Growth and Fruit Characteristics of Aneuploid Apple Obtained from Crosses between Diploid and Triploid
Chun-Hua Zhang and Sung-Min Park*
Department of Horticulture, Kangwon National University, Chuncheon 200-701, Korea
Abstract.
This study was conducted to observe some characteristics of aneuploid trees obtained from the crosses between diploid and triploid in the first experiment. The results showed that the difference among the percentage of aneuploid’s pollen grain germination was large, and all of them were lower than that of diploid ‘Hongro’ (82.4%). The average weight and size of each aneuploid's fruit was far lighter and smaller than that of diploid ‘Hongro’. According to the width and length, all of the fruits shape was flat or short-glo- bose conical shape except for JF3942. Almost all the aneuploid had higher sugar content than ‘Hongro’, nearly half of aneuploid fruits were firmer than that of diploid ‘Hongro’. As for the acid contents of aneup- loid fruits, the results were greatly different from those of previous studies on diploid apples, in present study the citric acid and tartaric acid contents were more than the malic acid which are dominant acid in most cultivars of apples. Both the length and width of aneuploid tree were shorter than that of diploid ‘Hon- gro’. Most aneuploid trees’ height was longer than width, but there were 4 exceptions. The size of leaves was smaller than that of diploid ‘Hongro’ according to the length and width of leaves measured. In all, ane- uploid’s vegetative growth is not as vigorous as diploid.
Key words :
apple, aneuploid, germination, growth and fruit characteristics
Introduction
Aneuploidy refers to the nuclear condition when the chromosome count is not exact multiple of the base num- ber of the species. The term was introduced by Tackholm (1922) following his studies on genus
Rosa. Realization of the possible utility of aneuploids generated a spurt of activity, and as a consequence much literature has accu- mulated on the subject of aneuploidy in various plants.
But little research was carried out on the apple aneup- loidy. To our knowledge at hand, aneuploid apple was first mentioned in the study results by Crane and Lawrence (1930) and Darlington and Moffett (1930).
Along with other researchers they found when triploid apples were crossed with diploids or open-crossed, most seedlings obtained were aneuploid with much poorer tree growth in the early growth stage (Crane and Lawrence, 1929, 1930, 1933; Darlington and Moffett, 1930; Mof- fett, 1931). Darlington and Moffett (1930) along with Crane and Lanwrence (1930) even concluded that from
the work already done, it seems safe to predict that ane- uploid forms rarely appear in the cultivated apple. So at that time no further information about aneuploid apple was offered. But Japanese researchers, Sato et al., showed a lot of interest on the aneuploid apple from 1989 until now. In his study, although they suggested most seedlings obtained from crosses were aneuploidy when one of the parents was triploid, especially when triploid was as female parent, they still further observed the efficiency of well-grown seedlings, vegetative growth, flowering and fruiting of seedling obtained from the crosses between diploid and triploid (Sato and Kanbe, 1989, 2007; Sato, 1990; Sato et al., 2007). Their results indicated the characteristic difference of whole proge- nies obtained from each cross, furthermore, they did not make sure those obtained from each cross were all ane- uploid or not, or how much percentage of aneuploid among of them. The seedlings obtained must contain some diploid seedlings. So as for describing the aneup- loidy seedlings characteristics, those data are not suffi- cient and exact enough.
Until now no paper purposely describes the character- istics of aneuploid apple tree in detail. Most researchers
*Corresponding author: [email protected]
*Received November 17, 2009; Revised November 30, 2009;
*Accepted December 15, 2009
neglected them. In fact, in our opinion, aneuploidy apple has a lot of potential worthy of exploiting.
In the last paper (Zhang and Park, 2009), 181 aneup- loid apple trees were obtained through crosses between diploid and triploid. As a result of meiotic irregularity, in this study we hypothesized that unreduced gametes and gametes with aberrant chromosome or some reduplicate segment may lead to special characteristics of their prog- enies. 12 aneuploid trees which began to flower after 6 year sowing were selected to observe their growth char- acteristics including vegetative growth and fruit in this study. The purpose of this is to find theirs special poten- tial which is worthy of using as breeding materials.
Materials and Methods
1. Pollen germination of aneuploids progenies obtained from the crosses between diploid and triploid The materials were the 12 aneuploids obtained from the progenies of crosses between diploid and triploid in last paper and a cultivar ‘Hongro’ (2x) as control. All of the trees were in the farm of Kangwon National Univer- sity, Chuncheon, Korea. The flowers were picked from each aneuploid tree before the cluster dehisced pollen in April 28th, 2008. The anthers were then put in the petri dish under fluorescent lamp over night at about 25
oC until the anthers chapped. Then pollen were scattered gently on the medium which contained 0.001 g/L Boric acid, 20 g/L Sucrose and 8 g/L agar at 22
oC for 4 h under light. The grains from each replication in five micro- scopic areas were counted randomly. All observations of slides were carried out at 200 × magnification using a light microscope.
2. Vegetative growth of aneuploid progenies obtained from the crosses between diploid and triploid
The aneuploid seedlings were not pruned until the third growing season after sowing. After that, pruning was carried out each winter to achieve a central leader tree shape.
The growth was measured in October, 2008. As for length of the one-year-old new branch, five branches were measured for each aneuploid tree. The length and width of the fifth leaf of each branch were measured, rep- licated 5 times per aneuploid tree. The length and width
of whole tree were measured using a tapeline and repli- cated 5 times.
