pISSN 1738-6640 eISSN 2234-4020 http://dx.doi.org/10.6114/jkood.2016.29.4.061
Original Article / 원저
Effects of Torilis japonica Extract on DNCB-induced Contact Dermatitis in BALB/c Mice
Kyou-Young Lee
1)·Eun-Hye Song
2)·Chul-Hee Hong
1)*1 Dept. of Oriental Opthalmology and Otolaryngology and Dermatology, College of Korean Medicine, Sangji University
2 Dept. of Pharmacology, College of Korean Medicine, Sangji University
사상자 추출물이 DNCB로 유도된 BALB/c 마우스의 접촉성 피부염에 미치는 효과
이규영
1)·송은혜
2)·홍철희
1)*1 상지대학교 한의과대학 안이비인후피부과학교실 2 상지대학교 한의과대학 약리학교실
Abstract
목적: 본 연구에서는 TJ 열수추출물이 아토피 피부염에 미치는 효과를 확인하고자 하였다.
방법: 2, 4-dinitrochlorobenzen (DNCB)으로 BALB/c 마우스에 1차 감작 후 3주간 2차 감작을 시행하여 아토피 유사 피 부병변을 나타내는 접촉성 피부염을 유발한 뒤 동결건조 처리된 TJ 분말을 (PBS/EtOH/Cremophor=6:1:3)에 용해시켜 쥐의 등 피부에 10, 50 ㎎/㎖ 농도로 3주간 도포하였다.
결과: TJ 열수추출물은 아토피 피부 병변의 염증세포 침윤을 억제하였고 (10, 50 ㎎/㎖) 표피와 진피 두께를 회복시켰으며 (10, 50 ㎎/㎖), 혈청에서 히스타민 방출을 억제하였다 (10 ㎎/㎖). 또한 Th2 세포와 관련된 cytokine인 interleukin (IL)-4, IL-5, IL-13과 thymic stromal lymphopoietin (TSLP)의 mRNA 발현 양을 감소시켰고 (10, 50 ㎎/㎖), Th2 chemokine인 thymus- and activation-regulated chemokine (TARC or CCL17)의 mRNA의 발현 양을 감소시켰다 (10, 50 ㎎/㎖).
결론: TJ 열수추출물을 BALB/c 마우스에 외용하였을 때 항아토피 효과가 있음을 확인하였으며 따라서 TJ가 아토피 피부 염 치료의 외용제로 활용될 수 있을 것으로 사료된다.
Key words : Torilis japonica (TJ); Type 2 helper T (Th2); Atopic dermatitis (AD); Allergic contact dermatitis (ACD)
ⓒ 2016 the Society of Korean Medicine Ophthalmology & Otolaryngology & Dermatology
This is an Open Access journal distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/license/by-nc/3.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Ⅰ. Introduction
Allergic contact dermatitis (ACD) is a common occupational and environmental health issue. In common with other forms of allergy, this disease progresses in two phages; an initial stage during which sensitization is acquired, and if the sensitized individual is exposed subsequently, at the same or a different skin site, to the inducing chemical allergen, then a more vigorous secondary immune response will be provoked at the point of contact. This in turn initiates the cutaneous inflammatory reaction that is defined clinically as ACD
1).
2,4-dinitrochlorobenzene (DNCB) is an acute allergy-inducing substance. Atopic dermatitis (AD) mice model commonly uses chemical antibodes such as DNCB, 2,4-dinitrofluorobenzene and 2,4,6-trinitrochlorobenzene in order to cause atopy-like skin lesions. DNCB-induced mice model is frequently used for the investigation of AD
2,3). In the present study, acute ACD was induced by applying DNCB for 3 weeks of short period after first challenge, so skin lesions were atopy-like which is Th2 cell dominant.
