
Quercus mongolica 16 4 8 12 3 1 - - - - - -
Rhododendron schlippenbachii 56 1 - - - - - - - - - -
Q. variabilis
Quercus variabilis 4 - 2 2 3 4 9 5 2 3 - -
Quercus mongolica 16 9 8 4 - - - - - - - -
Lespedeza maximowiczii 116 - - - - - - - - - - -
Q. mongolica
Quercus variabilis - - 1 - 2 1 - - - - - -
Quercus mongolica 36 5 36 20 10 2 1 - - - 1 -
Lespedeza maximowiczii 144 - - - - - - - - - - -
Q. mongolica - B davurica
Betula davurica - - - 1 - 2 - 2 - 1 - -
Corylus sieboldiana 16 4 1 - - - - - - - - -
Quercus mongolica 36 - 2 12 8 1 1 - 1 - - -
Tripterygium regelii 80 - - - - - - - - - - -
Fraxinus sieboldiana 48 3 1 - - - - - - - - -
Q. mongolica - Q. variabilis
Quercus variabilis - - 6 6 3 3 2 2 - - - 1
Quercus mongolica 56 - 5 9 5 5 - 1 - - - -
Fraxinus sieboldiana 72 - - - - - - - - - - -
Larix leptolepis
Larix leptolepis - - 2 5 4 - - - - - - -
Quercus mongolica 4 2 - - - - - - - - - -
Lespedeza maximowiczii 68 - - - - - - - - - - -
* SHR: SHRUB, D2: 2<=ba<7, D3: 7<=ba<12, D4: 12<=ba<17, D5: 17<=ba<22, D6: 22<=ba<27, D7: 27<=ba<32, D8: 32<=ba<37, D9: 37<=ba<42, D10: 42<=ba<47, D11:47<=ba<52, D12:ba>=52
Table 3 The distribution of major woody species' DBH by 7 communities in Aemirang hill
가 고르게 출현한 반면에 신갈나무는 DBH 2~17cm 구 간에 11주만이 출현하였고, 관목층에서는 72주가 분포하 였다. 본 군집은 교목층 굴참나무의 세력이 우세하여 현 상태를 유지할 것이다. 신갈나무군집은 DBH 7~22cm인 구간에 신갈나무가 66주 분포하였고 DBH 47~52㎝ 구 간에 1주가 출현하였다. 교목성상인 굴참나무가 DBH 7~47㎝ 구간에 4주 출현하였으나 개체수가 적었고 아 교목․관목층에 경쟁종이 출현하지 않아 신갈나무가 우 점하는 현상태를 유지할 것으로 판단되었다. 신갈나무- 물박달나무군집에서는 신갈나무가 DBH 7~42㎝ 사이에 25주, 물박달나무가 DBH 12~47㎝ 구간에 1~2주씩 분 포하였다. 본 군집은 물박달나무에 비해 신갈나무의 세 력이 우세하였으며, 특히 관목층에 신갈나무 차대목이 형성되어 당분간은 물박달나무군집을 유지하겠으나 장차 신갈나무가 우점하는 군집으로 변할 것으로 예측할 수 있다. 신갈나무-굴참나무군집에서는 DBH 7~37㎝ 구간 에 굴참나무 22주, 신갈나무 25주가 고르게 분포하고 있 었으나 관목층에서는 신갈나무 56주만이 출현하였다. 따 라서 본 군집은 관목층에 신갈나무 차대목의 출현이 활 발하고 굴참나무 차대목의 출현이 없는데다가, 전 흉고 직경급에서 굴참나무와 신갈나무의 세력이 고르게 분포
하고 있어 세력 균형이 유지될 것으로 판단되었다.
일본잎갈나무군집에서는 DBH 7~22㎝ 사이에 일본잎 갈나무 11주가 출현하였고, 교목성상의 수목은 DBH 7~
12㎝의 굴참나무 1주, DBH 2~7㎝에 신갈나무 2주가 출현하였다. 관목층에서는 신갈나무, 조록싸리 등이 분포 하였으나 세력이 약해 당분간 일본잎갈나무군집을 유지 할 것으로 예측되었다.
