간암에서 FGFR3 mRNA의 과발현과 FGFR3 유전자의 돌연변이 분석
장영균*,**·배현진*,**·남석우*,**,#
*가톨릭대학교 의과대학 병리학교실, **가톨릭대학교 기능전사체 연구소 (Received September 10, 2012; Revised November 26, 2012; Accepted November 29, 2012)
Overexpression of FGFR3 mRNA and Mutational Analysis of FGFR3 Gene in Hepatocellular Carcinoma
Young Gyoon Chang*,**, Hyun Jin Bae*,** and Suk Woo Nam*,**,#
*Department of Pathology, College of Medicine, The Catholic University of Korea, Seoul 137-701, Korea
**Functional RNomics Research Center, The Catholic University of Korea, Seoul 137-701, Korea
Abstract — FGFR3 is a member of the fibroblast growth factor receptor family which interacts with fibroblast growth fac- tors, setting in motion a cascade of downstream signals, ultimately influencing mitogenesis and differentiation. This par- ticular family member binds acidic and basic fibroblast growth hormone and plays a role in bone development and maintenance. Accumulated evidence suggests that aberrant regulation of FGFR3 and genetic alterations are implicated in the development and progression of various cancers. Despite a high incidence of FGFR3 over-expression, no such inves- tigation has been performed in hepatocellular carcinoma. Thus, we investigated genetic alterations of the FGFR3 gene in 73 cases of hepatocellular carcinoma by single-strand conformational polymorphism (SSCP) and sequencing. One silent mutation (A369A) was found in the extracellular domain of FGFR3, and one genetic alteration in the immunoglobulin-like III domain of FGFR3 appeared to be polymorphism. Taken together, we concluded that over-expression of FGFR3 in hepa- tocellular carcinoma is not associated with genetic alterations of FGFR3 gene, and we suggest that there could be another underlying mechanism of aberrant FGFR3 expression in hepatocellular carcinoma.
Keywords □ FGFR3, overexpression, mutation, hepatocellular carcinoma
간암(Hepatocellular carcinoma, HCC)은 세계적으로 5번째로 발병률이 높은 암종이며, 사망률에 있어서는 2번째로 높은 암종 으로 알려져 있고, 지정학적으로는 아시아와 아프리카에서 발병 률이 높은 것으로 알려져 있다.1-3) B형 간염 바이러스나 C형 간 염 바이러스(Hepatitis B virus, hepatitis C virus), 아플라톡신 B1(aflatoxin B1), 알코올에 대한 과다한 노출 등이 간암을 유발 하는 요소들로 알려져 있지만, 정확한 치료법은 아직 정립되지 않았다.4)여러 가지 종양 억제 유전자의 발현 저해나, ERK/
MAPK, Wnt/β-catenin pathway와 같은 세포 신호전달 체계의 비정상적인 조절이 간암 발생과 진행에 연관되어 있다고 제안되 고 있지만, 간암의 분자병리학적 발암기전은 정확히 밝혀지지 않 았다.5)한편, p53, β-catenin 그리고 AXIN1와 같은 간암유발에 연관된 종양억제유전자(tumor suppressor)나 발암유전자
(oncogene)등의 유전적 변이가 발견되고 있다.6-8)그러나 간암 환 자들에서 이 유전자들의 돌연변이율은 매우 낮게 나타나며 이러 한 유전변이를 통하여 간암 환자 개개인의 임상적 특징을 구별하 는 데는 많은 한계가 따르고 있다. 섬유모세포 성장인자(Fibroblast growth factor receptors, FGFR) 그룹은 FGFR1-4의 네 종류의 단백질로 구성된다. FGFR은 다른 수용체들과 마찬가지로 세포 외 도메인, 막 관통 도메인, 그리고 인산화효소 활성을 갖는 세 포 내 도메인으로 구성된다.9) FGFR의 세포 외 도메인은 2~3개 의 Immunoglobulin과 같은 고리 구조를 가지는데, 이 중 중심부 쪽의 고리 부위에 FGF가 결합하여, 수용체들의 이합체화를 유 도하여 Ras/MAPK, Erk1/2 등을 통한 다양한 신호 전달 과정이 매개된다.10)
여러 종양에서 FGFR3 유전자의 돌연변이가 유전자의 다양한 기능적 부위에 고루 분포되어 있는 것이 알려져 있다. 피부암에 서, FGFR3 유전자의 돌연변이는 7, 10, 12, 15, 19번 Exon 부 위에서 주로 나타난다.11)방광암과 자궁경부암에서 FGFR3의 돌 연변이가 있다는 증거들이 제시되었고, FGFR3의 유전적 변이가
#본 논문에 관한 문의는 저자에게로 (전화) 02-2258-7314 (팩스) 02-537-6585 (E-mail) [email protected]
종설
종양에서 FGFR3의 과발현과 연관성이 있다는 사실이 보고되었 다. 간암에서는 FGFR3가 전사 수준에서 과발현되어 있음이 보고 되어 있지만, FGFR3의 돌연변이와 과발현 간의 연관성에 대해서 는 아직 보고되지 않았다.12)그러므로, 본 연구에서는 간암 조직 에서 FGFR3의 과발현을 다른 그룹의 간암 환자군에서 확인하 고, FGFR3의 유전적 변이와 FGFR3의 과발현 간의 연관성을 조 사하기 위하여 단일쇄 DNA 고차 구조 변화 다형성(single strand conformation polymorphism, SSCP)을 이용하여 73쌍의 간암조 직에서 FGFR3 유전자의 돌연변이를 조사하고자 하였다.
