Y. J. Jung ・ K. K. Kang ( )
Institute of Genetic Engineering, Hankyong National University, Ansung 456-749, Korea
e-mail: [email protected]
I. H. Lee ・ K. H. Han ・ C. Y. Son ・ K. K. Kang
Department of Horticulture, Hankyong National University, Ansung, 456-749, Korea
Y. G. Cho
Department of Crop Science, Chungbuk National University, Cheongju, Korea
M. C. Lee ( )
National Agrobiodiversity Center, National Academy of Agricultural Science, RDA, 88-20, Suwon, 441-707, Korea
Expression analysis and characterization of rice oligopeptide transport gene (OsOPT10) that contributes to salt stress tolerance
Yu Jin Jung ・ In Hye Lee ・ Kyung Hee Han ・ Cho Yee Son ・ Yong Gu Cho ・ Myung Chul Lee ・ Kwon Kyoo Kang
Received: 2 September 2010 / Accepted: 16 September 2010
ⓒKorean Society for Plant Biotechnology
Abstract Knock-out of a gene by insertional mutagenesis is a direct way to address its function through the mutant phenotype. Among ca. 15,000 gene-trapped Ds insertion lines of rice, we identified one line from selected sensitive lines in highly salt stress. We conducted gene tagging by TAIL-PCR, and DNA gel blot analysis from salt sensitive mutant. A gene encoding an oligopeptide transporter (OPT family) homologue was disrupted by the insertion of a Ds transposon into the OsOPT10 gene that was located shot arm of chromosome 8. The OsOPT10 gene (NP_ 001062118.) has 6 exons and encodes a protein (752 aa) containing the OPT family domain. RT-PCR analysis showed that the expression of OsOPT10gene was rapidly and strongly induced by stresses such as high-salinity (250 mM), osmotic, drought, 100 µM ABA. The subcellular localization assay indicated that OsOPT10 was localized specifically in the plasma mem- brane. Overexpression of OsOPT10 in Arabidopsis thaliana and rice conferred tolerance of transgenic plants to salt stress.
Further we found expression levels of some stress related genes were inhibited in OsOPT10 transgenic plants. These results suggested that OsOPT10 might play crucial but differential roles in plant responses to various abiotic stresses
Keywords Arabidopsis, Ds insertion mutant, GFP, OPT family, rice, salt stress, transgenic plant
Introduction
Rice (Oryza sativa L.), one of the most important crops, is an ideal model species for the study of crop genes due to its commercial value, relatively small genome size (ca.430 Mb), diploid origin (2x = 24) and close relationship to other important cereal crops. Insertional mutagenesis based on T-DNA and maize Activator/Dissociator (Ac/Ds) transposons is most suitable for a systematic functional analysis of multifamily plant genes. In the previously studies, we used the Activator/Dissociation (Ac/Ds) two- component transposon system (Han 2002) in rice to prepare a large collection of gene-trapped Ds insertion lines of rice carrying a single Ds insertion (Chin et al.
1999; Kim et al. 2002, 2004). To assessing the potential of reverse genetics to identify insertional mutants in salt stress, we screened salt adaptation mutants from Ds-tagged rice lines. One of the most efficient ways to achieve the goal of producing more rice is to develop different varieties that are high seed yielding, biotic and abiotic stress resistant by modern breeding technology.
One of the abiotic stress factor, high salinity influence plant growth and development, and are major limit factor on crop productivity (Teige et al. 2004; Bray 2004; Yun 2005). Salt stress, mostly caused by excess of NaCl, elicits two primary effects on plants such as osmotic stress and ionic toxicity due to high levels of Na+ and/or Cl- ions in the cytoplasm (Greenway & Munns 1980;
Zhu 2001). One of the early responses of plant cells to many stresses, including salt stress, is the generation of a cytosolic Ca2+increase. In plant cells, Ca2+ serves as a second messenger during stress signaling and it activated the signal delivery system against the stress of DOI:10.5010/JPB.2010.37.4.483
Research Article
the cell (Sanders,Brownlee & Harper, 1999). It was in control of influx of K+ ions and relationship of K+/Na+ which participate to salt stress adaptation of the cell (Maeda et al. 1994; Yun 2005). However, at high NaCl condition, Na+ competition at binding sites for K+ may result in K+ deficiency (Maathuis & Amtmann 1999).
Knockout of a gene by insertional mutagenesis is a direct way to address its function through the mutant phenotype. Among ca. 15,000 gene-trapped Ds insertion lines of rice (data not shown), we identified one line from selected sensitive lines in highly salt stress that were closely linked to insertion sites of the Ds element, and termed salt sensitive mutant which has a disrupted oligopeptide transporters (OsOPT10) gene. Peptide transport is characterized by the ability of cells to transport peptides or peptide derivatives across membranes in an energy-dependent manner (Lubkowitz et al. 1997;
Stacey et al. 2006). Peptide transporters can be classified into three groups, namely the ATP-binding cassette transporters (ABC family), the peptide transporters (PTR family) and the oligopeptide transporters (OPT family) (Stacey et al. 2002, 2006). The OPT family was first characterized in the pathogenic yeast Candida Albicans and likely use the proton gradient to energize transport (Lubkowitz et al. 1997; Koh et al. 2002; Stacey et al.
