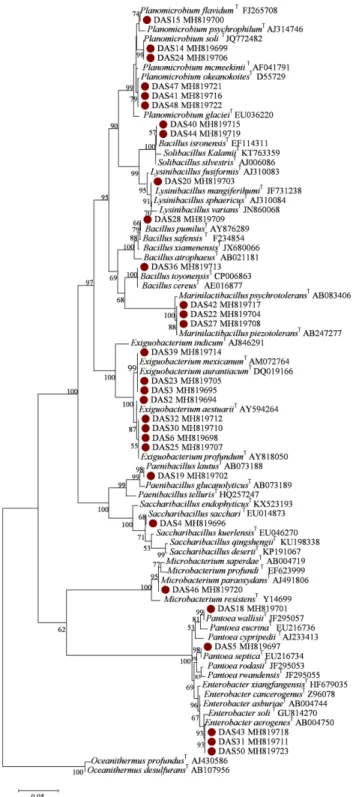
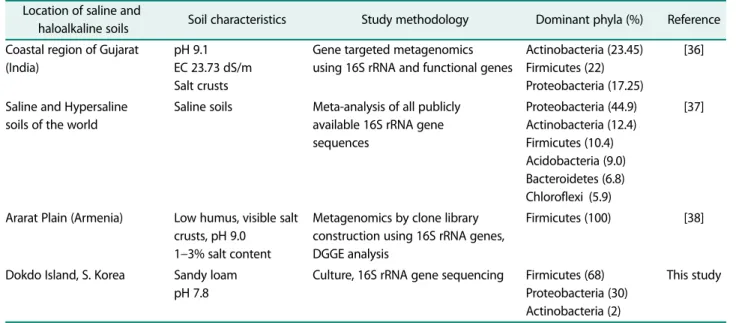
Isolation and Identification of Alkali-tolerant Bacteria from Near-Shore Soils in Dokdo Island
Teddy Namirimu
1, Jinnam Kim
1,2, and Young-Gun Zo
1,2*
1
Department of Biology, Kyungsung University, Busan 48434, Republic of Korea
2
전체 글
1
2
수치
관련 문서
In this thesis, a delay tolerant distance-based broadcasting protocol is proposed, which can change latency according to circumstance of network topology..
Distribution of Cd in stream sediments, paddy soils, rice stalks and grains from the Kubong
Changes in the composition and structure of Mediterranean rokey-shore communities following a gradient of nutrient enrichment: Descriptive study and test
Changes of lactic acid bacteria numbers and photodiode during fermentation mode and storage mode in S kimchi · · ·40..
Days to emergence and heading, duration of growth, leaf traits and stem diameter of sweet sorghum as affected by soils. Soil type Days
Concentration of soil residual endosulfan-sulfate with days after treatment of endosulfan in different soils
Probiotic properties of lactic acid bacteria isolated from mukeunji, a long-term ripened kimchi. Journal of Food
The obligate methanotrophic bacteria Methylococcus, Methylomonas, Methylosinus and related bacteria. Schleifer