실험방법
조직 시료
삼성서울병원에서 절제술을 받고 진단된 73명의 간암(HCC) 환자의 종양 조직과 정상 조직, 그리고 서울대학교병원에서 얻 어진 13명의 정상인에게서 얻어진 간 조직과 26명의 간암 환자 에게서 얻어진 간암 조직을 대상으로 했다. 가톨릭대학교 의과 대학의 IRB로부터 승인을 받았고 헬싱키선언에 의해 요구되는 원칙에 따라 사전에 동의를 얻었다(CUMC11U010). 삼성서울병 원의 환자 중에서 유전적 요인의 암의 증거는 없었다. 환자 나이 는 26~89세(평균51.6세)이고 남자는 61명 여자는 12명이다. 암 주변 조직은 간경화 50명(68.5%)명 만성 간염 13명(17.8%), 특 정 변화 없음 10명(13.7%). HBV에 감염된 환자는 64명(86.5%) 이었고 HCV에 감염된 환자는 5명(6.8%)으로 검출되었다.
Edmonson grade로 구분하였을 때, grade I은 9명, grade II는 32명, grade III은 32명이었다.
전유전체 미세배열칩
13명의 정상인에서 얻어진 정상 간 조직과 26명의 간암 환자 에게서 얻어진 간암 조직을 이용하여 34,595개의 사람 유전자를 검사할 수 있는 47,224개의 probe를 가진 Sentrix Beadchip array human HT-12-v4-Beadchip(Illumina, San Diego, CA, USA)을 사용하였다. 정상 간 조직과 간암 조직에서 total RNA 를 Trizol(Invitrogen, Carlsbad, CA, USA)을 사용하여 분리하였 다. 분리된 RNA를 Illumina total prep RNA amplification kit (Applied biosystems, Foster City, CA, USA)로 증폭하고, cRNA를 정제하였다. 정제된 cRNA의 순도를 Experion analyzer (Bio-Rad Laboratories, Hercules, CA, USA)를 이용하여 확인한 후, GEX-HYB 용액과 혼합하여 beadchip에 주입하여 58oC에서 18시간 동안 교잡반응을 시켰다. 교잡반응이 끝난 beadchip을 세 척한 후, Block E1 완충액으로 차단하고, streptavidin-Cy3가 포 함된 Block E1완충액으로 염색한 후 세척 및 건조 과정을 거쳤 다. Illumina BeadScanner(Illumina)로 각 bead의 강도(intensity) 를 측정한 후, Illumina BeadStudio 프로그램(Illumina)을 이용해
서 원 자료(raw data)를 추출하였다.
DNA 추출
액체질소를 이용하여 냉동보관된 조직을 곱게 갈아 500 µg/ml 농도의 proteinase K가 포함된 500 µl lysis buffer(5 mM Tris-Cl pH 8.0, 20 mM EDTA, 0.5% Triton X-100)에 넣고 50oC에서 하 루 동안 방치하였다. 그리고 Phenol : chloroform : isoamylalcohol (25 : 24 : 1, Sigma, St. Louis, MO, USA)용액을 각 샘플에 넣고 15,000 rpm으로 4oC에서 30분 동안 원심분리하였다. 이를 한번 반복한 후, 상층액과 동일한 부피의 isopropanol과 1/10 부피의 5 M NaCl을 상층액과 섞고 -70oC에서 1시간 이상 방치하였다.