2002, 2006). Database searches for homology with the C. albicans CaOpt1p revealed nine putative Arabidopsis OPT orthologs that form a distinct subfamily compared with the fungal OPTs. Both of these motifs (i.e. NPG motif, NPG[P/A]F[N/T/S]XKEH[V/T/A][L/I/V][I/V]I[T/
S/V][I/V/M][F/M][A/S][N/S/A] and KIPPR motif, K[L/F]
[G/A][H/M/T]YMK[I/V/L][P/D/S] PR) were found in regions of the protein predicted to be hydrophilic. Members of the PTR family transport dipeptides and tripeptides as well as amino acids andnitrate (reviewed by Williams &
Miller 2001; Stacey et al. 2002a). PTR proteins have been identified in yeast (Perry et al. 1994), human (Liang et al. 1995), rabbit (Fei et al. 1994, 2000), mouse (Ganapathy et al. 1995), barley (West et al. 1998), and Arabidopsis (Frommer et al. 1994; Steiner et al. 1994).
In contrast to PTR peptide transporters, the yeast OPT transporters predominantly recognize tetra- and pentape- ptides (Lubkowitz et al. 1997; Lubkowitz et al. 1998), including the endogenous opioids Met-enkephalin (YG GFM) and Leu-enkephalin (YGGFL) (Hauser et al.
2000). Database searches for homology with the C.
albicans CaOpt1p revealed nine putative Arabidopsis OPT orthologs that form a distinct subfamily compared with the fungal OPTs (Koh et al. 2002; Stacey et al.
2002). Full-length cDNAs for seven Arabidopsis OPT transporters (AtOPT1 to AtOPT7) were cloned and tested for their ability to take up various peptides when expressed in yeast (Koh et al. 2002). More recently, OsGT1 form Oryza sativa (Zhang et al. 2004), and AtOPT6 from Arabidopsis (Cagnac et al. 2004) have been reported to enable the uptake of glutathione (GSH and GSSG) and glutathione derivatives (GS conjugates) when expressed in yeast. The OPT family of transporters are larger and more diverse than was previously recognized. However, no OPT transporter has yet been identified in animals (Stacey et al. 2006). Until now in plants, unique expression patterns were also found for selected AtOPTs such as expression in pollen tubes, ovules, endosperm, integument of developing seeds and tissue-specific expression pattern (Stacey et al. 2006;
Koh et al. 2002). However, the exact role of peptide transport in plants is still undefined. In this study, we carried out physiological analysis on the selected salt sensitive and expression pattern OsOPT10 gene. Also, we will find expression pattern and gene function of OsOPT10 in transgenic Arabidopsis and rice.
Materials and methods
Plant growth and stress treatment
We used japonica rice cultivar (cv.) Dongjin from Activator/
Dissociation (Ac/Ds) two-component transposon system (Han 2002) in rice to prepare a large collection of gene- trapped Ds insertion lines of rice carrying a single Ds insertion (Kim et al. 2002, 2004; Chin et al. 1999). Rice seeds were germinated for 20 days under normal conditions at 28 ºC and 14 h light / 10 h darkness in soil in a green house. For high salt stress treatment, 20-day-old (three or four-leaf-old) rice seedlings were grown in nutrient solution containing 250 mM NaCl, and imbibed for 14 days under a normal light/dark cycle at 28 ºC Plants were measured for fresh weight according to Xiang et al.
(2007).
Isolation of Ds-flanking sequences by TAIL-PCR
Analysis of TAIL-PCR and DNA gel blot was carried out as described by Kim et al. (2004). The sequence of FSTs (franking sequencing tags) was analyzed online at the following website: http://www.ncbi.nlm.nih.gov and
http://www.gramene.org. The insertion site of Ds element was confirmed by PCR analysis between Ds and OsOPT10 gene region. The Ac/Ds PCR analysis and Southern blot were carried out as described by Kim et al. (2004).Total RNA was isolated with easy-spin plant RNA extraction kit (iNtRON Biotechnology, South Korea) according to the manufacturer’s manual. The first-strand cDNA was gene- rated by using SuperscriptTM III Reverse Transcriptase according to the manufacturer’s instructions (Invitrogen, USA). Based on the flanking sequence of OsOPT10 gene, the 2.25 kb full-length gene primer set Fw (5′-ATGGA GCAACCACGGCT TGA-3′) and Rv (5′-TCACTGCA CCGGGCAGC-3′) was designed and synthesized. The amplification program consisted of an initial step at 94°C for 2 min, 30 cycles (94°C for 15 s, 58°C 30 s, 72°C for 1 min), and a final step at 72°C 10 min. The PCR product was ligated into the pGEM-T easy vector (Promega, USA) and sequenced.
Phylogenetic analysis
Identification of nucleotide sequence was established by comparing with the GRAD Rice Genome Annotation Database (Sasaki et al. 2002, http://rgp.dna.affrc.go.jp/
RiceHMM/). All of the OPT sequences were compared using MegAlign (Version 7.0, DNASTAR Inc.). This led to the identification of two conserved motifs found in all OPT proteins. The bioinformatics tools at the web site http://www.expasy.org were used to analyze the deduced protein. A phylogenetic tree was constructed using at the web site http://www.phylogeny.fr/.
The Arabidopsis orthologs were identified by comparison of the CaOpt1p sequence (Lubkowitz et al. 1997). For phylogenetic analysis, OPT family members in Arabidopsis OPT orthologs gene sequences were retrieved from GenBank database (Koh et al. 2002). Using DNASTAR program, overall amino acid sequences were aligned, and then the unrooted phylogenetic tree was generated. This tree was set up with the distance matrix using the neighbor-joining method. Poisson correction with the complete deletion of gaps was used to calculate protein distances. Bootstrap values are based on 100 iterations.
Gene expression analysis under the abiotic stress treatments Seeds of rice (Oryza sativa) var. Dongjin were grown under controlled conditions having 28°C day / 25°C night temperatures, 14 h light / 10 h darkness in Yosida solution (Yosida et al. 1976), and 75% relative humidity.