4. 종수 및 개체수
낙동정맥 애미랑재 지역 식물군집의 유형별 종수 및 개체수를 분석한 결과이다(Table 10). 군집별 비교를 위 하여 단위면적 100㎡에 출현하는 평균 종수 및 개체수 로 환산하여 산출하였다. 산출결과 전체출현종수는 7~
15종이었으며, 각 군집별 평균 출현종수는 신갈나무-물 박달나무군집이 5.3종으로 최소였으며 소나무군집이 9.7 종으로 최대였다. 출현 종수는 굴참나무군집, 신갈나무군 집, 소나무-신갈나무군집에서 다양한 종이 출현한 반면, 신갈나무-물박달나무군집, 신갈나무-굴참나무군집 등 급 경사지역에 입지한 군집에서는 종수가 낮았다.
단위면적당(100㎡) 군집별 출현 개체수는 교목층 7.
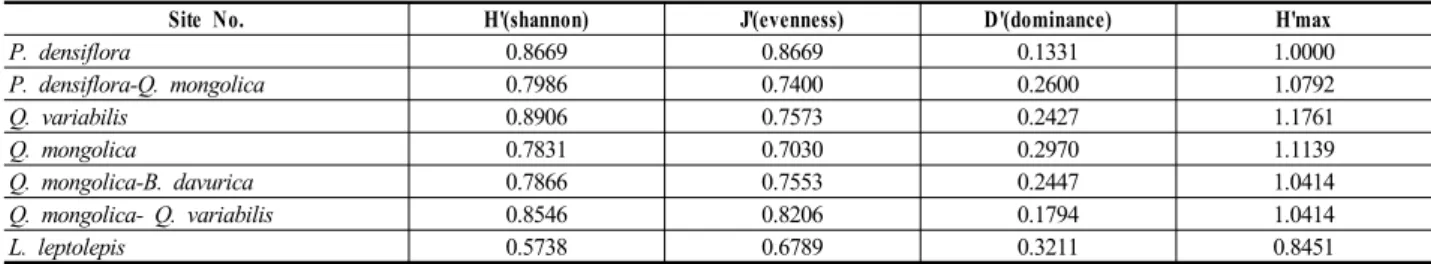
Site No. H'(shannon) J'(evenness) D'(dominance) H'max
P. densiflora 0.8669 0.8669 0.1331 1.0000
P. densiflora-Q. mongolica 0.7986 0.7400 0.2600 1.0792
Q. variabilis 0.8906 0.7573 0.2427 1.1761
Q. mongolica 0.7831 0.7030 0.2970 1.1139
Q. mongolica-B. davurica 0.7866 0.7553 0.2447 1.0414
Q. mongolica- Q. variabilis 0.8546 0.8206 0.1794 1.0414
L. leptolepis 0.5738 0.6789 0.3211 0.8451
Table 5 7 communities' species diversity indices in Aemirang hill
Communities
Q. variabilis Q. mongolica P. densiflora P. densiflora
-Q. mongolica Q. mongolica
- Q. variabilis Q. mongolica -B. davurica
Q. mongolica 35.04
P. densiflora 24.40 60.84
P. densiflora
-Q. mongolica 33.65 34.38 64.86
Q. mongolica
-Q. variabilis 60.18 61.36 53.89 31.99
Q. mongolica
-B. davurica 30.76 59.41 55.08 33.22 56.31
L. leptolepis 26.43 24.20 11.22 21.32 21.18 16.18
Table 6 Similarity index among 7 communities in Aemirang hill
Community Individuals Mean number of
species
Number of total species
Canopy Under story Shrub Total
P. densiflora 7.0 3.8 38.0 48.8 9.7 10
P. densiflora-Q. mongolica 25.0 13.0 158.0 196.0 8.0 12
Q. variabilis 8.7 12.0 93.3 114.0 8.3 15
Q. mongolica 14.5 6.5 77.0 98.0 6.3 13
Q. mongolica-B. davurica 9.0 7.3 69.3 85.7 5.3 11
Q. mongolica- Q. variabilis 13.7 2.7 81.3 97.7 5.7 11
L. leptolepis 9.0 6.0 100.0 115.0 7.0 7
Table 4 7 Communities of species and individuals in Aemirang hill (Unit: 100㎡)
0~25.0개체, 아교목층 3.8~13.0개체, 관목층 38.0~100.0 개체이었다. 소나무-신갈나무군집이 교목층 25.0개체, 아 교목층 13.0개체 관목층 158.0개체로 가장 많은 개체가 출현한 반면 소나무군집은 다양한 종이 출현한 것과 다 르게 총 출현개체수가 48.8개체로 가장 적었다. 이는 소 나무군집이 능선부의 급경사지에 분포하는 원인 때문인 것으로 분석되었다.