이를 다시 15,000 rpm으로 4oC에서 30분 동안 원심분리하여, DNA를 침전시켰다. 침전된 DNA를 70% 에탄올로 washing 후 건조시킨 후, 증류수에 녹여 사용하였다.
단일쇄 DNA 고차 구조 변화 다형성(Single strand con- formation polymorphism, SSCP)
간암조직과 쌍을 이루는 정상조직에서 추출한 유전체 DNA(genomic DNA)에서 Exon을 포함하는 8쌍의 Primer를 제 작하였다. 방사성 동위원소인 [32P]dCTP (Amersham, Bucks, UK)를 중합효소연쇄반응에 포함시켜서 자기방사법(autoradiogram) 으로 중합효소연쇄반응 산물을 분석할 수 있게 하였다. 중합효 소연쇄반응(PCRs)은 혼합액을 처음에 95oC에서 12분간 변성시 킨 후 95oC에서 30초, 49~54oC에서 30초, 72oC에서 30초씩 각 각 40회 반복하였으며 72oC에서 5분간 연장반응을 실시하였다.
중합효소연쇄반응 산물을 Single strand conformation poly- morphism(SSCP) 분석을 위해 non-denaturing gel에 running 후 gel을 건조한 후 Kodak X-OMAT X-ray 필름((Eastman Kodak, Rochester, NY, USA)에 감광시켜 관찰하였다. 정상 DNA에서 관 찰되는 wild-type band 이외의 band가 나타난 경우, 2회 이상 반 복하여 확인하고, aberrant band를 잘라서 코스모진텍(Seoul, Korea)에 sequencing을 의뢰하여 염기 서열의 변화를 확인하였다.
실험결과 및 고찰
최근의 여러가지 연구들에서, FGFR3 유전자의 돌연변이가 종 양에서 FGFR3 단백질의 과발현이나 종양 진행 단계에 관여한 다는 것이 제시되었다. 방광암(bladder cancer)에서 7, 10, 15번 Exon 부위에서 64%의 비율로 FGFR3의 돌연변이가 발견되었 다.13)또한, 요르단인 방광암 환자 그룹에서는 R278C, S249C, Y375C, G382R 등의 돌연변이가 각각 10%, 59%, 18%, 그리고 10%의 확률로 나타났으며, 이러한 돌연변이는 이른 단계나 낮은 단계의 방광암 환자 그룹에서의 FGFR3 단백질의 과발현과 높 은 상관관계가 있는 것으로 나타났다.14)FGFR3 유전자의 S248C
돌연변이를 가지는 방광암 세포주인 97-7 세포주를 이용한 연구 에서는 돌연변이가 일어나는 유전자 발현을 선택적으로 억제하 였을 때, 암세포주의 성장이 현저히 억제되는 것이 발견되었다.15) 이러한 결과들은 FGFR3 유전자의 유전적 변이에 의해 종양에 서 FGFR3가 과발현되고, 이를 통해 FGFR3를 통한 신호전달체 계가 종양에서 활성화되어, 종양 형성에 기여할 수 있다는 가능 성을 제시하고 있다.