After 2 weeks of germination, seedlings were exposed to high-salinity (250 mM), osmotic (15% polyethylene glycol (PEG) 6000, 15% PEG6000 + 250 mM NaCl), drought, 100 µM ABA stresses. The plants were harvested after 0, 30', 1, 5, 10, and 24 h of stress treatment, frozen in liquid nitrogen, and stored at -80°C for further analysis (Rabbani et al. 2003). Total RNA from were extracted using Trizol reagent (Invitrogen, USA) according to the manufacturer’s protocol. Real-time PCR was conducted using a Bio-RAD I Cycler IQ5 machine as previously described using RT pre-mix (TOYOBO Co., Japan). The threshold cycle (Ct) values of the triplicate PCRs were averaged and the relative quantification of the expression levels was performed using the comparative Ct method (Livak et al., 2001). The fold change in total RNA (target gene) relative to the reference gene (actin gene) was determined by the following formula: fold change
= 2−∆∆Ct, where ∆∆Ct = (Ct target gene − Ct actin gene) transgenic plants − (Ct target gene − Ct actin gene) wild-type plants. The following primers were used for the gene specific primers, i.e. OsOPT10: 5′-ACCGAAGAGGAG GTAGACGA-3′ and 5′-GCCACCACATGTAAGGTG AG-3′, Actin2: 5′-CGCCCGTCATCGTGCGGTTC-3′ and 5′-TCCGCGGGGT CCATGGTCTG-3′. RT-PCR analysis of vegetative tissue (leaf, stem, and root) was conducted using 50 ng of total RNA treated DNase I. The amplification program consisted of an initial step at 42°C for 60 min, 94°C for 5 min, 30 cycles (94°C for 1 min, 58°C for 1 min, 72°C for 1 min), and a final step at 72°C for 10 min and the amplified fragments were run on 1.2% agarose gel.
Subcellular localization analysis
The full open reading frame (ORF) of OsOPT10 was amplified using the cDNA clone mentioned above as template. The PCR product was ligated into the pGWB5- GFP vector, resulting in an in-frame fusion protein of GFP
(green fluorescent protein) gene and the OsOPT10 ORF.
The construct (p35S:GFP-OsOPT10) was transformed into onion epidermal cells by particle bombardment using a Biolistic PDS-1000/He gene gun system (BIO-RAD).
After 24 h incubation of transformed onion epidermal cells, GFP signal was detected by a confocal fluorescence microscope.
Transformation to Arabidopsis and rice
OsOPT10 gene isolated from rice (Oryza sativa) var.
Dongjin was introduced into the pB2GW7,0 gateway vector for Agrobacterium-mediated plant transformation (Karimi et al. 2002). Arabidopsis thaliana (ecotype Columbia, Col-O) and rice (Japonica ‘Dongjin’) were transformed with Agrobacterium LBA4404. Stable transformation of Arabidopsis was transformed by the floral dip of simplified method as described by Clough and Bent (1998). Sixty-four independent transformants (T0) were screened on the media containing 3 mg/L phosphinotricine (PPT) and selfed. T1 transformants showing 3:1 segregation ratio were selected, made for T2 homozygous transformants, and analyzed. For the experiments of germination rate of transgenic Arabidopsis homozygote lines (TG-OPT 3), seeds of wild-type and transgenic plants were germinated on ½ Murashige and Skoog (MS) medium (Murashige & Skoog 1962) with or without supplemented NaCl for 2-3 weeks in a culture room maintained at 25°C under a 16 h light/8 h dark cycle. The construct included OsOPT10 gene was trans- formed into Oryza sativa L. cv. Japonica as described by Toki (2006). Transgenic plants in rice were selected in MS medium containing 3 mg/L phosphinotricine (PPT) and analyzed transgenic rice (T2 generation). Genomic DNA was extracted from leaves with the CTAB (hexade- cyltrimethl ammonium bromide) method (Xu et al. 2000).
For 35S::OsOPT10 transgenic analysis, PCR primer set 35S Fw (5′-CGCACAATCCCA CTATCCTT-3′), OsOPT10 Rv (5′-GATGAGCACATG CTCCTTGA-3′) and bar Fw (5′-GTCATCAGATTTC GGTGACG -3′), bar Rv (5′-TT ATTGCGCGTTCAA AAGTC-3′) were designed and synt- hesized. The amplification program consisted of an initial step at 94°C for 2 min, 30 cycles (94°C for 15 s, 53°C 30 s, 72°C for 1 min), and a final step at 72°C 10 min.
Total RNA from 14-day-old seedlings was isolated with
easy-spin plant RNA extraction kit, respectively. RT- PCR analysis was performed as mentioned in the previous chapter.
Salt treatments to transgenic plants
Arabidopsis 35S::OsOPT10 transgenic T2 seeds ger- minated for 7 days in MS medium containing 3 mg/L phosphinotricine (PPT) were transferred to pots filled with a mixture of peat/vermiculite (1:1, v/v) under light at 23°C for 3 weeks. Three-week-old plants were exposed to 150 mM NaCl for 6 days and 6 days for recovery.
Then, the selected plant (TG-T3) was measured for fresh weight compared with wild type (Fig. 6A and B). The four transgenic rice seeds (T4, T7, T9 and T13) were germinated on MS medium containing 6 mg/L PPT for 5 days and grown in MS medium for 3 days with wild type plants. Then at least 60 seedlings for each family were transferred to pots in a green house and grown for 6 days and used in the subsequent salt stress assay.