3. Fruit characteristics of aneuploid progenies obtained from the crosses between diploid and triploid The fruits from each aneuploid tree were weighed one by one and then averaged value. The length and width of each fruit from each aneuploid tree were measured using vernier caliper and averaged the value. Total sugar con- tent (degrees
oBrix) was measured by refractometer (Atago PR1, Atago, Tokyo, Japan).
The firmness of the fruit was measured at two posi- tions, red blushed and unblushed background surface areas, at the equator of each fruit after removing the skin.
The firmness was measured by the instrument of Com- pac-100 II (Sun Scientific Co., Tokyo, Japan) with an 11 mm probe. The probe ran at a speed of 2 mm/s and penetrated the peeled portion of the fruit 9 mm/s. 5 fruits of each aneuploid tree were measured. Maximum forces determined kg/cm
2was used as a measure of fruit firm- ness.
Then the flesh was cut to small pieces and was imme- diately frozen in liquid nitrogen, put in plastic bag and flushed with nitrogen before storage at
−80
oC for the analysis the acid content. Citric acid, malic acid, and tar- taric acid were determined by titrating the juice extracted from 5 ± 0.1 g frozen flesh sample and 20 ml distilled water using a digital blender (Ultra-Turrax, IKA, T25) to pH 8.0 using 0.1 M NaOH in a Mettler DL15 titrator (Mettler-Toledo, DL15, Switzerland). The titrated vol- ume (milliliters) expressed directly in grams/litre of citric acid, malic acid, and tartaric acid. All of them repeated 5 times, each replicate was prepared separately.
The results were statistically evaluated by SPSS pro- gram version 10.01 (SPSS Inc., Chicago, IL).
Results
1. Germination rate of pollen grain in the aneuploid progenies obtained from the crosses between diploid and triploid
After being sown on the media, the pollen grains
began to germinate within two or three hours. Pollen
tubes developed from the pollen grains. Pollen tubes first
emerged through a germ furrow and grew in length. 12
aneuploid trees’ germination rate ranged from 6.6 (JN4566) to 73.5% (JS6), pollen germination rate of most aneuploid trees was less than 50%. The percentage of pollen germination of most aneuploids was low, as compared with that of diploid (82.4%), only JS6 was
near to diploid (Table 1). Although all of them are ane- uploids, the percentage differences of pollen germination were large. No significant correlation was found between aneuploid chromosome number and pollen germination.
Aneuploid apple pollen grains were triangular with four
Table 1. Pollen germination rate of aneuploid progenies.
Aneuploid trees Ploidy level No. of pollen observed No. of germinated pollen Germination rate (%)
SJ277. 2n = 2x + 1 = 35 044.3 29.6 66.7 cz
JT4. 2n = 2x + 1 = 35 067.3 16.0 23.8 f
JN811. 2n = 2x + 2 = 36 077.9 11.5 14.7 h
JF1049. 2n = 2x + 2 = 36 070.3 26.3 37.4 d
JF10. 2n = 2x + 2 = 36 039.6 13.0 32.9 e
JS6. 2n = 2x + 2 = 36 042.6 31.3 73.5 b
JF3942. 2n = 2x + 3 = 37 047.5 07.7 16.2 gh
JN6645. 2n = 2x + 3 = 37 080.0 05.3 06.6 i
JN4831. 2n = 2x + 6 = 40 066.9 11.1 16.6 gh
JF1650. 2n = 2x + 11 = 45 052.6 18.9 36.1 de
JF3252. 2n = 2x + 11 = 45 102.0 19.6 19.2 g
JT1035. 2n = 2x + 12 = 46 098.4 13.1 13.3 h
Hr 2n = 2x = 34 088.8 73.2 82.4 a
Note: F, ‘Fuji’; J, ‘Jonagold’; N, ‘NJ.53’; S, ‘Senshu’; T, ‘Tsugaru’; Hr, ‘Hongro’.
Z: Mean separation within columns by Duncan’s multiple range test at 5% level.
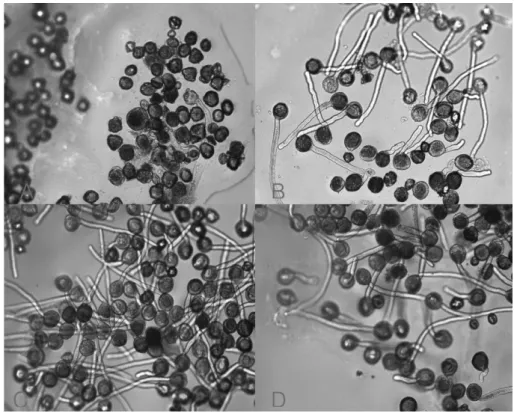
Fig. 1. Figures of some representative pollen germination in the aneuploid progenies. A: JN6645; B: SJ277; C: ‘Hongro’; D:
JS6.
germspores. The pollen tube came out through only one pore and attains roughly a length of three to eight times the diameter of the pollen grain. Some pollen tubes remain short and bulge at the tips. Most of the pictures contain a mixture of normal and germinating pollen grains in varying proportions (Fig. 1).