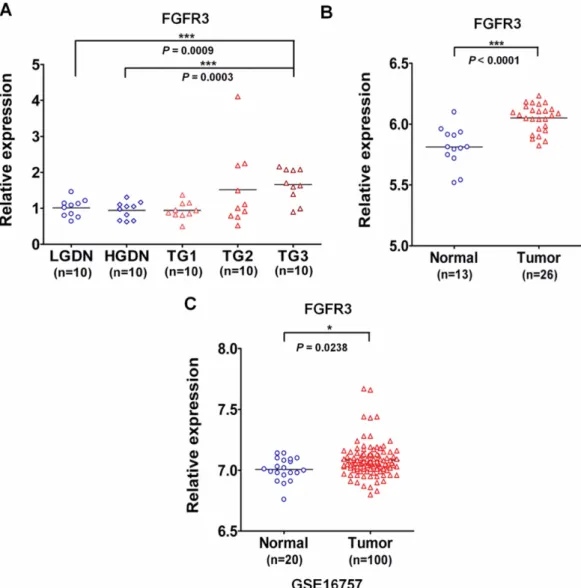
이러한 선행 연구 결과들을 토대로, 본 연구에서는 간암 조직 에서 FGFR3의 발현 양상을 조사하였다. 먼저, 간암 조직에서의 FGFR3의 mRNA 과발현을 확인하기 위하여 본 연구진의 선행 연구 결과로부터 FGFR3의 발현을 분석하였다.16) Fig. 1A에서
볼 수 있듯이 간암 병기가 진행될수록 FGFR3의 mRNA 발현이 증가하는 것을 확인하였다. 간암에서 FGFR3의 mRNA 과발현 을 다른 환자군에서 재확인하기 위하여 13명의 정상인에게서 얻 어진 간 조직과 26명의 간암 환자에서 얻어진 간암 조직을 이용 하여 DNA microarray를 수행하여 FGFR3의 probe의 신호 강 도를 분석하였다. 그 결과 정상인의 간 조직과 비교하여 간암 환 자의 종양 조직에서 FGFR3의 mRNA의 발현이 현저히 증가되 는 것을 확인하였다(Fig. 1B). 이러한 사실을 대단위 환자군에서 얻어진 데이터와 비교하기 위하여 NCBI(National Center for Biotechnology Information)의 GEO(Gene expression omnibus) 데이터를 이용하여 FGFR3의 간암에서의 발현을 분석하였다
Fig. 1− Over-expression of FGFR3 mRNA in hepatocellular carcinoma. (A) Analysis of FGFR3 expression from the previously reported large- scale gene expression analysis. FGFR3 is aberrantly over-expressed in tumor grade 3, compared to non-cancerous lesions, LGDN (Low-grade dysplastic nodule, ***P=0.009) and HGDN (High-grade dysplastic nodule, ***P=0.0003, unpaired t-test). (B) Analysis of FGFR3 expression from the large-scale gene expression analysis with 13 normal liver tissues and 26 tumor tissues. The expression level of FGFR3 is significantly elevated in tumor tissues compared to normal tissues (P<0.0001, unpaired t-test). (C) Analysis of FGFR3 expression from the large-cohort GEO dataset, GSE16757. FGFR3 mRNA expression was significantly up-regulated in tumor patients (*P=0.0238, unparied t-test).
(GSE16757). 그 결과 대단위 환자군에서도 FGFR3의 mRNA발 현이 증가되는 것을 확인하였다(Fig. 1C). 이러한 분석 결과를 토 대로 FGFR3 유전자의 mRNA가 정상 조직과 비교해서 간암 종 양 조직에서 과발현되어 있다는 사실을 확인하였다. 이러한 결 과들은 FGFR3의 mRNA의 발현이 미국인 간암 환자에서 증가 되어 있다는 다른 논문의 결과와도 일치한다.12)한편, 종양 억제 효과를 갖는 vitamin K가 Huh-7 간암세포주에서 FGFR3의 mRNA의 발현을 감소시켜, FGFR3-ERK1/2 신호전달체계를 억 제함으로써 암세포주의 성장을 억제한다는 결과가 보고되었다.17) 이러한 연구 결과들은 간암에서도 FGFR3가 전사단계에서 과발 현되며, FGFR3에 의한 신호전달체계가 간암에서 활성화되어 종 양 발생과 진행에 관여할 수 있음을 시사한다.