AD, also called atopic eczema and allergic disorders, is a chronic inflammatory skin disease associated with cutaneous hyperreactivity to immunoglobulin E (IgE)-mediated sensitization
4). AD appears during babyhood and childhood but can last or start in adult
5). The prevalence of AD is 10–20% in children and 1–3% in adults
6). The
pathogenesis of AD can be attributed to complex interactions dependent on heritable and environmental factors
7,8).
The T cell plays important roles in allergic disorders
9). T helper cells, known as CD4+T cells, are subdivided into the type 1 helper T (Th1) and the type 2 helper T (Th2), which produce different profiles of cytokine
10). Pathognomonic features of AD include eczematous skin lesions with inflammatory cell accumulation, Th1/ Th2 balance skewed toward Th2
11). Th2 cells produce Th2 cytokines such as interleukin (IL)-4, IL-5, IL-13 and elevate serum IgE levels
11,12). IL-4 and IL-13 stimulate IgE production in B cell
13,14). IgE released from B cells binds to FcεRI, the high-affinity receptor for IgE of mast cell
15). Binding of an antigen by FcεRI-bound specific IgE leads to FcεRI clustering, which in turn induces downstream signaling and ultimately the release of mediators. The antigen complex IgE−
FcεRI on the surface of the mast cell induces mast cell degranulation
16). Mast cell releases histamine, leukotrienes, prostaglandins, chemokines and pro-inflammatory cytokines
17,18). IL-5 plays an important role in infiltration of eosinophil
19). The histology of AD is characterized by a dermal inflammatory infiltration containing eosinophils and mast cells
20). AD exhibits a compromised barrier function that leads to the activation of keratinocytes. Barrier disruption promotes the production of thymus- and activation-regulated chemokine (TARC or CCL17) from keratinocytes
21). Thymic stromal lymphopoietin (TSLP) is highly expressed in the epidermis of the skin lesions of AD patients and keratinocytes
22,23). Overexpression of TSLP by
Corresponding author : Chul-Hee Hong, School of Oriental Medicine, Sangji University, St. 80, Wonju city, Gangwon, 220-717, South Korea.
(Tel : 033-741-9266, E-mail : [email protected])
∙
Recieved 2016/10/6∙
Revised 2016/11/3∙
Accepted 2016/11/10keratinocytes results in an AD-like inflammatory phenotype in mice
24).
Corticosteroids and anti-histamines are drugs commonly available to treat AD
25,26). However, these drugs are accompanied by side effects that may include hypertension, osteoporosis, blurred vision, and dizziness
27). Calcineurin inhibitors also have been used topically and are considered quite safe
28), but prolonged application of these drugs is reported to cause skin cancer
29). Thus, development of alternative therapeutic agents is necessary for long-term therapy of AD.
The fruits of Torilis japonica (TJ) is 'Sa-Sang-Ja', which is traditional medical herbs prescribed as anti-fungal, anti-allergenic, antibacterial and sedative agent
30). In addition, several studies have demonstrated that TJ possess anti-inflammatory
31), anti-angiogenic
32), anti-invasive
33), and anti-microbial
34)effects. Although these biological effects of TJ have been investigated, there were no reports of AD. In this study, we examined the inhibitory effect of TJ on DNCB-induced contact dermatitis (CD) animal models, and through acute CD skin lesions effects on AD were investigated.
Ⅱ. Materials and Methods
1. Materials for Experiments 1) Reagents
1-Chloro-2, 4-dinitrobenzene, dexamethasone, acetone and all other chemicals were purchased from Sigma Chemical Co. (St. Louis, MO, USA).
Histamine ELISA kits were purchased from Enzo
Life Sciences (East Farmingdale, NY, USA).
Power SYBR® Green Master Mix was purchased from applied biosystems (Foster City, CA, USA).
Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) and IL-4, IL-5, IL-13, TARC, TSLP oligonucleotide primers were purchased from Bioneer (Daejeon, South Korea).