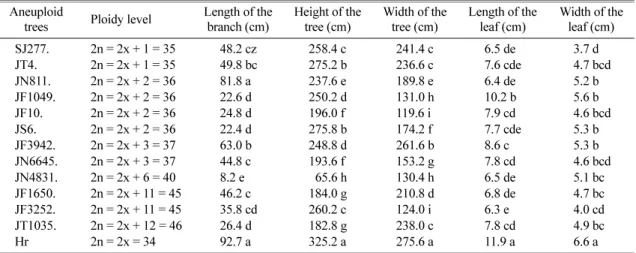
2. Vegetative growth of the aneuploid progenies obtained from the crosses between diploid and triploid The average length of the shortest and longest branch was 8.2 cm and 81.8 cm for JN4831 and JN811, respec- tively (Table 2). Only the branch length of JN811 had no difference from ‘Hongro’, the others of that had signifi- cant difference from the control. The length of leaf ranged from 6.3 to 10.2 cm and the width of leaf ranged from 3.7 to 5.6 cm in all the 12 aneuploid trees. The leaf size of ‘Hongro’ (control) was significantly different from all the aneuploid trees. The height of 6-year-old aneup- loid ranged from 65.6 to 275.8 cm, shorter than that of diploid, 325.2 cm. As for width of aneuploid trees, it ranged from 119.6 to 261.6 cm, significantly shorter than diploid of that. On the whole most aneuploid tree’s height was longer than width, but there were some exceptions.
Aneuploid JN3942, JN4831, JF1650, and JT1035, whose length/width of tree were 248.8/261.6, 65.6/130.4,184.0/
210.8, and 182.8/238.0, respectively. JN4831 was the lowest tree among all of the aneuploids, and the width was 2 times of its height (Table 2). Furthermore, the leaf
size and one-year-old branch length of aneuploid did not increase as the tree size increased. In the whole, all of aneuploid were not as vigorous as diploid apple trees.
3. Fruit characteristics of aneuploid progenies obtained from the crosses between diploid and triploid The average fruit weight of each aneuploid tree ranged from 25.4 g (JT1035) to 112.9 g (JF3252) (Table 3) and had significant difference from the ‘Hongro’. The weight per fruit of ‘Hongro’ was more than 2 times of that of aneuploid trees. The length and width of fruit indicated the shape of the fruit through the Length/Width ratio which was not listed in the table. Except for the Length/
Width of aneuploid JF3942, the others were less than 1.
This indicates all of the fruit shape was flat or short-glo- bose conical shape except for JF3942. The Length/Width of JF3942 indicated the shape of fruit was flat-globose or intermediate conical, the height of fruit was longer than width of it. The measurement of length and width of each fruit not only can judge the shape of fruit, but also can see the size of each fruit. Fruit size is important factor influencing apple orchard profitability (Miranda et al., 2007). JT1035 had the smallest size of fruit, JT4 had the biggest size fruit among the aneuploid relatively, but the size of all the aneuploid fruits were smaller than that of
‘Hongro’. The size of the fruit had close relationship with the weight of fruit.
It has been verified that the mechanical properties of
Table 2. The vegetative growth of aneuploid progenies.
Aneuploid
trees Ploidy level Length of the
branch (cm) Height of the
tree (cm) Width of the
tree (cm) Length of the
leaf (cm) Width of the leaf (cm)
SJ277. 2n = 2x + 1 = 35 48.2 cz 258.4 c 241.4 c 6.5 de 3.7 d
JT4. 2n = 2x + 1 = 35 49.8 bc 275.2 b 236.6 c 7.6 cde 4.7 bcd
JN811. 2n = 2x + 2 = 36 81.8 a 237.6 e 189.8 e 6.4 de 5.2 b
JF1049. 2n = 2x + 2 = 36 22.6 d 250.2 d 131.0 h 10.2 b 5.6 b
JF10. 2n = 2x + 2 = 36 24.8 d 196.0 f 119.6 i 7.9 cd 4.6 bcd
JS6. 2n = 2x + 2 = 36 22.4 d 275.8 b 174.2 f 7.7 cde 5.3 b
JF3942. 2n = 2x + 3 = 37 63.0 b 248.8 d 261.6 b 8.6 c 5.3 b
JN6645. 2n = 2x + 3 = 37 44.8 c 193.6 f 153.2 g 7.8 cd 4.6 bcd
JN4831. 2n = 2x + 6 = 40 8.2 e 065.6 h 130.4 h 6.5 de 5.1 bc
JF1650. 2n = 2x + 11 = 45 46.2 c 184.0 g 210.8 d 6.8 de 4.7 bc
JF3252. 2n = 2x + 11 = 45 35.8 cd 260.2 c 124.0 i 6.3 e 4.0 cd
JT1035. 2n = 2x + 12 = 46 26.4 d 182.8 g 238.0 c 7.8 cd 4.9 bc
Hr 2n = 2x = 34 92.7 a 325.2 a 275.6 a 11.9 a 6.6 a
Note: F, ‘Fuji’; J, ‘Jonagold’; N, ‘NJ.53’; S, ‘Senshu’; T, ‘Tsugaru’; Hr, ‘Hongro’.
Z: Mean separation within columns by Duncan’s multiple range test at 5% level.
fruits, including firmness, are good indicators of their maturity, degree of firmness and predicted shelf life. The data obtained are expressed in terms of the force required to rupture cortex parenchyma cells, and thus represent a compound of many cellular and macrocellular properties including cell turgor and wall strength (King et al., 2000).
The firmness of aneuploid fruits for JF1049, JN6645, JF1650 had no difference, 10.8, 10.5, and 10.3 kg/cm
2, respectively. They have the firmest flesh among all the fruits measured including control ‘Hongro’. The fruits of JT1035 (7.1 kg/cm
2) was the softest among all aneuploid fruits. The firmness of half aneuploid apples was higher than that of diploid ‘Hongro’. Half of them lower than diploid ‘Hongro’. Wide differences in firmness occurred among fruits within aneuploid trees and within individual fruits in each tree.