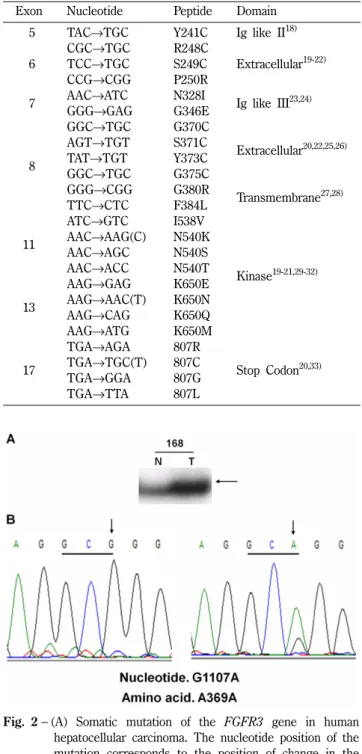
FGFR3 유전자의 유전적 돌연변이들은 여러 부위의 기능적 도 메인 (functional domain)에서 발견되어 있다(Table I). 따라서, 본 연구에서는 간암에서 FGFR3의 과발현과 FGFR3 유전자의 돌 연변이의 상관관계를 알아보기 위하여 다른 암종에서 발견된 돌 연변이를 포함하는 5, 6, 7, 8, 11, 13, 그리고 17번 Exon에서 FGFR3 유전자의 돌연변이를 조사하였다. 간암에서 유전적 변이 가 FGFR3의 과발현에 미치는 영향을 조사하기 위하여, 다른 몇 몇 연구들에서 FGFR3 유전자의 돌연변이가 발견된 exon 5, 6, 7, 8, 11, 13, 17 부위를 PCR로 증폭한 후 SSCP법을 이용하여 돌연변이를 분석하였다. 그 결과 73명의 간암 환자 중 다섯 명 의 환자에서 두 가지의 유전적 변이가 검출되었다. Exon 7번에 서는 정상 조직과 간암 조직에서 모두 SSCP상에서의 비정상적 인 band가 나타났으며, 이를 sequencing을 통하여 확인한 결과 990번째 염기인 시토신(cytosine)이 티민(thymine)으로 변하였지 만, 아미노산은 티로신(tyrosine)으로 그대로 유지되는 synony- mous SNP(single nucleotide polymorphism)임이 확인되었다 (Table II). 또한 exon 8번에서는 한 명의 환자의 간암 조직에서
만 SSCP상에서의 비정상적인 band가 확인되었으며, sequencing 을 통하여 1107번째 염기인 구아닌(guanine)이 아데닌(adenine) Table I− Primer sequences used in this study
Name of
primer Nucleotide sequence Product
size (bp) E5F 5'-CGCATTGGAGGCATCAAG-3'
E5R 5'-TTCTCCACGACGCAGGTGT-3' 151 E6-1F 5'-CCCCTGAGCGTCATCTG-3' E6-1R 5'-CACCTTGCTGCCATTCAC-3' 236 E6-2F 5'-TACAGTGACGCACAGCCCCACAT-3' E6-2R 5'-GACCCAAATCCTCACGCAACCC-3' 229
E7F 5'-CGCCTATCGCTCTGCTCTC-3' E7R 5'-CGTCCCCCAGACAGTGC-3' 219 E8F 5'-CCAACGCCCATGTCTT-3' E8R 5'-GCTCAGAACCTGGTATCT-3' 241
E11F 5'-AGCCGTGAAGATGCTGAAAGGTGAGG-3' E11R 5'-CACCGCCGCTACCGCACCTAC-3' 234 E13F 5'-GGGGAGAGGTGGAGAGGCTTCA-3' E13R 5'-GGTGTGGGAAGGCGGTGTTG-3' 216 E17F 5'-AGAGGTGGCTGTGCGAAGA-3' E17R 5'-CTCACATTGTTGGGGACCAGT-3' 235
Table II− Previously reported mutations of FGFR3 gene Exon Nucleotide Peptide Domain
05 TAC→TGC Y241C Ig like II18) 06
CGC→TGC R248C
Extracellular19-22)
TCC→TGC S249C
CCG→CGG P250R
07 AAC→ATC N328I
Ig like III23,24)
GGG→GAG G346E
08
GGC→TGC G370C
Extracellular20,22,25,26)
AGT→TGT S371C
TAT→TGT Y373C
GGC→TGC G375C
GGG→CGG G380R
Transmembrane27,28)
TTC→CTC F384L
11
ATC→GTC I538V
Kinase19-21,29-32)
AAC→AAG(C) N540K
AAC→AGC N540S
AAC→ACC N540T
13
AAG→GAG K650E
AAG→AAC(T) K650N
AAG→CAG K650Q
AAG→ATG K650M
17
TGA→AGA 807R
Stop Codon20,33) TGA→TGC(T) 807C
TGA→GGA 807G
TGA→TTA 807L
Fig. 2− (A) Somatic mutation of the FGFR3 gene in human hepatocellular carcinoma. The nucleotide position of the mutation corresponds to the position of change in the coding sequence of the gene, where position 1 represents the A of the ATG. (Genbank Accession NM_000142.4).