2) Preparation of aqueous extract of Torilis japonica (TJ)
TJ was purchased from Daehyo chamchowon (Suwon, Gyeong-gi, Korea). The dried fruit of TJ (400g) was extracted with 2800 ㎖ boiling distilled water for 2 h, and the extract was then concentrated under low pressure. After filtration, an aqueous solution of the extract was concentrated in a rotary evaporator, freeze-dried for five days, and stored in aliquots at 4 ℃. The yield of dried extract from starting crude materials was about 8.2% (w/w) for 32.8g dry weight.
3) Animals
BALB/c mice (female, 20-25g, 6 weeks old) were obtained from Daehan Biolink (Daejeon, South Korea). Animals were housed in an animal room at 23 ℃ with a relative humidity of 55%, a 12 h light/dark cycle. The mice were given an unlimited amount of water and food throughout the duration of the experiment. Animal care and experimental procedure were performed according to “the guide for the Care and Use of Laboratory Animal”, as adopted and promulgated by the
“Institutional Animal Care Committee, Sang ji
University” (Reg. No. 2016-03).
2. Experiments methods
1) Induction of CD skin lesions in BALB/c mice
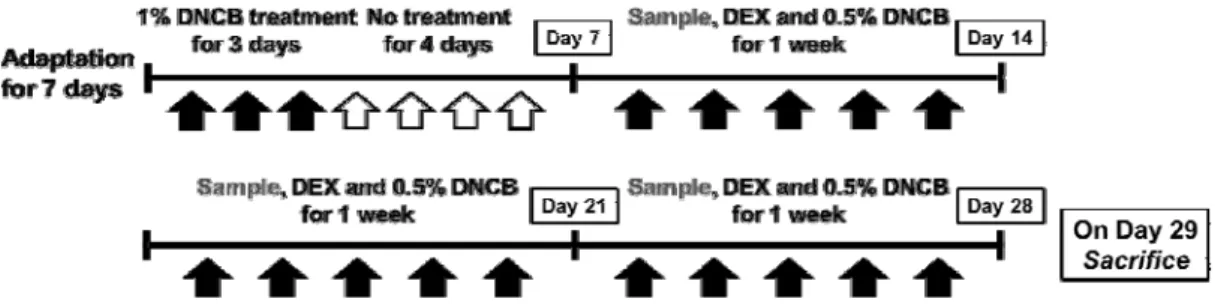
Animals were randomized into five groups (n=6 per group): Normal (mice treated with vehicle alone), DNCB (mice sensitized with DNCB, negative control), DNCB+DEX (mice sensitized with DNCB and treated with 10 µM dexamethasone, positive control) and DNCB+TJ 10 and 50 (mice sensitized with DNCB and treated with 10 and 50 ㎎/㎖ TJ).
After a 7 day adaptation period, the hair on the upper back was shaved and the experiment was immediately started. To induce CD skin lesions which were AD-like, mice were sensitized topically with 100 ㎕ of 1% DNCB in corn oil/acetone (1:4, v/v) for three days, followed by no treatment for four days. The same volume of vehicle(acetone/corn oil) was applied to the Normal group. After the first challenge, the treatment was repeated with 100 ㎕ of 0.5%
DNCB for 21 days. The DEX group and TJ group were respectively topically treated with 100
㎕ of DEX (10 µM, dissolved in PBS/EtOH/Cremophor=6:1:3) and 100 ㎕ of TJ (10, 50 ㎎/㎖, dissolved in PBS/EtOH/ Cremophor
Fig. 1. Schematic representation of the experimental design
=6:1:3) 4 h after DNCB treatment once a day, whereas Normal and DNCB groups were treated with vehicle (PBS/EtOH/Cremophor). Mice were sacrificed on day 29 of the experiment(Fig. 1).
Skin tissues from the back of the mice were excised and subjected to histological analysis, real time polymerase chain reaction examination.