The sugars and organic acids have been related to fla- vor quality for a variety of fruits and specifically to apple flavor and determine the optimum time for harvesting, because it is considered as an index of quality. The con- centrations of mean total sugars among the aneuploid fruits were found statistically significant. The total sugar concentration was varied between 11.2 and 19.1
oBrix and the values were obtained from JT1035. and JF1049, respectively. Almost all of the aneuploid fruits had higher total sugar content than the ‘Hongro’ (Table 3).
The malic acid concentration ranged from1.2 to 4.9 g/
L. The concentration of malic acid for SJ277 and JN6645 were the lowest and had no significant difference. Except for SJ277 and JF3252, and JN6645 all the other aneup- loid had higher content of malic acid than the ‘Honro’, 2.7 g/L. The malic acid concentration of JT1035, JS6, and JN811 were the highest, no significant difference among them was found (Table 3).
The highest concentration of citric acids in aneuploid fruits were on the JN811 (6.3 g/L), JS6 (6.5 g/L), JT1035 (6.6 g/L), no differences were found among them, but they had significant difference from the control ‘Hon- gro’ (4.5 g/L). The tartaric acid concentration was varied between 3.2 to 8.4 g/L, significantly different from the
‘Hongro’, 4.2 g/L.
Discussion
Fruit development in the apples is dependent in large degree upon successful pollen development and subse- quent fertilization. The quality of pollen available for fer- tilization may affect not only the fruitfulness of existing varieties, but also may influence the germinal constitu- tion of the progeny (Shoemaker, 1926). Different nutri- tion conditions and germination methods for many varieties were used by researchers to know about the pol- len germinability of apple (Calzoni et al., 1979; Adams, 1916). For example, all the triploids germinated were less
Table 3. Fruit characteristics of aneuploid progenies.
Aneuploid
trees Ploidy level Weight of
fruit (g) Width of
fruit (cm) Length of
fruit (cm) Firmness (kg/cm2)
Total sugar (oBrix)
Citric (g/L)acid
Malic
(g/L)acid Tartaric acid (g/L) SJ277. 2n = 2x + 1 = 35 112.4 bz 5.9 cd 5.3 c 07.7 gh 14.2 e 2.8 f 1.7 f 3.7 ghi JT4. 2n = 2x + 1 = 35 107.2 bc 6.1 b 6.0 ab 09.2 de 16.8 bcd 3.8 e 2.8 e 4.6 ef JN811. 2n = 2x + 2 = 36 081.2 cd 4.9 ef 4.4 d 08.7 def 16.0 cde 6.3 a 4.6 ab 8.4 a JF1049. 2n = 2x + 2 = 36 068.8 d 5.0 ef 4.0 d 10.8 a 19.1 a 4.7 cd 3.2 de 5.2 e JF10. 2n = 2x + 2 = 36 064.8 d 5.4 de 3.9 d 09.6 cd 18.0 abc 5.0 bc 4.1 c 6.5 d JS6. 2n = 2x + 2 = 36 087.1 cd 5.8 cd 4.2 d 07.4 gh 11.7 fg 6.5 a 4.5 abc 7.4 b JF3942. 2n = 2x + 3 = 37 079.2 cd 5.4 de 5.7 bc 10.0 b 18.2 abc 5.3 b 4.2 bc 6.6 cd JN6645. 2n = 2x + 3 = 37 103.2 bc 6.2 bc 5.1 c 10.5 ab 17.6 abc 1.4 g 1.2 g 3.4 hi JN4831. 2n = 2x + 6 = 40 070.5 d 5.3 de 4.1 d 08.1 fg 13.8 ef 4.1 e 3.1 de 4.9 e JF1650. 2n = 2x + 11 = 45 085.2 cd 5.8 cd 4.3 d 10.3 ab 18.8 ab 1.7 g 3.5 d 3.2 i JF3252. 2n = 2x + 11 = 45 112.9 b 6.3 b 5.2 c 08.3 fg 15.0 de 3.0 f 2.1 f 3.9 gh JT1035. 2n = 2x + 12 = 46 025.4 e 3.3 g 2.8 e 07.1 h 11.2 g 6.6 a 4.9 a 7.1 bc
Hr 2n = 2x = 34 294.3 a 7.4a 6.8 a 08.7 def 13.8 ef 4.5 d 2.7 e 4.2 fg
Note: F, ‘Fuji’; J, ‘Jonagold’; N, ‘NJ.53’; S, ‘Senshu’; T, ‘Tsugaru’; Hr, ‘Hongro’.
Z: Mean separation within columns by Duncan’s multiple range test at 5% level.
than 30% (Hall, 1931). Larsen and Tung (1950) reported the apple germination rate of James Grieve (2x) and Belle de Boskoop (3x) ranged from 13 to 76% and 0 to 6% under different conditions, respectively; the germi- nation rate of ‘Golden Delicious’ and ‘Starkrimson’
ranged from about 6~91% or 30~90% under different conditions (Calzoni et al., 1979). As for the aneuploid observed in this paper, the germination rate was similar to triploid and diploid on the whole but little lower than the maximum rate of diploid. This is consistent with reports earlier: Beaumont and Knight (1922) described the average percentage of germination denoted differ- ences not only between individual varieties, but also between the members of the Winesap group and Deli- cious. And the germination of pollen themselves is not always the same on media of different origins (Jaroslav, 1962). Generally speaking, some aneuploid apples, like SJ277 and JS6 (Table 1) which have the similar germi- nation rate with diploid, have possibility and potential to be used as male parent of cross in the breeding.