SSCP of DNAs obtained from cancer cells showed one aberrantly migrating band (arrowed) as compared with corresponding normal cells, indicating a genetic alteration in one allele. (B) Sequencing analysis identified the mutation to involve a GCG to GCA (A369A) change in exon 8 of the FGFR3 gene.
으로 변한 것이 확인되었다(Fig. 2A and 2B). 이 돌연변이 역시 아미노산은 알라닌(alanine)으로 그대로 유지되는 silent mutation 임이 판명되었다(Table II, Fig. 2). 이러한 결과를 토대로 간암 환자군에서 FGFR3의 유전적 변이는 매우 낮은 확률로 발생하 며, 유전적 변이에 의한 아미노산의 변화가 없기 때문에, 실험에 사용된 환자군에서 FGFR3의 과발현이 유전적 변이와는 밀접한 관계가 없다는 결론을 도출하였다.
결 론
본 연구는 간암 환자의 정상 조직과 종양 조직에서, 그리고 정 상인의 간 조직과 간암 환자의 종양 조직에서 FGFR3의 mRNA 의 발현 양상을 비교하고, FGFR3의 유전적 변이가 FGFR3의 발현 양상과의 상관 관계가 있는가를 조사하였다. 실험 결과 FGFR3의 mRNA의 발현이 정상 조직과 비교하여 간암 종양 조 직에서 매우 증가되어 있음을 확인하였다. 하지만 돌연변이 조 사 결과 극히 빈도가 낮은 C990T, G1107A의 두 종류의 염기 서 열 변화가 나타났으며, 이 중 C990T는 정상 조직과 종양 조직 에서 동일하게 나타나는 SNP로 판명되었으며, G1107A는 종양 조직에서만 나타났다. 두 유전적 변이에서 유래되는 아미노산 변 화는 T330T, A369A로서, 아미노산 자체는 변화하지 않음을 알 게 되었다. 이는 간암에서 FGFR3의 과발현이 유전적 변이에 의 한 것이 아니라 다른 요인에 의한 것임을 시사하며, FGFR3에 의한 신호전달체계가 간암 발생과 진행에 어떻게 연관되어 있는 지에 대해서는 좀 더 많은 연구가 필요하다.
감사의 말씀
본 연구는 기초의과학분야 선도연구센터인, "가톨릭 암진화연 구센터(2012047939)" 연구지원으로 수행되었으며 이에 감사 드 립니다.
참고문헌
1) Song, J., Kim, C. J., Cho, Y. G., Kim, S. Y., Nam, S. W., Lee, S. H., Yoo, N. J., Lee, J. Y. and Park, W. S. : Genetic and epigenetic alterations of the KLF6 gene in hepatocellular
carcinoma. Journal of Gastroenterology and Hepatology 21, 1286 (2006).
2) Villanueva, A., Newell, P., Chiang, D. Y., Friedman, S. L. and Llovet, J. M. : Genomics and signaling pathways in hepatocellular carcinoma. Seminars in Liver Disease. 27, 55 (2007).
3) Sahadevan, K., Darby, S., Leung, H. Y., Mathers, M. E., Robson, C. N. and Gnanapragasam, V. J. : Selective over- expression of fibroblast growth factor receptors 1 and 4 in clinical prostate cancer. J. Pathol. 213, 82 (2007).
4) Jemal, A., Bray, F., Center, M. M., Ferlay, J., Ward, E. and Forman, D. : Global cancer statistics. CA Cancer J. Clin. 61, 69 (2011).
5) Whittaker, S., Marais, R. and Zhu, A. X. : The role of signaling pathways in the development and treatment of hepatocellular carcinoma. Oncogene. 29, 4989 (2010).
6) de La Coste, A., Romagnolo, B., Billuart, P., Renard, C. A., Buendia, M. A., Soubrane, O., Fabre, M., Chelly, J., Beldjord, C., Kahn, A. and Perret, C. : Somatic mutations of the beta- catenin gene are frequent in mouse and human hepatocellular carcinomas. Proceedings of the National Academy of Sciences of the United States of America. 95, 8847 (1998).
7) Pang, A., Ng, I. O., Fan, S. T. and Kwong, Y. L. : Clinicopathologic significance of genetic alterations in hepatocellular carcinoma. Cancer Genetics and Cytogenetics 146, 8 (2003).
