431 http://dx.doi.org/10.4196/kjpp.2014.18.5.431
ABBREVIATIONS: Rb2, Ginsenoside-Rb2; ACh, acetylcholine; Bay- K-8644, methyl-1,4-dihydro-2,6-dimethyl-3-nitro-4-(2-trifluoro-methyl- phenyl)-pyridine-5-carboxylate; CA, catecholamines; DMPP, 1.1- dimethyl-4-phenyl piperazinium iodide; NO, nitric oxide.
Received August 12, 2014, Revised September 3, 2014, Accepted September 16, 2014
Corresponding to: Dong-Yoon Lim, Department of Pharmacology, College of Medicine, Chosun University, 375, Seosuk-dong, Dong-gu, Gwangju 501-759, Korea. (Tel) 82-62-230-6335, (Fax) 82-62-227- 4693, (E-mail) [email protected]
This paper was presented at the 24th Scientific Meeting of the International Society of Hypertension, Athens, Greece, June 13-16, 2014.
This is an Open Access article distributed under the terms of the
Creative Commons Attribution Non-Commercial License (http://
creativecommons.org/licenses/by-nc/3.0) which permits unrestricted non-commercial
use, distribution, and reproduction in any medium, provided the original work
is properly cited.
Inhibitory Effects of Ginsenoside-Rb2 on Nicotinic Stimulation- Evoked Catecholamine Secretion
Hyo-Jeong Lim
1, Hyun-Young Lee
2, and Dong-Yoon Lim
3
1
Department of Internal Medicine (Division of Pulmonary and Critical Care Medicine), Veterans Health Service Medical Center, Seoul 134-791,
2Department of Anesthesiology and Pain Medicine,
3Department of Pharmacology, School of Medicine, Chosun University, Gwangju 501-759, Korea
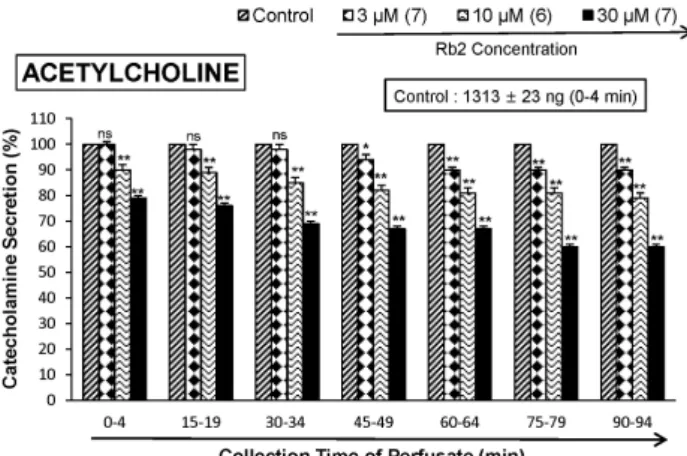
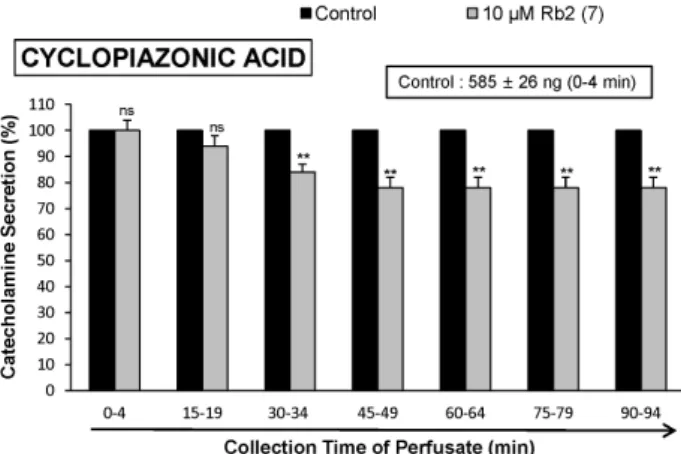
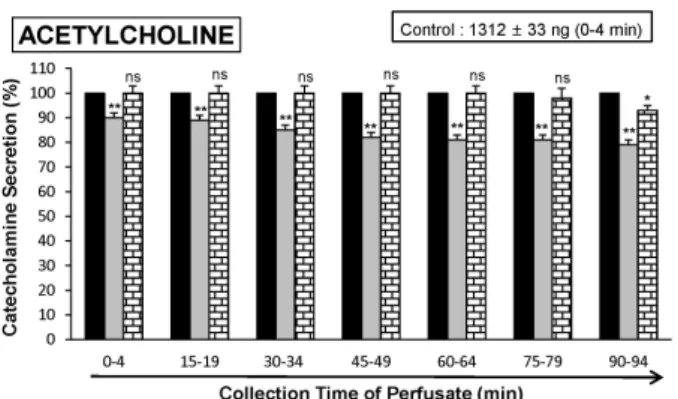
The aim of the present study was to investigate whether ginsenoside-Rb2 (Rb2) can affect the secre- tion of catecholamines (CA) in the perfused model of the rat adrenal medulla. Rb2 (3∼ 30 μ M), per- fused into an adrenal vein for 90 min, inhibited ACh (5.32 mM)-evoked CA secretory response in a dose- and time-dependent fashion. Rb2 (10 μ M) also time-dependently inhibited the CA secretion evoked by DMPP (100 μ M, a selective neuronal nicotinic receptor agonist) and high K
+ (56 mM, a direct membrane depolarizer). Rb2 itself did not affect basal CA secretion (data not shown). Also, in the presence of Rb2 (50 μ g/mL), the secretory responses of CA evoked by veratridine (a selective Na
+
channel activator (50 μ M), Bay-K-8644 (an L-type dihydropyridine Ca
2+ channel activator, 10 μ M), and cyclopiazonic acid (a cytoplasmic Ca