2) Evaluation of dermatitis severity
Clinical dermatitis severity was tested using the method described by Yamamoto and colleagues
35). The severity of dermatitis was evaluated at experiment start day and end day. The development of erythema/hemorrhage, scarring/
dryness, edema, and excoriation/erosion was scored as follow: 0, none; 1, mild (<20%); 2, moderate (20-60%); and 3, severe (>60%). The sum of the individual scores was used as the dermatitis score.
3) Histological analysis of skin lesions
Upon euthanasia, skin samples from the dorsal
area were collected. The samples were fixed in
10% buffered formalin, embedded in paraffin,
sectioned into 4 ㎛ slices, and stained with
hematoxylin and eosin. Pathological changes
(epidermal, dermal hyperplasia, hyperkeratosis,
dermal edema, parakeratosis, vesicular formation, and inflammation) were assessed and compared between these groups. The images of the stained slides were observed using a DM IL LED microscope (Leica, Wetzlar, Germany) and photographed using a DFC295 (Leica, Wetzlar, Germany). Digital images were taken from each slide (2 per group), and measured using Leica Application Suite (Leica, Wetzlar, Germany).
4) Histamine assay
Blood from the orbital sinus was collected from each mouse at the end of the experiment. Serum was obtained by centrifugation at 1700 g for 30 min and stored at -80 ℃ until analysis. Serum levels of histamine in mice were measured using commercially available EIA kits (Enzo Life Sciences, Farmingdale, NY, USA) according to the manufacturer’s recommendations.
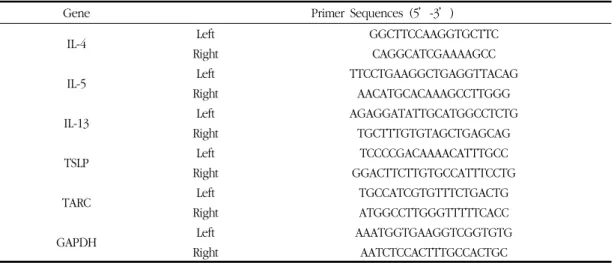
5) Quantitative real-time polymerase chain reaction (qRT-PCR)
The dorsal skin tissues were homogenized, and
Gene Primer Sequences (5’-3’)
IL-4 Left GGCTTCCAAGGTGCTTC
Right CAGGCATCGAAAAGCC
IL-5 Left TTCCTGAAGGCTGAGGTTACAG
Right AACATGCACAAAGCCTTGGG
IL-13 Left AGAGGATATTGCATGGCCTCTG
Right TGCTTTGTGTAGCTGAGCAG
TSLP Left TCCCCGACAAAACATTTGCC
Right GGACTTCTTGTGCCATTTCCTG
TARC Left TGCCATCGTGTTTCTGACTG
Right ATGGCCTTGGGTTTTTCACC
GAPDH Left AAATGGTGAAGGTCGGTGTG
Right AATCTCCACTTTGCCACTGC
Table 1. Primer Sequences Used for qRT-PCR Ampification
total RNA was isolated using the Easy-Blue®
Reagent according to the manufacturer’s instructions Kits (Intron Biotechnology, Seongnam, South Korea). Total RNA was quantified using an Epoch® micro-volume spectrophotometer system (BioTek Instruments;
Winooski, VT, USA). In brief, total RNA was converted to cDNA using a high-capacity cDNA reverse transcription kit (Applied Biosystems;
Foster City, CA, USA) and thermocycler (Gene Amp® PCR system 9700; Applied Biosystems) with the following program: initiation for 10 min at 25 ℃, followed by incubation at 50 ℃ for 90 min and at 85 °C for 5 min. Real-time PCR analysis was conducted using a Step One Plus®
Real-time PCR system (Applied Biosystems).