They were exposed to 250 vmM NaCl for 7 days and allowed to grow on water in a green house for recovery for 7 days (Fig. 8C). For performance of high salt tole- rance of two transgenic plants (T4 and T9) and wild type plants after 250 mM NaCl treatment at 25℃ for 3 weeks and recovery for 2 weeks (Fig. 8B).
Determination of chlorophyll fluorescence
The leaf discs of detached leaves of transgenic OsOPT10 plants (2 weeks old) and WT plants were floated in 1/2 MS liquid medium containing 250 mM NaCl without sucrose (MSH) for 48 h under light at 23°C to examine the health and chlorophyll contents of various discs.
Measurement of chlorophyll fluorescence was performed according to Aono et al. (1993). We repeated experiments on salt stress tolerance three times (Fig. 6C and D).
Results
Mutant selection under salt stress
Altogether, ~15,000 seedlings of T4 seeds from gene- trapped Ds insertion lines, in which Ds had a promoterless
Fig. 1 Appearance of Ac/Ds transgenic plants and wild-type plants after high salt stress in rice. A and B, Growth performance of transgenic in soil containing 250 mM NaCl for 3 days and selection of salt-susceptible mutant rice. C, Fresh weight of mutant line and wild type growing in the 250 mM NaCl containing water for 3 days. The average values were calculated from measures of salt treatment at least 3 times
Fig. 2 Transcript analysis of OsOPT10::Ds. A, Genomic structure of OsOPT10 showing the position of the trap Ds element. Exons are represented by boxes and introns by lines.
The arrows correspond to inserted site. The putative splicing donor (S.D.) and acceptor sites (S.A.) are indicated by the bent arrow. B, Southern hybridization of EcoRI-digested DNA from OsOPT10 plants was performed using probes recognizing a 1.2 kb GUS-encoding region and PCR analysis from OsOPT10 genomic DNA. C, RT-PCR analysis of OsOPT10 mRNA. Total RNA was extracted from the leaves of wild-type and OsOPT10::
Ds plants. D, Nucleotide and deduced amino acid sequence of OsOPT10. The OPT family motifs in OsOPT10 gene are underlined with bold lines
uidA gene encoding β-glucurnidase (GUS) to trap gene (Han, 2002), were screened the one of highly salt-stress sensitive mutants on 250 mM NaCl by using the NaCl responses of Wu et al. (1996). Southern blot analysis showed that the mutant contained single copy of Ds element (data not shown). Flanking sequences of the Ds element revealed that the Ds element was inserted into OPT gene by Blast analysis. Fig. 1 shows the phenotype of the salt sensitive mutants under the conditions of 250 mM NaCl stress. Ds insertion lines on salt-containing solution had significantly lower fresh weight per plant than the wild type (Fig. 1).
Cloning of OsOPT10 gene
To identify the locus in which the Ds element insertion, DNA fragments adjacent to the 5′ and 3′ ends of the inserted Ds were amplified from salt sensitive mutant seedlings using the thermal asymmetric interlaced (TAIL) polymerase chain reaction (PCR) technique (Liu and Whittier 1995). To further characterize the Ds element insertion and fusion transcript from the mutant allele, the junction sequence between the first OsOPT10 exon and β-glucurnidase (GUS)-encoding region was amplified by RT-PCR using mRNA from mutant leaves. Analysis of
flanking sequences revealed that the Ds were inserted into near the end of the third exon of OsOPT10, which gene locus of chromosome 8 (Fig. 2A). The linkage between the phenotype of salt sensitive mutant plants and the Ds insertion was confirmed by DNA gel blot and PCR analysis using genomic DNA isolated from wild- type control, Ds-inserted normal plants, Ds-inserted salt sensitive mutant plants (Fig. 2B). OsOPT10 expression in salt sensitive mutant plants was examined by RT-PCR analysis. Total RNA was extracted from the leaves of wild-type and OsOPT10::Ds plants. RT-PCR analysis indicated that the OsOPT10 gene was not expressed in mutant plants. Therefore these results indicate that disruption of the OsOPT10 gene causes a salt sensitive phenotype (Fig. 2C). Based on the sequence of flanking
Fig. 3 Sequence alignment of NPG and KIPPR motifs from different proteins. A, Multiple sequence alignment of the known members of the OPT family. Accession numbers are as follows: AtOPT1, AB026659 GI:9758213; AtOPT2, AAB60748 GI:2160185; AtOPT3, Z97341 GI:2244994; AtOPT4, AB008268 GI:9759417; AtOPT5, AL078465 GI:4938497; AtOPT6, AL035602.1 GI:4469024; AtOPT7, AF080119 GI:3600039; AtOPT8, BAB09728.1 GI:9759191; AtOPT9, AB015476 GI:9759190; OsOPT8, AP005859; OsOPT9, AK100814;
Opt1p, Z49487; CaOpt1p, U60973 and Isp4p, P40900. B, The residues in motif forming the OPT family protein sites. C, Phylogenetic relationship between OsOPT10 protein and other plant OPT-domain proteins
region, we obtained the cDNA of OsOPT10 contains a 2259-bp open reading frame encoding a protein of 752 amino acids with a calculated molecular mass of 82.8 kDa, tentatively designated as OsOPT10 (GenBank accession no. NP_001062118.) (Fig. 2D).