Vegetative growth is an important indicator for crop management in fruit tree cropping systems. The vigor of tree has close relationship with the first flowering,
namely, the vigor of tree affect the juvenility in apple.
Many observations agreed with this phenomenon.
Jonkers (1971) reported the tallest plants flowered first.
Visser (1965) also thinks juvenile period can be short- ened by providing optimal conditions for growth. In this study only some trees finished juvenile within 6 years.
Jonkers (1971) mentioned the average height of 3- year-old James Grieve (2x) was 311 cm, 238 cm and 244 cm, respectively. However, in this experiment, the height of 6-year-old aneuploid ranged from 65.6 to 275.2 cm. This indicates the height of aneuploid was really lower than that of diploid reported before and the control ‘Hongro’. The leaf size was smaller than diploid, there seems no relationship between leaf size or branch length with tree size. In all, the aneuploid tree’s vegeta- tive growth has some different traits from diploid trees (Fig. 2). Aneuploid had more short and blend branches which are easy to grow toward the two sides of the tree not toward upright as in diploid. The growing speed of the whole tree is slow, although manage them carefully and well. This result confirmed the report by Ozaki et al.
(2004): aneuploid, including trisomics, are often morpho- logically distinct because of the addition or lack of chro-
Fig. 2. Figures of some representative aneuploid progenies with fruits. A: JF10; B: JN811; C: JN6645; D: JF1650; E: JT4; F:
JN4831.
mosomes. Some researchers suggested similarly that there was a correlation between weight of seeds and the vigour of seedlings. Abnormalities occur which cause low seed content, poor gerimination of seed and lack of vigour in the seedlings. Lack of vigor of the trees inev- itablely result from the irregularities in germ-cell forma- tion in the parents, and the aneuploid chromosome constitution of their offspring (Crane and Lawrenece, 1933). So conclusion was made that in this study the veg- etative growth of 12 aneuploid trees were not vigorous probably because the weight of seeds which developed to these aneuploid trees was light. Their weight distribution of the seeds mostly ranged from 20 to 35 mg was showed in the last paper (Zhang and Park, 2009). Of course, the underlying reason is the addition or lack of chromosomes of these aneuploid themselves.
Although aneuploid apple trees are not as vigorous as diploid apple trees according to the data obtained, it has a lot of value as a bridge for breeding. For example, this dwarf trait maybe contribute aneuploidy to be as root- stock. Dwarfing rootstocks produce smaller trees that facilitate harvesting, pruning and high-density planting.
Current production methods have shifted towards high- density plantings on dwarfing or very dwarfing root- stocks to achieve greater yields per unit of area (Autio and Southwick, 1986). Other desirable characteristics conferred by the rootstock on the scion include fruiting at an earlier age, improved fruit quality and so on (Wer- theim, 1998).
Some aneuploid fruits in present study were a little firmer than that of the ‘Hongro’ and the previous study in
‘Red Elstar’ by Awad et al. (2001) which ranged from 6.4 to 6.9 kg/cm
2. Some aneuploid fruits were softer than
‘Hongro’ and the reports before about ‘Granny Smith’,
‘Braeburn’, and ‘Fuji’ ranged from 7.8 to 10.1 kg/force (Wand et al., 2006). The fruits of aneuuploid in the present study seem have the similar firmness with these diploid, this maybe due to their small fruit size. Just because the fruit was small, it may has smaller size cells, so the density of cells within the aneuploid fruit is denser than that the big fruit which has big cells with big cyto- plasm and can not stand for heavy pressure. That has relationship with rupturing cortex parenchyma cell, and thus represent a compound of many cellular and macro- cellular properties including cell turgor and wall strength.
Furthermore, as concluded before, the fruit firmness is determined multifactorially by positive and negative effects caused for instance by fruit size, number and size of cells, volume of intercellular space, specific gravity, harvest maturity, dry matter (pectin) and mineral content, enzyme activity, and so on (Link, 2000). So existing cul- tivars vary considerably in their textural properties, and in their response to various environmental factors. How- ever, this is also possibly dependent on the gene they car- ried or inherited from their parents (King et al., 2000).
Each parent possesses different QTL alleles for pene- trometer readings, is able to contribute to increase fruit firmness.
In test conducted by marketing inspectors at harvest, and in breeding selection programs, texture has often been equated with quality in apples; firm, crisp apples being the ideal (King et al., 2000). So some of aneuploid trees has possibility as the material of breeding referring to the firmness.
In the development of new apple varieties, it is impor- tant to evaluate the phenotypic characteristic of apples.