SYBR® Green master mix and primers were used for PCR analysis of GAPDH, IL-4, IL-5, IL-13, TARC or CCL17 and TSLP. The PCR cycling parameters were as follows: 10 min at 95 ℃; 40 cycles of 5 s at 95 ℃ and 45 s at 60 ℃; and a final melting curve of 15 s at 95 ℃, 1 min at 60
℃, and 15 s at 95 ℃. All primer sequences are
shown in Table 1. Gene expression was calculated according to the comparative threshold cycle (Ct) method.
6) Statistical analysis
All the values reported have been expressed as the mean ± standard deviation (SD). Data were analyzed using one-way analysis of variance (ANOVA) with Dunnett’s test, and p-values <
0.05 were considered statistically significant. All statistical analyses were performed using GraphPad Prism (version 5.00 for Windows, San Diego, CA, USA).
Ⅲ. Results
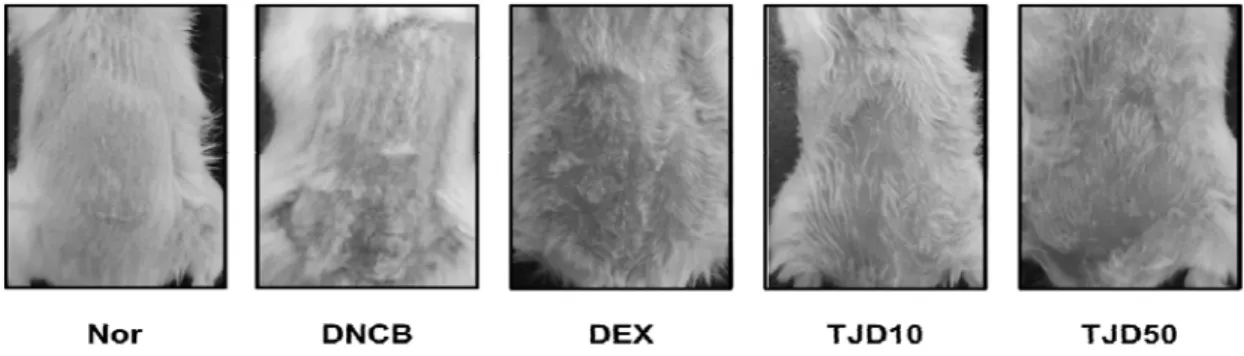
1. Effects of TJ on clinical features of skin lesions in DNCB-induced CD
Clinically, topical exposure of DNCB results in skin symptoms of itching, erythema, and hemorrhage after a week, then it rapidly progresses into edema, superficial erosion, deep excoriation, scaling, and dryness of the skin within two weeks following the repeated treatment with DNCB
11,36). As shown in Fig. 2, repeated topical application of DNCB induced
Fig. 2. Effects of TJ on clinical features of skin lesions in DNCB induced CD
erythema, superficial erosion, excoriation, scaling, and dryness. However, treatment with TJ inhibited these skin symptoms. Representative photographic images of mice from different treatment groups at the time of a sacrifice. Nor:
Normal control; DNCB: DNCB treated; DEX:
Dexamethasone treated with DNCB; TJD 10: TJ 10 ㎎/㎖ treated with DNCB; TJD 50: TJ 50 ㎎/㎖
treated with DNCB.
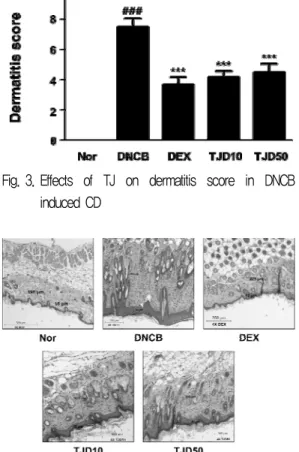
2. Effects of TJ on dermatitis score in DNCB-induced CD
Skin conditions were evaluated at experiment start day and end day using a dermatitis scoring method. The dermatitis scores of DNCB group increased compared with those of the Normal group. However, in DEX and TJ treated mice, the dermatitis scores were lower than DNCB group(Fig. 3).