Molecular characterization of OsOPT10 protein
An alignment of the predicted amino acid sequence of OsOPT10 with the complete sequence of the C. albicans OPT, CaOpt1p (Lubkowitz et al. 1997) was conducted using the DNASTAR program. This analysis led to the identification of nine possible Arabidopsis OPT orthologs and two possible O. sativa OPT paralogs (Fig. 3A and C) that exhibited 49% to 53% sequence similarity to CaOpt1p. The OPTs of Arabidopsis and rice formed a distinct subgroup when compared with the yeast OPT members and showed 61% to 85% sequence similarity when compared with each other (Fig. 3A). The size of proteins predicted for each of the AtOPTs were similar
in size to CaOpt1p (783 amino acids) and Opt1p (799 amino acids) according to Koh et al. 2002. Interestingly, sequence comparisons revealed two domains that were strongly conserved among all of the nine OPT family members in Arabidopsis and the two OPT family members in rice (Vasconcelos et al. 2008). Both of these motifs (i.e. NPG motif and KIPPR motif) were found in regions of the protein predicted to be hydrophilic (Fig. 3B). As reported previously by Lubkowitz et al. (1998), none of the OPT family members showed any significant sequence similarity to the known ABC or PTR peptide transporters (data not shown).
Expression of OsOPT10 gene with abiotic stresses
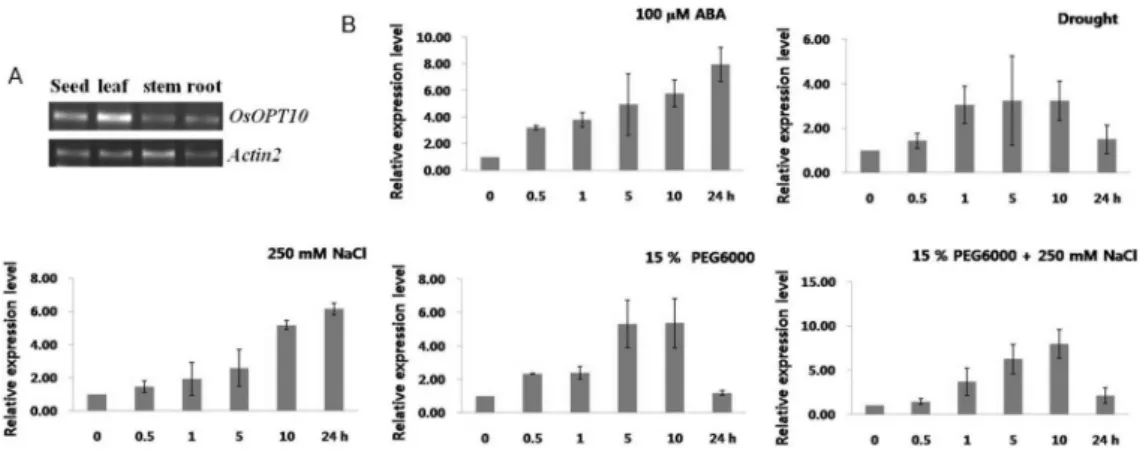
The results from RT-PCR analysis showed that the OsOPT10 gene was detected in various organs including vegetative tissue (leaf, stem, and root), respectively, of rice of 2 weeks old. The gene expression was much higher in one of the leaves than other tissues (Fig. 4A).
Fig. 5 Plasma membrane localization of OsOPT10 gene.
p35S:GFP-OsOPT10 was transiently expressed in onion epidermal cells. The photograph was taken in the dark field for green fluorescence (a), under bright light for the morphology of the cell (b)
Fig. 4 A, Expression profiles of OsOPT10 in organs collected from rice. The actin2 gene was expressed as a positive control and was detected in all organs B, Real-time PCR analysis of OsOPT10 gene in response to 250 mM NaCl, 15% PEG600, osmotic (PEG+
NaCl), drought and 100 μM ABA for the indicated amount of time in leaves of 14 days old seedlings, respectively. Data presented are means and standard deviations of 3 independent replicated experiments
Under conditions of salt stress (250 mM NaCl), the expression levels peaked at 24 h and these levels increased to a very high value within 30 min and decreased after 24 h were exposed to 15% PEG6000 and 15% PEG6000 + 250 mM NaCl conditions. In the case of 100 µM ABA treatment, the OsOPT10 gene was induced strongly within 30 min and continued to be expressed until 24 h after treatment. However, under drought stress, induction of OsOPT10 expression was relatively low, thought the levels increased to 30 min until 10 h (Fig. 4B). Therefore, it can be concluded that OsOPT10 gene expression may lead to the tolerance of transgenic plants to salt and osmotic stress.
OsOPT10 gene was localized in the plasma membrane We analyzed the subcellular localization of OsOPT10 by Agrobacterium-mediated transient expression analysis of a OsOPT10–GFP fusion gene in onion epidermal cells.
As shown in Figure 5A, cells transformed with OsOPT10 –GFP displayed fluorescence specifically in the plasma membrane, suggesting that OsOPT10 is a membrane- localized protein.
Increases of salt stress tolerance in transgenic plants To study the physiological functions of OsOPT10, we generated transgenic Arabidopsis and rice plants that overexpressing OsOPT10 by fusing its full-length cDNA sequence to the CaMV 35S promoter in the pB2GW 7.0 gateway vector (data not shown). The positive trans-
genic plants were confirmed by genomic PCR, Southern blot, and RT-PCR (data not shown). We measured the fresh weight of transgenic Arabidopsis plants used for salt tolerance after 6 days of recovery and transgenic lines that showed no apparent signs of chlorosis (Fig. 6). Figs.
