대한소화기학회지 2002;39:4 - 12
서 론
이동성 위장관복합운동(migrating motor complex, MMC)
은 공복 상태에서 위와 십이지장 근위부에서 시작하여 말단 회장부까지 움직여 내려가는 위장관 벽의 전기 및 기계적 활동의 집합을 지칭한다. 이 현상의 생리적 의의는 공복기 에 위와 소장에 남아있던 음식물이나 탈락된 상피세포 등 을 일시에 청소하여 다음 번 음식을 받아들이기 용이하게 하는 역할을 하며 입과 반대 방향으로 분당 5 cm의 속 도로 내려가며 약 1.5~2시간 간격으로 발생한다. MMC는 정지기인 phase I에서 시작하여 불규칙한 전기 및 기계적 활성을 보이는 phase II로 이어지고, 규칙적인 전기 및 기계적 활성을 보이는 phase III로 끝난다 .1 ,2 모틸린
모틸린에 의한 사람 위 평활근의 수축 기전에 관한 연구
성균관대학교 의과대학 마산삼성병원 내과학교실 , 삼성서울병원 내과학교실 *, 삼성제일병원 내과학교실†, 생리학교실‡, 경상대학교 의과대학 내과학교실§
심상군・이종철 *・이풍렬 *・최규완 *・전성국
†・강동묵
‡・엄대용
‡・이종석
§・성인경・김현서
Me c h a n i s m s o f Mo t i l i n A c t i o n o n S m o o t h Mu s c le o f th e H u m a n S to m a c h
S a n g Go o n S h i m , M .D ., J o n g C h u l R h e e , M .D .*, P o o n g -L y u l R h e e , M .D .*, K y o o Wa n C h o i , M .D .*, S u n g K o o k J e o n , M .D .†, T o n g M o o k K a n g , P h .D .‡,
D a e -Y o n g U h m , M .D ., P h .D‡, J o n g S e o k L e e , M .D .§, In K y u n g Su n g , M .D ., a n d H y u n S e o K i m , M .D .
Dep artment of Internal Medicine, Masan Samsumg Hosp ital, Masan; Dep artment of Internal Medicine *,
Samsung Medical Center and Samsung Cheil Hosp ital†, Seoul; Dep artment of Physiology§, Sungkyunkwan University School of Medicine, Suwon; Dep artment of Internal Medicine‡, Gyeongsang National University College of Medicine, Chinj u, Korea
Background/Aims: Motilin is an intestinal peptide that stimulates the contraction of gut smooth muscle. A discrepancy exists between the in vivo (neurally mediated) and in vitro (direct action on a smooth muscle receptor) mechanisms of motilin action in many species. We investigated in vitro mechanisms of motilin action on human gastric smooth muscle.
Methods: Antral cirular muscle strips of the surgical tissue obtained during gastrectomy, were used to measure contractile force and electrical activity. Dispersed muscle cells were used to measure L-type Ca
2 +current and electrical activity. Results: Motilin of 1-100 nM contracted smooth muscle in a concentration-dependent manner . Motilin-induced contractions were unaffected by tetrodotoxin or atropine treatment. Nifedipine or Ca
2 +-free bath solution blocked motilin (10 nM)-induced contractions. Low concentration of motilin (1 nM) resulted in an increase in acetylcholine (0.1~100 M)-induced contractions. By patch clamp recording technique, motilin (1 or 10 nM) did not modify the L-type Ca
2 +current, but motilin-induced membrane depolarization was detected. Erythromycin also contracted smooth muscle with membrane depolarization but verapamil inhibited the contraction. Conclusions: These results suggest that motilin contracts smooth muscle through a direct action on smooth muscle receptor and Ca
2 +influx through the L-type Ca
2 +channel, which is due to membrane depolarization, also mediates motilin-induced contractions. (Korean J Gastroenterol 2002 ;39 :4-12)
Key Words: Motilin, Atropine, L-type calcium channel, Membrane depolarization
접수: 2001년 5월 19일, 승인: 2001년 10월 10일 연락처 :이종철, 135-230, 서울시 강남구 일원동 50,
삼성서울병원 소화기내과
Tel: (02) 3410-3409, Fax: (02) 3410-3849 E-mail: jchrhee @smc.samsung.co.kr
※본 연구는 삼성생명공학연구소 연구비(C-95-002-6) 지원을 받아 시행했음 .