Dermatitis scores were measured at experiment start day and end day. The dermatitis score was defined as the sum of scores graded as 0 (none), 1 (mild), 2 (moderate), 3 (severe) for each of the four symptoms (erythema/hemorrhage, scarring/
dryness, edema, and excoriation/erosion). Data are
presented as mean±standard deviation (SD; n=6).
Data were analyzed using one-way analysis of variance followed by Dunnett’s post hoc test.
***P < 0.001 versus DNCB group; ### P < 0.001 versus Normal group.
3. Effects of TJ on histological analysis in DNCB-induced CD
In addition, we examined application of TJcould affect the interrupted skin barrier using H&E staining. Histological features of the skin lesions showed epidermal, dermal hyperplasia and
Fig. 3. Effects of TJ on dermatitis score in DNCB induced CD
Fig. 4. Effects of TJ on histopathological feature of skin lesions in DNCB induced CD
Tissues were excised, fixed in 10% formaldehyde, embedded in paraffin, and sectioned. The sections were stained with hematoxylin and eosin (H&E) (magnification, 4x).
accumulation of inflammatory cells in the dermis/epidermis of DNCB group compared with Normal group. In contrast, TJ treated group exhibited less of these changes in DNCB-induced mice(Fig. 4).
4. Effects of TJ on histamine release in DNCB-induced CD
Mast cells play a key role in allergic reactions
37). Mast cells secrete inflammatory mediators such as histamine
38). Histamine, a major component of mast cells, is a symptom inducing substance in AD
36). Histamine is involved in induction of itching and edema
39). In order to evaluate whether TJ can modulate degranulation of mast cells, histamine assay was performed using EIA kit. The histamine release in the serum markedly increased in the DNCB group compared with Normal group, whereas TJ treatment significantly suppressed histamine levels. Treatment with DEX also attenuated the changes in histamine levels(Fig. 5).
Fig. 5. Effects of TJ on histamine release in DNCB-induced CD
Histamine concentrations were measured in mouse serum by ELISA. Data are presented as mean±standard deviation (SD;
n=6). Data were analyzed using one-way analysis of variance followed by Dunnett’s post hoc test. *P < 0.05 and **P < 0.01 versus DNCB group; ### P < 0.001 versus Normal group.
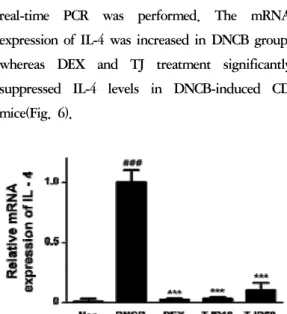
5. Effects of TJ on the mRNA expression of IL-4 in DNCB-induced CD
IL-4 promotes differentiation of Th2 cells and IgE class switching by B cells. Its receptor has been associated with AD
40). To investigate the effect of TJ on the mRNA expression of IL-4, real-time PCR was performed. The mRNA expression of IL-4 was increased in DNCB group, whereas DEX and TJ treatment significantly suppressed IL-4 levels in DNCB-induced CD mice(Fig. 6).
Fig. 6. Effects of TJ on mRNA expression of IL-4
Total RNA was prepared for the real-time PCR analysis of IL-4 gene expression from skin tissues. Real-time PCR analysis was conducted using a Step One Plus® Real-time PCR system. Data are presented as mean±standard deviation (SD; n=6). Data were analyzed using one-way analysis of variance followed by Dunnett’s post hoc test. ***P < 0.001 versus DNCB group; ###
P < 0.001 versus Normal group.
6. Effects of TJ on the mRNA expression of IL-5 in DNCB-induced CD
IL-5 produced by Th2 cells, induces eosinophil differentiation, activation, mobilization, and survival
41). Eosinophils are important mediators of the inflammatory process in AD, so agents that block IL-5 might be developed as therapeutics
42). In this study, the mRNA levels of IL-5 was increased in DNCB group compared with Normal group, but TJ inhibited the mRNA levels of IL-5
in DNCB induced CD mice(Fig. 7).