5. 종다양도 및 유사도지수
낙동정맥 애미랑재 지역 7개 유형 식물군집별 종다양 도지수를 분석한 결과 Shannon의 종다양도 값의 분포는 0.5738~0.8906로 나타났다. 이 중 남사면과 북사면의 주 연부에 해당되는 건조한 능선부 또는 상부의 남사면에 위치한 굴참나무군집(0.8906), 소나무군집(0.8669), 신갈나 무-굴참나무군집(0.8546)의 종다양도 지수가 높았고 저지
대에 위치하나 식재한지 얼마되지 않은 일본잎갈나무군 집(0.5738)의 종다양도가 가장 낮았다. 신갈나무가 우점 하는 군집의 종다양도 지수는 중간정도이며 대부분 계곡 과 능선의 중간정도에 위치하여 상대적으로 출현종이 적 었던 것이 그 원인으로 판단되었다. 종다양도 값은 가지 산에서 능동산 구간의 0.6651~1.0425(김동필, 최송현, 2004), 낙동정맥 용소골 계곡의 0.7914~0.9442(조현서, 이수동, 2010)의 지수값과 비교했을 때 군집모두 0.9 미 만으로 전반적으로 낮은 상태라고 할 수 있다.
군집간 유사도지수를 분석한 결과(Table 12) 11.22~
64.86%로 나타났다. 소나무군집과 소나무-신갈나무군집, 신갈나무군집과 신갈나무-굴참나무군집, 신갈나무-소나무 군집의 유사도가 60% 이상으로 타 군집간에 비해 유사 도가 높았다. 반면에 일본잎갈나무는 전체 군집과 유사 도 25% 이하로 이질적이었고 굴참나무군집과 소나무-신 갈나무군집, 소나무-신갈나무군집과 신갈나무가 우점하
는 군집간 유사도가 35% 미만으로 이질적인 것으로 분 석되었다. 군집별 유사도지수가 다양하게 나타난 이유는 능선부를 중심으로 다양한 환경요건을 갖는 사면 지역에 조사구를 설치했기 때문인 것으로 판단되었다.
6. 정맥 관리 및 복원 방향 제안
기후변화에 의한 산림생태계의 천이영향은 산림군집 구조의 다양성을 저해하는 간접적인 요소로 작용할 수 있으며, 이는 생물종 다양성 및 종의 감소의 연결고리로 작용하고 있다. 따라서 이에 대한 생물종 다양성 보전과 유지가 필요한 상태이다. 무엇보다도 산림생태계 생물종 다양성 유지 및 보전을 위한 가장 효과적인 방법은 백두 대간과 정맥을 중심으로 건강한 상태의 산림군집구조를 가지고 있는 곳에 대한 관리와 훼손된 지역에 대해서는 복원이 필요하다. 본 연구결과에서 나타난 바와 같이 낙 동정맥 애미랑재 산림생태계 종다양도 값은 다른 지역 (가지산-능동산 구간, 낙동정맥 용소골)에 비해 전반적으 로 낮은 상태를 보이고 있는 실정이었다. 따라서 애미랑 재 종다양성 증가를 위해 관리 및 복원 방향을 제안해 보자면 다음과 같이 정리할 수 있다.
애미랑재 구간 식생분포 특성 및 식생유형별 천이 경향 분석을 통해 애미랑재 대표 식생 유형은 소나무군 집, 소나무-신갈나무군집, 굴참나무군집, 신갈나무군집, 신갈나무-물박달나무군집, 신갈나무-굴참나무군집, 일본 잎갈나무군집의 7개 유형으로 구분할 수 있었으며, 소나 무 군집과 소나무-신갈나무군집의 경우, 소나무가 우점 하여 소나무 세력이 우세한 군집에 대해서 우리나라에서 소나무림이 가지고 있는 정서적, 상징적, 경관적 가치를 고려하여 생물종 다양성 관리와 문화적 가치측면에서 장 기적으로 소나무군집의 유지를 목표로 하는 관리 및 복 원 방안을 수립하도록 한다. 소나무와 신갈나무등과 같 은 참나무류가 경쟁하는 군집에 경우(소나무-신갈나무군 집)와 소나무의 우점도가 높거나 소나무 세력이 우세한 군집에 대해서는 소나무와 생태적 지위가 중복되는 아교 목성상 및 관목 성상의 수목을 관리하여 방해극상적 방 안을 도입하고 식재 및 하예작업을 통해 소나무 하층식 생 발달을 유도하는 적극적 관리 및 복원 방안을 수립한 다. 소나무와 참나무류 간의 경쟁군집 중 참나무세력이 우세한 군집(소나무-신갈나무군집)과 참나무류 군집(굴참 나무군집, 신갈나무군집, 신갈나무-물박달나무군집, 신갈 나무-굴참나무군집), 일본잎갈나무 군집(인공림)에 대해 서는 산림 천이과정 및 생태계 순리에 맞게 자연적으로 천이가 진행될 수 있게 하며, 인위적인 간섭 및 교란이 발생되지 않도록 조방적 관리 및 복원 방안을 수립한다.