8) Satoh, S., Daigo, Y., Furukawa, Y., Kato, T., Miwa, N., Nishiwaki, T., Kawasoe, T., Ishiguro, H., Fujita, M., Tokino, T., Sasaki, Y., Imaoka, S., Murata, M., Shimano, T., Yamaoka, Y.
and Nakamura, Y. : AXIN1 mutations in hepatocellular carcinomas, and growth suppression in cancer cells by virus- mediated transfer of AXIN1. Nature Genetics. 24, 245 (2000).
9) McKeehan, W. L., Wang, F. and Kan, M. : The heparan sulfate- fibroblast growth factor family: diversity of structure and function. Prog. Nucleic. Acid Res. Mol. Biol. 59, 135 (1998).
10) Eswarakumar, V. P., Lax, I. and Schlessinger, J. : Cellular signaling by fibroblast growth factor receptors. Cytokine Growth Factor Rev. 16, 139 (2005).
11) Hafner, C., Vogt, T. and Hartmann, A. : FGFR3 mutations in benign skin tumors. Cell Cycle. 5, 2723 (2006).
12) Qiu, W. H., Zhou, B. S., Chu, P. G., Chen, W. G., Chung, C., Table III− Genetic alterations of FGFR3 in hepatocellular carcinoma
Exon No. of tissues Nucleotide Codon Amino acid Type
7
14N, 14T, 16N, 16T, 33N, 33T, 34N, 34T
C990T ACC→ACT T330T SNP (Synonymous)
8 168T G1107A GCG→GCA A369A Mutation (Silent)
Shih, J., Hwu, P., Yeh, C., Lopez, R. and Yen, Y. : Over- expression of fibroblast growth factor receptor 3 in human hepatocellular carcinoma. World J. Gastroenterol. 11, 5266 (2005).
13) Kompier, L. C., Lurkin, I., van der Aa, M. N., van Rhijn, B. W., van der Kwast, T. H. and Zwarthoff, E. C. : FGFR3, HRAS, KRAS, NRAS and PIK3CA mutations in bladder cancer and their potential as biomarkers for surveillance and therapy.
PLoS One. 5, e13821 (2010).
14) Bodoor, K., Ghabkari, A., Jaradat, Z., Alkhateeb, A., Jaradat, S., Al-Ghazo, M. A., Matalka, I., Musleh, H. and Haddad, Y. : FGFR3 mutational status and protein expression in patients with bladder cancer in a Jordanian population. Cancer Epidemiol. 34, 724 (2010).
15) Tomlinson, D. C., Hurst, C. D. and Knowles, M. A. : Knockdown by shRNA identifies S249C mutant FGFR3 as a potential therapeutic target in bladder cancer. Oncogene. 26, 5889 (2007).
16) Nam, S. W., Park, J. Y., Ramasamy, A., Shevade, S., Islam, A., Long, P. M., Park, C. K., Park, S. E., Kim, S. Y., Lee, S. H., Park, W. S., Yoo, N. J., Liu, E. T., Miller, L. D. and Lee, J. Y. : Molecular changes from dysplastic nodule to hepatocellular carcinoma through gene expression profiling. Hepatology 42, 809 (2005).
17) Cao, K., Liu, W., Nakamura, H., Enomoto, H., Yamamoto, T., Saito, M., Imanishi, H., Shimomura, S., Cao, P. and Nishiguchi, S. : Vitamin K2 downregulates the expression of fibroblast growth factor receptor 3 in human hepatocellular carcinoma cells. Hepatol Res. 39, 1108 (2009).
18) Onwuazor, O. N., Wen, X. Y., Wang, D. Y., Zhuang, L., Masih- Khan, E., Claudio, J., Barlogie, B., Shaughnessy, J. D., Jr. and Stewart, A. K. : Mutation, SNP, and isoform analysis of fibroblast growth factor receptor 3 (FGFR3) in 150 newly diagnosed multiple myeloma patients. Blood. 102, 772 (2003).
19) Cappellen, D., De Oliveira, C., Ricol, D., de Medina, S., Bourdin, J., Sastre-Garau, X., Chopin, D., Thiery, J. P. and Radvanyi, F.
: Frequent activating mutations of FGFR3 in human bladder and cervix carcinomas. Nat Genet. 23, 18 (1999).