6A and B show absolute variation in the experiment such as the stressed transgenic seedlings decreased 0.17 ~0.21 g/plant of fresh weight, whereas WT plants decreased 1.22~1.31 g/plant of fresh weight as compared to unst- ressed seedling. When the leaf discs of WT and transgenic plants were floated in 250 mM NaCl solution for 48 h, the discs of the transgenic leaves were fresher than those of WT leaves, which agreed with the measurement of their chlorophyll content (Fig. 6C, D). Salt stress tolerance revealed that the percentage of germination of WT plants was lower than the transgenic plants over 2-3 weeks period under the treatment of 50 mM and 150 mM NaCl (Fig. 7A, B). Also, the percentage of green seedlings between WT and transgenic plants showed a
Fig. 6 Phenotype of the 35S::OsOPT10 transgenic plants and wild-type plant exposed to high-salt stress in Arabidopsis. A, Growth performance of transgenic in soil containing 150 mM NaCl for 6 days and 6days for recovery and selection of salt-resistance mutant in Arabidopsis. B, Fresh weight of mutant line and wild type growing in the 150 mM NaCl containing water for 6 days. The average values were calculated from measures of salt treatment at least 3 times. C, For high-salt treatment, leaves from 3-week-old T2 transgenic plants and controls were cut and floated in ½ MS medium containing 250 mM NaCl without sucrose (MSH) for 48 h under light at 23℃. B and C, Phenotype differences were observed and photos were taken at 48 h and chlorophyll contents (nmol g-1 fresh weight) were measured at 48 h for leaves in 250 mM NaCl
Fig. 7 Effect of salt stress on Arabidopsis seedlings from wild type (WT) and OsOPT10 transgenic homozygote lines (TG-OPT 3).
A, Seeds were germinated and grown for 2-3 weeks on agar plates containing ½MS medium with or without supplemented NaCl.
B and C, Salt stress tolerance revealed that the percentage of germination and green seedling of WT and transgenic plants. The percentage is shown relative to germination and green seedling of unstressed seedlings. Error bars are based on three replicates
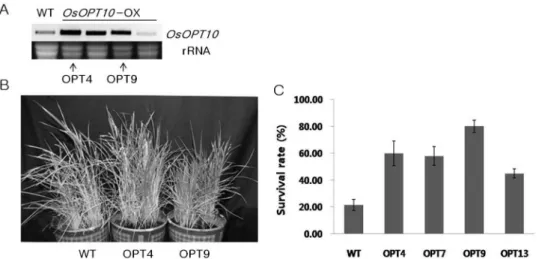
marked difference (Fig. 7C). To analyze whether over- expression of OsOPT10 in rice can improve salt tolerance, we generated transgenic rice plants that were confirmed by transcript analysis (Fig. 8A). Four transgenic plants (T4, T7, T9 and T13) were exposed to 250 mM NaCl for 7 days and allowed to grow on water in a green house for recovery for 7 days. In salt stress treatment, we clearly observed transgenic rice plants grew better than wild type plants as reflected by survival rate com-
parisons (Fig. 8C). Also, selected transgenic plant lines (T4 and T9) were significantly higher than the survival rate of the wild type (Fig. 8B). These results suggested that overexpression of OsOPT10 could enhance the tolerance of transgenic plants to salt stress. Drought tolerance of OsOPT10 transgenic plants were also tested, but no significant effect was detected for this stress (data not shown).
Fig. 8 Identification and salt-tolerance testing of transgenic rice. A, RT-PCR analysis of part of the transgenic plants and the wild type. B, Performance of salt tolerance of two transgenic families and wild type plants after salt treatment (250 mM NaCl for 3 weeks) and recovery (25℃ for 2 weeks). C, Survival rate of four transgenic lines and the wild type after salt treatment (250 mM NaCl for 7 days and 25℃ for 7 days for recovery). Error bars are based on three replicates
Discussion
In the present study, we showed some lines of evidence that support the role of a rice oligopeptide transporter, OsOPT10, as a salt tolerance gene. First, OsOPT10 overe- xpressing plants are tolerance to salt stress (Fig. 6~8), and OsOPT10 Ac/Ds knockout mutants are sensitive to salinity (Fig. 1). Second, OsOPT10 overexpressing plants germinate much better in sodium-containing 1/2 MS agar plates than wild-type plants (Fig. 7). Third, chlorophyll contents and fresh weight of OsOPT10 overexpressing plants are higher than those of wildtype plants (Fig.
6, 8). Fourth, OsOPT10 gene is expressed in a cell membrane (Fig. 5). These data suggest that OsOPT10 contributes to salt tolerance of Arabidopsis and rice in cell membrane by reducing sodium contents, at least in part. Searching of the available Arabidopsis sequences using the CaOpt1p sequence identified nine (AtOPT1 ~
AtOPT9) possible OPT orthologs (Koh et al. 2002).
The presence of several OPT transporters in Arabidopsis suggests that these proteins are important and, so small peptides might play a crucial physiological role in plants.
All OPT family members (plant and fungus) appear to be integral membrane proteins with 12 to 14 predicted transmembrane (TM) domains but the rice OPT proteins appear to have 15 to 18 TM domains (Vasconcelos et al., 2008). Furthermore, the availability of these OsOPT sequences in rice allowed us to identify two highly conserved sequence motifs found in all OPT members
such as Arabidopsis. The OsOPTs (OsOPT1~ OsOPT9) contain the two signature motifs characteristic of the OPTs of PT clade (Koh et al. 2002) (Fig. 3A and B).
The same was true for OsOPT1 and OsOPT2, which showed different expression profiles despite having 81%
sequence identity (Vasconcelos et al. 2008). OsOPT10 gene was predicted to subfamily with OsOPT1 and OsOPT8 (Fig. 3C). It has been shown in Arabidopsis and rice that AtOPT3 (Stacey et al. 2002), AtOPT6 (Cagnac et al. 2004) and OsOPT1 (Vasconcelos et al. 2008) are expressed in developing embryos and in seed maternal tissue. In all plants, including rice, the transport of ions across the cell plasma membrane and across intrace- llular membranes of different organelles must be done with the help of specific ions transporters. It is known that sodium has a toxic effect on many enzymes if present at high concentrations. In addition, high salt conditions in the environment cause low water availability, making it difficult for the plant to acquire water and nutrients.