IV. 결 론
본 연구는 애미랑재 구간의 식물생태계 특성을 파악 하고자 10m×10m(100㎡) 조사구 25개소를 설치하여 주 요환경 인자 및 식생구조 조사를 실시하였다. 상대우점 치를 바탕으로 TWINSPAN classification에 의한 군집 분 석 결과 소나무군집, 소나무-신갈나무군집, 굴참나무군 집, 신갈나무군집, 신갈나무-물박달나무군집, 신갈나무-굴 참나무군집, 일본잎갈나무군집의 7개 유형으로 구분되었 다. 소나무군집, 소나무-신갈나무군집은 천이계열에 있어 소나무 다음 단계라고 할 수 있는 신갈나무로 이행되는 과정에 있었고 굴참나무와 신갈나무가 우점하는 군집은 하층에 차대목인 서어나무, 까치박달 등이 출현하지 않 아 현 상태를 유지할 것으로 판단되었다. 전체출현종수 는 7~15종으로 나타났으며, 각 군집별 평균 출현종수는 5.3~9.7종으로 나타났다. 애미랑재의 종다양도지수는 0.5738~0.8906로 타 지역에 비해 낮은 것으로 분석되었 다. 유사도지수를 분석한 결과 군집별 유사도지수는 11.22~64.86으로 나타났고, 소나무군집과 소나무-신갈나 무군집과의 유사도가 64.86%로 군집간 유사도가 비교적 높게 나타났다. 그러나 신갈나무, 굴참나무 등의 다른 참 나무군집과는 30% 가량의 유사도를 보였다.
본 연구를 바탕으로 낙동정맥 애미랑재 구간 식생분 포 특성 및 식생유형별 천이 경향 분석을 통해 다음과 같은 관리 방안을 제시할 수 있었다. 첫째, 소나무가 우 점하여 소나무 세력이 우세한 군집은 장기적으로 소나무 군집의 유지를 목표로 하여 관리 및 복원 방안을 수립하 도록 한다. 둘째, 소나무와 참나무류 간의 경쟁군집과 참 나무류군집, 일본잎갈나무군집(인공림)은 산림 천이과정 및 생태계 순리에 맞게 자연적 천이가 진행될 수 있게 하며, 인위적인 간섭 및 교란이 발생되지 않도록 조방적 관리 및 복원 방안을 수립한다.
백두대간에 대한 관심은 1990년대 초반 국민적 관심 이 증대되면서 현황조사 및 분석, 관리 및 복원 방안 등 정부, 시민단체, 학계를 중심으로 다양한 연구결과를 도 출한 반면, 지맥인 정맥에 대한 관심은 증가하고 있으나 현황연구는 없는 상태이다. 따라서 본 연구는 백두대간 의 지맥인 낙동정맥을 대상으로 향후 관리 및 복원방안 을 제안하기 위한 기초연구로서 지맥의 연구 방향을 제 안하는데 중요한 역할을 할 수 있을 것이라 기대한다.
본 논문은 농촌진흥청 공동연구사업
(과제번호: PJ00742606)의 지원에 의해 이루어진 것임
참고문헌
1. 국토연구원, 2001, 백두대간의 효율적 관리방안연 구: 관리범위설정을 중심으로, 국토연구원, 2001, 심 포지엄 자료, 64.
2. 국토연구원, 2002a, 백두대간의 효율적 관리방안 연 구 Ⅱ: 관리방안 수립을 중심으로(2차연구), 환경부, 323.