The novel size per aneuploid fruit was significantly
smaller than that of diploid trees, and their weight due to
small size was far lighter than the diploid fruit both in
this study and the studies earlier. Some reports, such as
Fuji (2x), the average weight ranged from 144.0 to
170.4 g, Granny Smith (2x) ranged from 154.4 to 181.9 g
(Wand et al., 2006). Almost all the aneuploid fruits were
flat oblate shapes in this study. Climate (Shaw, 1914),
moisture and mild (McKenzie, 1971), altitude (Eccher,
1986), light (Noe and Tommaso, 1996) and so on, affect
the length/diameter (L/D) and size of apple fruit. But all
of the experimental trees were in the same farm, the big
size of ‘Hongro’ was obtained, this indicted the condi-
tions of the climate and altitude seems have little effect
on the shape and size of aneuploid fruits. The results
obtained by Brown (1960) showed that the fruit shape
and size was a quantitative pattern of inheritance in the
apple progenies obtained from the crosses between dip-
loids. The progeny mean for fruit shape was found to be
approximately equal to the mid-parent value. The prog-
eny mean for fruit size is generally less than the mid par-
ent value, the average decreases from the mid parent
were about 34%. The results obtained on aneuploid agree
with this conclusion partially, furthermore, probably
abnormal chromosome number has additive or major effect on the very small size of aneuploid fruit. The small size of aneuploid fruit has special good trait for breeding, too, because it is easy to hold, less damage during trans- port, the look is delicate and cute, just like small tomato, it may be loved by children and young consumers.
The flavour of apple is greatly influenced by the pres- ence of sugar and acids. The malic acid content in this study were similar to the results reported before (Campo et al., 2006a, b; Jeuring et al., 1979; Whang, 2000; Kenis et al., 2008; Drake and Eisele, 1999), although they used different determined methods. However, the content of citric and tartaric acid in this study was very different from that of results studied before (Campo et al., 2006a, b; Drake and Eisele, 1999), approximately 10 times higher than that measured by Campo. Not only citric and tartaric acids content of aneuploid fruits were signifi- cantly higher than most reports before, but also the ratio of marlic/citric/tartaric was worthy of paying attention.
The sequence of average content of acids was tartaric (5.4 g/L) > citric (4.3 g/L) > malic (3.3 g/L) for the ane- uploid fruits, nearly 1 : 1 : 1. This result totally contrary to the most results reported earlier such as on ‘Fuji’ (2x),
‘Jonagold’ (3x), ‘Delicious’ ‘Gezamina’ and so on (Wu et al., 2007; Campo et al., 2006a, b; Drake and Eisele, 1999) that predominant organic acid in the apples was malic acid, malic acid accounts for about 90% of the acid content of apples; citric, succinic, and traces of several other acids make up the rest (Hulme and Rhodes, 1971), in those studies the ratio of malic, citric and tartaric acids were about 100 : 10 : 1. It is admitted that some degrees of content difference may exist for the observed data before, because the organic acid in fruits may be influ- enced by different types of fruit, even species and vari- ety within species, by the cultivation conditions such as environmental and cultivation techniques, growing sea- son, mesocarp growth curve, respiration (Chun et al., 2005; Lobit et al., 2003; Sturm et al., 2003), determined methods and so on, but the problem in this study is the malic acid was similar to the reports before, only the cit- ric acid, tartaric acid , and the proportion of three acids were much higher than them. This indicated that novel acids content maybe has relationship with the special chromosome number and its gene. Variability contributed by the genotype in the trait ‘fruit acidity’ was explained
by two QTLs on linkages 8 and 16. Namely, MdPEPC, MdVHA-A and MdcyME are putative apple homologues underlying enzyme acitivity of PEPC, VHA-A and cyME, respectively, and MA accumulation in developing and ripening fruits. The homeostasis of their expression, along with the corresponding enzyme activity, leads to the difference in fruit acidity between low and high acid genotypes. The data presented here only described three kinds of acids content during the first one year growth, further understanding of the kinds and content of the organic acid in next few years would be needed. In par- ticular, the proportion of various acids remains to be studied and quantified.
The sugar of aneuploid fruits was similar to that of dip- loids and triploids measured in 7 cultivars (Drogoudi et al., 2008) and in 9
Malus domesticaBorkh cultivars (Petk- ovsek et al., 2007), but higher than that of diploid and trip- loid measured in 3 cultivars (Link, 2000), Aroma cultivar (Hagen et al., 2007), and progenies (Kenis et al., 2008), even the control ‘Hongro’ of this study. This confirmed that intensity of sweetness and acidity of apples are, strongly dependent on both cultivar and climate (Rodbotten et al., 2009), the difference of temperature of day and night.
Although the size of aneuploid fruit was very smaller than the diploid or triploid reported earlier, the content of sugar and acids were not lower than them. The high sugar is benefit to balance the acid to reach suitable sweetness.
The higher organic acids are more responsible for the fresh, tart taste and color of fruit. High levels of these organic acids are known to contribute to the acidity of fruit juices, so this trait is probably benefit for breeding new cultivar for making apple juice or apple cider and so on.
This experiment was just a basic step to find some valuable and special traits of aneuploid apple tree prima- rily which are useful in breeding. Although in this exper- iment we did not measure the parameters of quality in detail, the data obtained primarily showed some aneup- loids has potential for breeding. Next year we will mea- sure the quality of fruit including peel colour, vitamin content, aromaticity and so on to provide more parame- ters for the characteristics of aneuploid apple trees.
Acknowledgements
This study was supported by grant from Bioherb
Research Institute, Kangwon National University, Chun- cheon, Korea.
Literature cited
1. Adams, J. 1916. On the germination of the pollen grains of apple and other fruit trees. Botanical Gazette.
61(2):131-147.
2. Autio, R.W. and F.W. Southwick.1986.The effects of rootstock and root-interstem combination on the growth, productivity, and anchorage of a spur and standard strain Delicious apple tree. Fruit Var. J.
40:128-133.
3. Awad, M.A., A.D. Jager, M. Dekker, and W.M.F. Jon- gen. 2001. Formation of flavonoids and chlorogenic acid in apples as affected by crop load. Scientia Horti- culturae. 91:227-237.