Fig. 7. Effects of TJ on mRNA expression of IL-5
Total RNA was prepared for the real-time PCR analysis of IL-5 gene expression from skin tissues. Data are presented as mean±standard deviation (SD; n=6). Data were analyzed using one-way analysis of variance followed by Dunnett’s post hoc test. ***P < 0.001 versus DNCB group; ### P < 0.001 versus Normal group.
Fig. 8. Effects of TJ on mRNA expression of IL-13
Total RNA was prepared for the real-time PCR analysis of IL-13 gene expression from skin tissues. Data are presented as mean±standard deviation (SD; n=6). Data were analyzed using one-way analysis of variance followed by Dunnett’s post hoc test. ***P < 0.001 versus DNCB group; ### P < 0.001 versus Normal group.
7. Effects of TJ on the mRNA expression of IL-13 in DNCB-induced CD
IL-13 is known as a major stimulator of
inflammation and tissue remodeling at sites of
Th2 inflammation, such as in asthma
43). To
investigate the effects of TJ on the mRNA
expression of IL-13, real-time PCR was
performed. Our data indicated that the mRNA
levels of IL-13 was increased in DNCB group compared with Normal group, but TJ inhibited the mRNA levels of IL-13 in DNCB induced CD mice(Fig. 8).
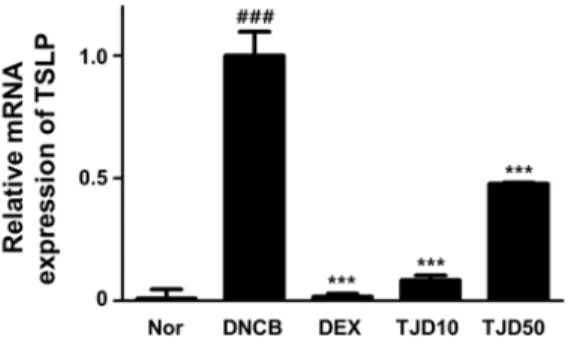
8. Effects of TJ on the mRNA expression of TSLP in DNCB-induced CD
TSLP represents a key molecule at the epithelial cell-dendritic cell (DC) interface to initiate allergic inflammation
44). To investigate the effect of TJ on the mRNA expression of TSLP, real-time PCR was performed. In this study, the TSLP was significantly higher in the DNCB-induced group than in the Normal group. In the TJ-treated group, it was significantly lower than that observed in the DNCB-induced group(Fig. 9).
Fig. 9. Effects of TJ on mRNA expression of TSLP
Total RNA was prepared for the real-time PCR analysis of TSLP gene expression from skin tissues. Data are presented as mean±standard deviation (SD; n=6). Data were analyzed using one-way analysis of variance followed by Dunnett’s post hoc test. ***P < 0.001 versus DNCB group; ### P < 0.001 versus Normal group.
9. Effects of TJ on the mRNA expression of TARC in DNCB-induced CD
Keratinocytes are inflammatory sites of epidermis which induce abnormal expression of cytokines and chemokines including Th2
chemokines such as TARC
45). Therefore, we examined the effects of TJ on the mRNA expression of TARC. As shown in Fig. 10, the TARC was increased in the DNCB-induced group compared with Normal group. The DEX and TJ-treated groups exhibited less of an increase in the levels of TARC mRNA.
Fig. 10. Effects of TJ on mRNA expression of TARC
Total RNA was prepared for the real-time PCR analysis of TARC gene expression from skin tissues. Data are presented as mean±standard deviation (SD; n=6). Data were analyzed using one-way analysis of variance followed by Dunnett’s post hoc test. ***P < 0.001 versus DNCB group; ### P < 0.001 versus Normal group.