2+-ATPase inhibitor, 10 μ M) were significantly reduced, respectively. Interestingly, in the simultaneous presence of Rb2 (10 μ M) and L-NAME (an inhibitor of NO synthase, 30 μ M), the inhibitory responses of Rb2 on ACh-evoked CA secretory response was considerably recovered to the extent of the corresponding control secretion compared with the inhibitory effect of Rb2-treatment alone. Practically, the level of NO released from adrenal medulla after the treatment of Rb2 (10 μ M) was greatly elevated compared to the corresponding basal released level.
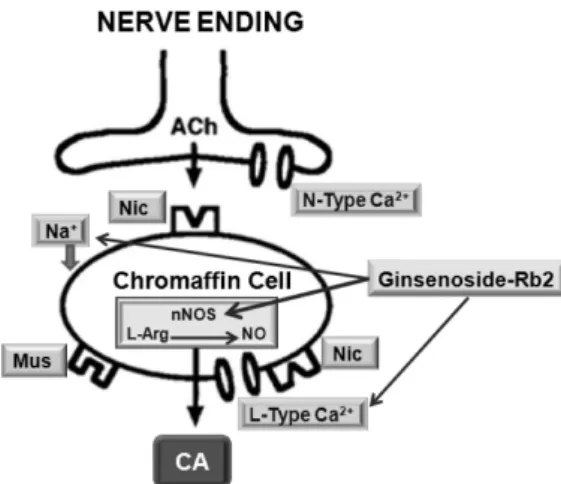
Collectively, these results demonstrate that Rb2 inhibits the CA secretory responses evoked by nicotinic stimulation as well as by direct membrane-depolarization from the isolated perfused rat adrenal medulla. It seems that this inhibitory effect of Rb2 is mediated by inhibiting both the influx of Ca
2+
and Na
+ into the adrenomedullary chromaffin cells and also by suppressing the release of Ca
2+ from the cytoplasmic calcium store, at least partly through the increased NO production due to the activation of nitric oxide synthase, which is relevant to neuronal nicotinic receptor blockade.
Key Words: Adrenal Medulla, Catecholamine secretion, Ginsenoside-Rb2 (Rb2), Nitric oxide synthase (NOS), NO production
INTRODUCTION
Previously, Lim and his coworkers [1-3] have reported that all of total giseng saponin (TGS), panaxadiol and pan- axatriol cause the increased secretion of catecholamines (CA) from the isolated perfused rabbit adrenal glands in
a Ca
2+-dependent fashion, which are mediated by the acti- vation of cholinergic (both nicotinic and muscarinic) re- ceptors and partly the direct action on the rabbit adrenome- dullary chromaffin cells. However, it has been reported that TGS shows the inhibitory effect on the CA release evoked by stimulation of nicotinic receptors in the perfused rat adrenal medulla [4], and also that several ginseng saponins (ginsenosides) from the ginseng root inhibit the CA secre- tion from bovine adrenal chromaffin cells stimulated by ACh [5-8].
In previous study, Tachikawa and his co-workers [6] have
shown that most of the ginsenosides (1∼100 μM) had a
tendency to reduce the ACh-evoked CA secretion. The order
of inhibitory potency (at the concentration of 10 μM) was
as follows: Rg
2
>Rf>Re>Rh1>Rb2, Rg1>Rb1>Rc>Rb3,
Rd, Ro, Rs
1
. The inhibition of ginsenoside-Rg2 at 10 μM
was 72%, but ginsenosides Rb3, Rd, Ro and Rs
1
did not