Therefore, salt stress results in a water-deficient condition in the plant, which resembles a physiological drought.
Conversely, drought decreases water contents in the cell and there by increases the salt concentration (Kim et al.
2010). Therefore, high salt, drought and osmotic stresses, which commonly result in dehydration of the cell and osmotic imbalance (Blumwald et al. 2000 Mahajan et al.
2005), can lead to the same defense mechanism involving OsOPT10 gene. In the present work, we showed that
OsOPT10 confers salt tolerance and relate to osmotic stress (Fig. 4B). Interestingly, some genes induced transcriptionally by osmotic stress also participate in plant defense responses to wounding and pathogen attack.
They include peroxidase, PR-1, PR-10 and osmotin (PR-5) genes (Zhu et al. 2000). Pretreatment of plants with SA, an important regulator of systemic acquired resistance and inducer of many pathogenesis-related (PR) genes, induces antioxidant enzymes and leads to increased salinity, drought or chilling tolerance (Kang and Saltveit 2002, Jacob et al. 2005). However, drought stresses induced very low the expression of OsOPT10 (Fig. 4B). In conclusion, our results show that OsOPT10 is important for osmotic and salt stress tolerance in Arabidopsis and rice, either directly or indirectly by reducing sodium contents in plants. For application, it might be useful to either search for crop varieties with constitutively high expression of the OsOPT10 gene or to develop crops with enhanced tolerance to salt stress.
References
Aono M, Kubo A, Saji H, Tanaka K, Kondo N (1993) Enhanced tolerance to photooxidative stress of transgenic Nicotiana tabacum with high chloroplastic glutathione reductase activity. Plant Cell Physiol 34:129-135
Blumwald E, Aharon GS, Apse MP (2000) Sodium transport in plant cells. Biochim Biophys Acta 1465:140-151 Bray EA (2004) Genes commonly regulated by water-deficit
stress in Arabidopsis thaliana. J Exp Bot 55:2331-2341 Cagnac O, Bourbouloux A, Chakrabarty D, Zhang MY,
Delrot S (2004) AtOPT6 Transports Glutathione Derivatives and Is Induced by Primisulfuron. Plant Physiology 135:
1378-1387
Chin HG, Choe MS, Lee SH, Park SH, Koo JC, Kim NY, Lee JJ, Oh BG, Yi GH, Kim SC, Choi HC, Cho MJ, Han CD (1999) Molecular analysis of rice plants harboring an Ac/Ds transposable element-mediated gene trapping system.
Plant J 19:615-623
Clough S, Bent AF (1998) Floral dip: a simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J 16:735-743
Fei YJ, Kanai Y, Nussberger S, Ganapathy V, Leibach FH, Romero MF, Singh SK, Boron WF, Hediger MA (1994) Expression cloning of a mammalian proton-coupled oligo- peptide transporter. Nature 368:563-566
Frommer WB, Hummel S, Rentsch D (1994) Cloning of an Arabidopsis histidine transporting protein related to nitrate and peptide transporters. FEBS Lett 347:185-189 Ganapathy ME, Brandsch M, Prasad PD, Ganapathy V, Leibach
FH (1995) Differential recognition of β-lactam antibiotics by intestinal and renal peptide transporter, PepT1 and PepT2. J Biol Chem 270:25672-25677
Greenway H, R Munns (1980) Mechanisms of salt tolerance in nonhalophytes. Ann Rev Plant Physiol 31:149-190 Hauser M, Donhardt AM, Barnes D, Naider F, Becker JM
(2000) Enkephalines are transported by a novel eukaryotic peptide uptake system. J Biol Chem 275:3037-3041 Han CD (2002) Rice functional genomics by transposon
mutagenesis. Asia Pacific Biotech News 6:930-935 Jacob G, Ton J, Flors V, Zimmerli L, Metraux JP, Mauch-Mani
B (2005) Enhancing Arabidopsis salt and drought stress tolerance by chemical priming for its abscisic acid responses.