3. 국토연구원, 2002b, 백두대간의 효율적 관리방안연 구: 관리방안 중심(2차연도) 세미나, 97.
4. 국토연구원, 2004, 한반도 산맥체계 재정립 연구:
산줄기 분석을 중심으로, 국토연구원, 229.
5. 국토연구원, 녹색연합, 1999, 백두대간 개념복원과 관리 방향 모색을 위한 심포지엄, 113.
6. 권태호, 이준우, 김동욱, 2004, 백두대간 마루금 등 산로의 훼손실태와 관리방향 :남덕유산-소사고개 구 간을 대상으로, 한국환경생태학회지, 18(2), 175-183.
7. 권태호, 최송현, 유기준, 2002, 백두대간 관리범위 설정에 관한 연구: 유역확장방식에 의한 접근, 한국 지리정보학회지, 5(4), 106-118.
8. 김갑태, 엄태원, 2004, 백두대간 자연생태계의 지역 구분을 위한 식생지수에 관한 연구: 남덕유산 - 소 사고개 구간, 한국환경생태학회지, 18(2), 158-166.
9. 김갑태, 추갑철, 2003, 백두대간 노고단-고리봉 구간 의 식생구조, 한국환경생태학회지, 16(4), 441-448.
10. 김갑태, 추갑철, 백길전, 2004, 백두대간 대덕산-금 대봉 자연생태계 보전지역의 산림군집구조, 한국환 경생태학회지, 17(1), 9-17.
11. 김남춘, 남언정, 신경준, 2008, 백두대간 절토 비탈 면의 생태복원녹화 모델에 관한 연구, 한국환경복 원기술학회지, 10(4), 72-84.
12. 김동필, 최송현, 2004, 낙동정맥 가지산~능동산 구 간의 능선부 식생구조, 한국환경생태학회지, 18(3), 279-287.
13. 김인택, 1998, 가지산의 식물상, 한국생물상연구지, 3. 57-51.
14. 김인택, 송민섭, 이지훈, 1996, 가지산 도립공원의 식물상: Ⅰ, 석남사지역, 창원대학교 환경문제연구 소 논문집, 5, 101-136.
15. 김인택, 송민섭, 이지훈(1997) 가지산 식생에 관한 생태학적 연구, 창원대학교 환경문제연구소 논문집, 6: 43-72.
16. 박상곤, 조현제, 이창배(2009) 백두대간 마루금일대 침엽수림의 식생형과 식물상 조성, 한국임학회지, 98(4): 464-471.
17. 박인협, 이경재, 조재창(1988) 치악산국립공원의 삼 림군집 구조-구룡사-비로봉지역을 중심으로-, 응용 생태연구, 2(1): 1-8.
18. 박인협, 조재창, 오충현(1989) 가야산지역 계곡부와 능선부의 해발고와 사면부위에 따른 삼림구조, 응 용생태연구, 3(1): 42-50.
19. 백재봉, 최송현, 조재우(2000) 석남사 지역의 식생경 관 관리를 위한 삼림구조 분석, 한국정원학회지, 18(4): 68-76.
20. 산림청(1996) 백두대간 관현 문헌집, 21-77.
21. 산림청, 대한지리학회(1997) 백두대간의 산림실태 및 합리적인 보전방안 연구, 263-271쪽.
22. 산림청, 한국환경생태학회(2001) 백두대간 자연생태 계 보전 및 훼손지 복원방안 조사연구, 306쪽.
23. 산림청, 한국환경생태학회(2002) 백두대간 자연생태 계조사 및 관리방안 수립에 관한연구, 279쪽.
24. 산림청, 한국환경생태학회(2003) 백두대간 관리범위 설정 및 관리방안 수립을 위한 연구, 252쪽.
25. 송민섭(1997) 가지산 식생에 관한 생태학적 연구, 창원대학교 석사학위논문, 50쪽.
26. 송유성(2007) 팔공산 파계사의 풍수지리 연구, 영남 대학교 석사학위논문, 115쪽.
27. 신경옥(2004) 낙동정맥 가지산~능동산 구간의 능 선부 식생구조, 밀양대학교 석사학위논문, 45쪽.
28. 신준환(2004) 백두대간 관리범위 및 관리방안, 한국 환경생태학회지, 18(2): 197-204
29. 양보경(1997) 조선시대의 ‘백두대간’ 개념의 형성, 진단학보 83: 85-106.