20) Rousseau, F., el Ghouzzi, V., Delezoide, A. L., Legeai-Mallet, L., Le Merrer, M., Munnich, A. and Bonaventure, J. : Missense FGFR3 mutations create cysteine residues in thanatophoric dwarfism type I (TD1). Hum Mol Genet. 5, 509 (1996).
21) Sibley, K., Cuthbert-Heavens, D. and Knowles, M. A. : Loss of heterozygosity at 4p16.3 and mutation of FGFR3 in transitional cell carcinoma. Oncogene. 20, 686 (2001).
22) Tavormina, P. L., Rimoin, D. L., Cohn, D. H., Zhu, Y. Z., Shiang, R. and Wasmuth, J. J. : Another mutation that results in the substitution of an unpaired cysteine residue in the extracellular domain of FGFR3 in thanatophoric dysplasia type I. Hum Mol.
Genet. 4, 2175 (1995).
23) Winterpacht, A., Hilbert, K., Stelzer, C., Schweikardt, T., Decker, H., Segerer, H., Spranger, J. and Zabel, B. : A novel mutation in FGFR-3 disrupts a putative N-glycosylation site and results in hypochondroplasia. Physiol. Genomics. 2, 9 (2000).
24) Park, W. J., Bellus, G. A. and Jabs, E. W. : Mutations in fibroblast growth factor receptors: phenotypic consequences during eukaryotic development. Am. J. Hum. Genet. 57, 748 (1995).
25) Hafner, C., van Oers, J. M., Vogt, T., Landthaler, M., Stoehr, R., Blaszyk, H., Hofstaedter, F., Zwarthoff, E. C. and Hartmann, A. : Mosaicism of activating FGFR3 mutations in human skin causes epidermal nevi. J. Clin. Invest. 116, 2201 (2006).
26) Superti-Furga, A., Eich, G., Bucher, H. U., Wisser, J., Giedion, A., Gitzelmann, R. and Steinmann, B. : A glycine 375-to- cysteine substitution in the transmembrane domain of the fibroblast growth factor receptor-3 in a newborn with achondroplasia. Eur. J. Pediatr. 154, 215 (1995).
27) Rousseau, F., Bonaventure, J., Legeai-Mallet, L., Pelet, A., Rozet, J. M., Maroteaux, P., Le Merrer, M. and Munnich, A. : Mutations in the gene encoding fibroblast growth factor receptor-3 in achondroplasia. Nature 371, 252 (1994).
28) Shiang, R., Thompson, L. M., Zhu, Y. Z., Church, D. M., Fielder, T. J., Bocian, M., Winokur, S. T. and Wasmuth, J. J. : Mutations in the transmembrane domain of FGFR3 cause the most common genetic form of dwarfism, achondroplasia. Cell.
78, 335 (1994).
29) Deutz-Terlouw, P. P., Losekoot, M., Aalfs, C. M., Hennekam, R. C. and Bakker, E. : Asn540Thr substitution in the fibroblast growth factor receptor 3 tyrosine kinase domain causing hypochondroplasia. Hum Mutat. Suppl 1, S62 (1998).
30) Grigelioniene, G., Hagenas, L., Eklof, O., Neumeyer, L., Haereid, P. E. and Anvret, M. : A novel missense mutation Ile538Val in the fibroblast growth factor receptor 3 in hypochondroplasia. Mutations in brief no. 122. Online. Hum Mutat. 11, 333 (1998).
31) Mortier, G., Nuytinck, L., Craen, M., Renard, J. P., Leroy, J. G.
and de Paepe, A. : Clinical and radiographic features of a family with hypochondroplasia owing to a novel Asn540Ser mutation in the fibroblast growth factor receptor 3 gene. J. Med. Genet.
37, 220 (2000).
32) Prinos, P., Costa, T., Sommer, A., Kilpatrick, M. W. and Tsipouras, P. : A common FGFR3 gene mutation in hypochondroplasia. Hum Mol Genet. 4, 2097 (1995).
33) Rousseau, F., Saugier, P., Le Merrer, M., Munnich, A., Delezoide, A. L., Maroteaux, P., Bonaventure, J., Narcy, F. and Sanak, M. : Stop codon FGFR3 mutations in thanatophoric dwarfism type 1. Nat. Genet. 10, 11 (1995).