Plant Physiol 139:267-274
Kim CM, Je BI, Piao HL, Park SJ, Kim MJ, Park SH, Park JY, Park SH, Lee EK, Chon NS, Won YJ, Lee GH, Nam MH, Yun DW, Lee MC, Cha YS, Lee KH, Eun MY, Han CD (2002) Reprogramming of the activity of the activator/
dissociation transposon family during plant regeneration in rice. Mol. Cells 14:231-237
Kim CM, Piao HL, Park SJ, Chon NS, Je BI, Sun BY, Park SH, Park JY, Lee EJ, Kim MJ, Chung WS, Lee KH, Lee YS, Lee JJ, Won YJ, Lee GH, Nam MH, Cha YS, Yun DW, Eun MY, Han CD (2004) Rapid, large-scale generation of Ds transposant lines and analysis of the Ds insertion sites in ride. Plant J 39:252-263
Kang HM, Saltveit ME (2002) Chilling tolerance of maize, cucumber and rice seedling leaves and roots are differentially affected by salicylic acid. Physiol Plant 115:571-576 Karimi M, Inze D, Depicker A (2002) Gateway vectors for
Agrobacterium-mediated plant transformation. Trends Plant Sci 7(5):193-195
Koh S, Wiles AM, Sharp JS, Naider FR, Becker JM, Stacey G (2002) An Oligopeptide Transporter Gene Family in Arabidopsis. Plant Physiol 128:21-29
Kim DY, Jin JY, Alejandro S, Martinoia E, Lee Y (2010) Overexpression of AtABCG36 improves drought and salt stress resistance in Arabidopsis. The physiologia Plantarum 139:170-180
Liang R, Fei YJ, Prasad PD, Ramammorthy S, Han H, Yang FRL, Hediger MA, Ganapathy V, Leibach FH (1995) Human intestinal H/peptide co-transporter: Cloning, functional expression, and chromosomal localization. J Biol Chem 270:6456-6463
Liu YG, Whittier RF (1995) Thermal asymmetric interlaced
PCR: automatable amplification and sequencing of insert end fragments from P1 and YAC clonesfor chromosome walking. Genomics 25:674-681
Livak KJ, Schmittgen TD (2001) Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 22-△△CT Method. METHODS 25:402-408
Lubkowitz MA, Hauser L, Breslav M, Naider F, Becker JM (1997) An oligopeptide transport gene from Candida albicans. Microbiology 143:387-396
Lubkowitz MA, Barnes D, Breslav M, Burchfield A, Naider F and Becker JM (1998) Schizosaccharomyces pombe isp4 encodes a transporter representing a novel family ofoligo peptide transporters. Mol Microbiol 28:729-741
Maathuis FJM, Amtmann A (1999) K+ nutrition and Na+
toxicity: the basis of cellular K+/Na+ ratios. Annals of Botany 84:123-133
Maeda T, Wurgler-Murphy SM, Saito H (1994) A two- component system that regulates an osmosensing MAP kinase cascade in yeast. Nature 369:242-245
Mahajan S, Tuteja N (2005) Cold, salinity and drought stresses:
an overview. Arch Biochem Biophys 444:139-158 Murashige T, Skoog F (1962) A revised medium for rapid
growth and bioassays with tobacco tissues. Physiol. Plant 15:473-497
Perry JR, Basrai MA, Steiner HY, Naider F, Becker JM (1994) Isolation and characterization of a Saccharomyces cerevisiae peptide transport gene. Mol Cell Biol 14:104-115 Rabbani MA, Maruyama K, Abe H, Khan MA, Katsura K, Ito
Y, Yoshiwara K, Seki M, Shinozaki K, Yamaguchi- Shinozaki K (2003) Monitoring expression profiles of rice genes under cold, drought, and high-salinity stresses and abscisic acid application using cDNA microarray and RNA gel-blot analyses. Plant Physiol 133:1755-1767
Sanders D, Brownlee C, Harper JF (1999) Communicating with calcium. Plant Cell 11:691-706
Stacey G, Koh S, Granger C, Becker JM (2002a) Peptide transport in plants. Trends in Plant Science 7:257-263 Stacey MG., Koh S, Becker J, Stacey G (2002) AtOPT3, a
member of the oligopeptide transporter family, is essential for embryo development in Arabidopsis. The Plant Cell 14:2799-2811
Stacey MG, Osawa H, Patel A, Gassmann G, Stacey G (2006) Expression analysis of Arabidopsis oligopeptide transporters during seed germination, vegetative growth and reproduction.
Planta 223:291-305
Steiner HY, Song W, Zhang L, Naider F, Becker JM, Stacey G (1994) An Arabidopsis peptide transporter is a member of a new class of membrane transport proteins. Plant Cell 6:1289-1299
Teige M, Scheikl E, Eulgem T, Doczi R, Ichimura K, Shinozaki K, Dangl JL, Hirt H (2004) The MKK2 pathway mediates cold and salt stress signaling in Arabidopsis. Mol Cell 15:141-152
Vasconcelos Ma W, Li GW, Lubkowitz MA, Grusak MA (2008) Characterization of the PT Clade of Oligopeptide Trans- porters in Rice. The plant genome 1:77-88
West CE, Waterworth WM, Stephens SM, Smith CP, Bray CM (1998) Cloning and functional characterization of a peptide transporter expressed in the scutellum of barley grain during the early stages of germination. Plant J 15:
221-230
Williams LE, Miller AJ (2001) Transporters responsible for the uptake and partitioning of nitrogenous solutes. Annu Rev Plant Physiol Plant Mol Biol 52:659-688
Wu SJ, Ding L, Zhu JK (1996) SOS1, a genetic locus essential for salt tolerance and potassium acquisition. Plant Cell 8:617-627
Xiang Y, Huang Y, Xiong L (2007) Characterization of Stress- Responsive CIPK Genes in Rice for Stress Tolerance Improvement. Plant Physiology 144:1416-1428
Xu RQ, Tomooka N , Vaughan DA (2000) AFLP Markers for characterizing the azuki bean complex. Crop Science 40:808-815
Yoshida S, Forno DA, Cock JH, Gomez KA (1976) Laboratory manual for physiological studies of rice (3eds). Philippines:
IRRI 83
Yun CH, Kim KH, Calcutt MW, Guengerich FP (2005) Kinetic analysis of oxidation of coumarins by human cytochrome P450 2A6. J Biol Chem 280:12279-12291
Zhang MY, Bourbouloux A, Cagnac O, Shrikanth CV, Rentsch D, Bachhawat AK, Delrot S (2004) A novel family of transporters mediating the transport of glutathione derivatives in plants. Plant Physiol 134:482-491
Zhu JK (2000) Genetic Analysis of Plant Salt Tolerance Using Arabidopsis. Plant Physiology 124:941-948
Zhu JX, Sasano Y, Takahashi I, Mizoguchi I, Kagayama M (2001) Temporal and spatial gene expression of major bone extracellular matrix molecules during embryonic mandibular osteogenesis in rats. Histochem J 33:25-35