30. 오구균, 박석곤(2002) 백두대간 피재-도래기재구간 의 능선부 식생구조, 한국환경생태학회지, 15(4):
330-343.
31. 오구균, 이정은(2003) 백두대간의 식물상, 식생, 이 용 및 환경훼손에 관한 학술자료: 지리산 천왕봉부 터 덕유산 향적봉 사이를 중심으로, 한국환경생태 학회지, 16(4) : 475-486.
32. 유기준(2002) 백두대간의 개념적 관리범위모형 수 립을 위한 접근, 한국환경생태학회지, 15(4): 408- 419.
33. 유기준, 권태호, 최송현(2004), 백두대간 지역의 효 율적 자원관리의 방향 :남덕유산~소사고개 구간을 중심으로, 한국환경생태학회지, 15(4): 408-419.
34. 이경재, 구관효, 최재식, 조현서(1991) Classification 및 Ordination 방법에 의한 지리산 대원계곡의 삼림 군집구조 분석, 응용생태연구 5(1): 55-67.
35. 이경재, 민성환, 한봉호(1997) 설악산국립공원 주전
골계곡 식물군집구조 분석, 환경생태학회지 10(2):
283-296.
36. 이경재, 최송현, 조재창(1992) 광릉 삼림의 식물군집 구조(Ⅱ)-Classification 및 Ordination 방법에 의한 죽 엽산 삼림의 식물군집구조 분석-, 한국식물학회지 33(3): 173-182.
37. 이경재, 최송현, 조현서, 이윤원(1994) 덕유산국립공 원의 삼림군집구조분석-백련사-금포탄 지역을 중심 으로-, 응용생태연구 7(1): 35-54.
38. 이동근, 송원경, 전성우, 성현찬, 손동엽(2007), 백두 대간지역의 산림훼손경향 분석, 한국환경복원기술 학회지, 10(4): 41-53.
39. 이지혜, 권혜진, 전기성, 김남춘, 박관수, 송호경 (2008) 백두대간 도로사면의 생태적 녹화를 위한 자 생수종 선정, 한국환경복원기술학회지, 11(4): 67-74.
40. 임동옥, 김용식 박양규, 유은미, 고명희(2004) 백두 대간 덕유산 지역의 식물상, 한국환경생태학회지, 18(2): 107-123
41. 임업연구원(2003) 백두대간 생태계 현황 및 관리범 위설정, 420쪽.
42. 조현서, 이수동(2010) 낙동정맥 삼척시 용소골 계곡 의 식생구조, 한국환경생태학회지, 24(5): 582-590.
43. 조현서, 최송현(2002) 백두대간 태백산지역 백천계 곡의 식물군집구조, 한국환경생태학회지, 15(4):
369-378.
44. 최송현(2002) 백두대간 청옥산지역 능선부의 식물 군집구조, 한국환경생태학회지, 15(4): 344-353.
45. 최송현, 오구균(2003) 백두대간 정령치-복성이재 구 간의 능선부 식생구조, 한국환경생태학회지, 16(4):
421-432.
46. 최송현, 오구균, 강현미(2004) 백두대간 남덕유산-소 사재 구간의 능선부 식생구조. 한국환경생태학회지.
18(2): 131-141.
47. 최송현, 이경재(1993) 북한산국립공원 삼림군집구조 의 5년간 변화 연구, 응용생태연구 7(1): 35-48.
48. 최송현, 조현서, 김보현(2003) 백두대간 정령치-고기 리 구간의 해발고에 따른 식생구조, 한국환경생태 학회지, 16(4): 433-440.
49. 추갑철, 김갑태(2004) 백두대간 수령-소사고개 구간 의 식생구조, 한국환경생태학회지, 18(2): 150-157.
50. 추갑철, 김갑태(2005) 백두대간 부봉-포암산 구간의 식생구조, 한국환경생태학회지, 19(2): 83-89.
51. Brower, J.E. and J.H. Zar(1977) Field and Laboratory Methods for General Ecology. Wm. C. Brown Company, 194pp.
52. Curtis, J.T. and R.P. McIntosh(1951) An upland forest continuum in the prairie-forest border region of Wisconsin. Ecology 32: 476-496
53. Pielou, E.C.(1977) Mathematical ecology. John Wiley&Sons, N.Y. 165pp
54. Whittaker, R.H.(1956) Vegetation of the Great Smoky Mountains Ecological Monographs 26: 1-80.