4. Beaumont, J.H. and L.J. Knight. 1922. Apple pollen germination studies. Proc. Amer. Soc. Hort. Sci.
19:151-163.
5. Brown, A.G. 1960. The inheritance of shape, size and season in progenies of the cultivated apple. Euphytica 9:327-337.
6. Calzoni, G.L., A. Speranza, and N.Bagni.1979. In vitro germination of apple pollens. Scientia Horticulturae.
10:49-55.
7. Campo, G.D., I. Berregi, N. Iturriza, and J.I. Santos.
2006a. Ripeningand changes in chemical composition of seven cider apple varieties. Food Sci Tech Int.
12:477-487.
8. Campo, G.D., I. Berregi, R. Caracena, and J.I. Santos.
2006b. Quantitative analysis of malic and citric acids in fruit juices using proton nuclear magnetic reso- nance spectroscopy. Analytica Chimica Acta. 556:462- 9. Chun, O.K., D.O. Kim, N. Smith, D. Schroeder, J.T.468.
Han, and C.Y. Lee. 2005. Daily consumption of phe- nolics and total antioxidant capacity from fruit and vegetables in the American diet. J. Sci. Food Agric.
85:1715-1724.
10. Crane, M.B. and W.K.C. Lawrenece.1929. Genetical and cytological aspects of incompatibility and sterility in cultivated fruits. J. Pom. Hort. Sci. 7:276-301.
11. Crane, M.B. and W.K.C. Lawrenece. 1930. Fertility and vigor of apples in relation to chromosome num- ber. J. Genet. 22:153-162.
12. Crane, M.B. and W.K.C. Lawrenece.1933. Genetical studies in cultivated apples. Journal of Genetics.
28:265-296.
13. Darlington, C.D. and A.A. Moffett.1930.Primay and secondary chromosome balance in Pyrus. Jo. Genet.
22:129-151.
14. Drake, S.R. and T.A. Eisele. 1999. Carbohydrate and
acid contents of gala apples and bartlett pears from regular andcontrolled atmosphere storage. Journal of Agricultural and Food Chemistry 47: 3181?3184.
15. Drogoudi, P.D., Z. Michailidis, and G. Pantelidis.
2008. Peel and flesh antioxidant content and harvest quality characteristics of seven apple cultivars. Scien- tia Horticulturae 115:149-153.
16. Eccher, T.1986. Russeting and shape of ‘Golden Deli- cious’ apples as related to endogenous GA content of fruitlets. Acta Horticulture. 179:767-770.
17. Hagen, S.F., G.I.A. Bore, G.B. Bengtsson, W. Bilger, A. Berge, K. Haffner, and K.A. Solhaug. 2007. Phe- nolic contents and others health and sensory related properties of apple fruit (Malus domestica Borkh., cv.
Aroma): effect of postharvest UV-B irradiation. Post- harvest Biol. Technol. 45:1-10.
18. Hall, S.R. 1931. The problem of unfruitfulness in the cultivated apple. The American Naturalist. 65(701):
512-530.
19. Hulme, A.C. and M.J.C. Rhodes.1971. Pome fruit. In:
Hulme, A.C. (Ed.), The Biochemistry of fruits and their products.Academic press, London. 333-373.
20. Jaroslav, T. 1962. Free amino-Acids in apple pollen from the point of view of its fertility. Biologia Plan- tarum. 5(2):154-160.
21. Jonkers, H. 1971. An intenational experiment on juve- nility in apple. Euphytica. 20:57-59.
22. Jeuring, H.J., A. Brands, and P.V. Doorninck. 1979.
Rapid determination of malic and citric acid in apple juice by high performance liquid chromatography. Z.
Lebensm. Unters. Forsch. 168:185-187.
23. Kenis, K., J. Keulemans, and M.W. Davey.2008.Identi- fication and stability of QTLs for fruit quanlity traits in apple. The Genetics and Genomes. 4:647-661.
24. King, G.J., C. Maliepaard, J.R. Lynn, F. H. Alston, E.E. Durel, K.M. Evans, B. Griffon, F. Laurens, A.G.
Manganaris, E. Schrevens, S. Tartarini, and J. Ver- haegh. 2000. Quantitative genetic analysis and com- parison of physical and sensory descriptors relating to fruit flesh firmness in apple (Malus pumila Mill).
Theor Appl Genet. 100:1074-1084.
25. Larsen, P. and S.M. Tung. 1950. Growth-promoting and growth-retarding substances in pollen from dip- loid and triploid apple varieties. Botanical Gazette.
111(4):436-447.
26. Link, H. 2000. Significance of flower and fruit thin- ning on fruit quality. Plant Growth Regulation 31:17- 27. Lobit, L., M. Genard, B.H. Wu, P. Soing, and R.26.
Habib. 2003. Modelling citrate metabolism in fruits:
responses to growth and temperature. Journal of Experimental Botany 54(392):2489-2501.
28. McKnezie, D.W. 1971. A survey of shape variation in some New Zealand apples. New Zealand Journal of
Agricultural Research 14:491-498.
29. Miranda, C., T. Girard, and P.E. Lauri. 2007. Random sample estimates of tree mean for fruit size and colour in apple. Scientia Horticulturae. 112:33-41.
30. Moffett, A.A. 1931. A preliminary account of chromo- some behavior in the pomoideae. J. Pom. Hort. Sci.
9:100-110.
31. Muriel, B. and V. Roscoe. 1933. The chromosomal constitution of certain cultivated apple varieties. Jour- nal of Genetics. 28:157-167.
32. Noe, N. and E. Tommaso. 1996. Golden delicious apple fruit shape and russeting are affected by light conditions. Scientia Horticulturae. 65:209-213.
33. Ozaki, Y., K. Narikiyo, C. Fujita, and H. Okubo. 2004.
Ploidy variation of progenies from intra- and inter- ploidy crosses with regard to trisomic production in asparagus (Asparagus officinalis L.). Sex Plant Reprod. 17:157-164.
34. Petkovsek, M.M., F. Stampar, and R. Veberic. 2007.
Parameters of inner quality of the apple scab resistant and susceptible apple cultivars (Malus domestica Borkh). Sci. Hortic. 114:37-44.
35. Rodbotten, M., B.K. Martinsen, G.I. Borge, H.S.
Mortvedt, S.H. Knutsen, P. Lea, and T. Naes. 2009. A cross-cultural study of preference for apple juice with different sugar and acid contents. Food Quanlity and Preference. 27:277-284.
36. Sato, M. and K. Kanbe. 1989. Studies on the seed for- mation and early growth of seedlings from reciprocal crosses between the triploid apple cultivar ‘Mutsu’ and diploid cultivars. Bull. Akita. Pre. Coll. Arg. 15:121- 128 (In Janpanese with English summary).
37. Sato, M. 1990. Studies on the stabilization of fruit set of the triploid applecultivar ‘Mutsu’. Bull. Akita. Pre.
Coll. Agr. 16:1-52.
38. Sato, M. and K. Kanbe. 2007. Comparison of the Establishment efficiency of well-grown seedlings at the early growth stage from reciprocal crosses between
diploid and triploid apple cultivars, and the possiblil- ity of these cultivars cross breeding. Hort. Res. (Japan) 6:347-354.
39. Sato, M., T. Nyui, H. Takahashi, and H. Kanda. 2007.
Comparison of flowering and fruiting of seedlings from reciprocal crosses between diploid and triploid apple cultivars. J. Japan Soc. Hort. Sci. 76:97-102.
40. Shaw, J.K. 1914. A study of variation in apples. Mass.
Agricultural Experimental Station Bulletin. 149.
41. Shoemaker, J.S. 1926. Pollen development in the apple, with special reference to chromosome behavior.
Botanical Gazette. 81(2):148-174.
42. Sturm, K., M. Hudina, A. Solar, M.V. Marn, and F.
Stamper. 2003. Fruit quality of different ‘gala’ clones.
European Journal of Horticultural Science 68(4).
43. Tackholm, G. 1922. Zytologische studien über die Gat- tung Rosa. Acta. Hort. Berg. 7:97-381.
44. Visser, T. 1965. On the inheritance of the juvenile period in apple. Euphytica. 14:125-134.
45. Wand, S.J.E., K.I. Theron, J. Ackerman, and S.J.S.
Marais. 2006. Harvest and post-harvest apple fruit quality following applications of kaolin particle film in South African orchards. Scientia Horticulturae. 107:
271-276.
46. Wertheim, S.J. 1998. Rootstock guide: apple, pear, cherry, European plum. Fruit Research Station. Wil- helminadorp. Netherlands.
47. Whang, H.J., S.S. Kim, and K.R. Yoon. 2000. Analy- sis of organic acid in Korean apple juice by high per- formance liquid chromatography. J. Korean Soc. Food.
Sci. Nutr. 29:181-187.
48. Wu, J.H., H.Y. Gao, L. Zhao, X.J. Liao, F. Chen, Z.F.
Wang, and X.S. Hu. 2007. Chemical compositional characterization of some apple cultivars. Food Chem- istry 103:88-93.
49. Zhang, C.H. and S.M. Park. 2009. Aneuploid produc- tion from crosses with diploid and triploid in apple tree. Hort. Environ. Biotechnol. 50:203-207.
이배체와 삼배체를 교배하여 얻은 이수체의 생장과 과일 특성
장춘화·박성민* 강원대학교원예학과
적 요. 본연구는이전에이배체와삼배체를교배하여얻은이수체사과나무의생장과과실특성을알 아보고자수행하였다
.
이수체사과나무의화분발아율,
영양생장,
과실의특징을조사하였다.
그결과,
이수 체사과나무 간의화분발아율의차이가크고,
모든 이수체사과 나무의화분발아율이이배체‘
홍로’ (82.4%)
보다낮은것으로나타났다.
이수체사과나무과실의평균크기와중량은이배체‘
홍로’
보다작고가벼운것으로나타났다
.
또한이수체사과나무과실의길이와너비에의해이수체JF3942
빼고다른이수체과실의외형이조금납작했다
.
거의 모든이수체사과나무과일의당함량은이배체‘
홍로’
보다높았다
.
본실험에서얻은이수체사과의산함량은이전에대부분의연구자의이배체사과결과와완전히달랐다
.
이수체사과의시트르산하고타르타르산함량은사과산함량보다더높은것으로나타났 다.
이수체사과나무의수고와폭은이배체‘
홍로’
보다짧았다.
대부분이수체사과나무의수고는폭보 다길지만네나무는반대로나타났다.
잎의길이와너비에의해이수체사과나무잎의크기가이배체‘
홍로’
보다작았다.
이수체사과나무는이배체보다잘자라지않는것으로나타났다.
주제어 : 사과