심상군 외 9인 . 모틸린에 의한 사람 위 평활근의 수축기전에 관한 연구
(motilin)은 22개의 아미노산으로 구성된 펩타이드로서 주로 십이지장과 공장점막에 있는 장크롬친화 (entero- chromaffin)세포로부터 공복시에 매 1.5~2시간 간격으로 주 기적으로 분비되어 위와 소장에서 강력한 연동파를 일으 키며 phase II 말기 및 phase III 기간에 일어난다.1-3 따라 서 혈중 모틸린 농도의 증가는 위와 소장의 위상성 및 긴 장성 수축의 증가와 시간적으로 일치하여 나타난다. 또한 외부에서 투여된 모틸린이 개와 사람의 위에서 MMC를 유 발함이 보고되었다.4 - 7
현재 모틸린의 수축기전에 대해서는 시험관내(in vivo) 및 생체(in vitro)실험에서 얻어진 결과가 서로 다르고, 시 험관내 실험일지라도 실험 조건이나 동물 종류, 위장관 부위에 따라 다른 작용기전이 제시되었다. 생체에서 모틸 린의 효과는 아트로핀(atropine)을 처리하거나 미주신경을 절단할 경우 억제되기 때문에 흥분성 신경계를 자극하여 위장관운동의 항진을 유도한다고 보고되었다.8- 10 반면, 시험 관내 연구에서는 모틸린에 의한 근수축이 항콜린성 약제나 신경차단제인 복어독 (tetrodotoxin, TTX) 등에 억제되지 않 아 모틸린이 평활근 세포에 직접 작용할 것이라고 하였
다 .1 1- 13 그럼에도 불구하고 토끼와 닭을 이용한 일부 시험
관내 연구에서 모틸린이 신경 자극을 통해서 위장관의 평 활근을 수축시킨다고 하였고14 - 1 8 또한 저농도 모틸린은 내 재신경의 콜린성 흥분을 통해서 근수축을 일으키고 고농 도는 직접 평활근세포를 자극한다고 보고하였다 .19 본 연 구에서는 사람의 위전정부 근육을 얻어 시험관 내에서 모 틸린의 수축력을 측정하고 모틸린이 어떠한 작용기전에 의하여 수축을 일으키는지 알아보고자 하였다 .
대상 및 방법
1 . 평활근 절편 (is o la t e d mus c le s t rips )의 수축력 측정 위유문부 평활근 조직은 삼성서울병원 수술실에서 위 절제술을 받은 위암 환자로부터 얻었으며 삼성의료원 임 상시험위원회의 승인를 받았다. 수술장에서 얻은 근육은 bicarbonate- buffered Tyrode용액에 넣어 즉시 실험실로 옮 겨 실험에 사용하였다. Tyrode용액에 95% O2와 5% CO2의 혼합가스를 지속적으로 공급하면서 조직절편으로부터 점 막층을 박리한 후 근육만을 실험에 사용하였다 . 길이 10~12 mm, 폭 2~3 mm의 윤상근 절편을 잘라 수직형 organ bath (36.5℃) 내에 위치시켰는데 절편의 한쪽 끝은 백금으 로 만든 고리에 실로 묶어 고정하였고 다른 한쪽은 등장 성 수축력측정기에 연결하여 수축력을 측정하였다 . 최적 의 수축력을 측정하기 위해 실험 전에 1 g의 장력을 근육 에 걸어주고 1시간 이상 안정화시킨 후 본 실험을 수행하 였다 . 모틸린은 반복 혹은 지속 투여시 탈감작 현상이 있
으므로 농도 변화에 대한 수축력 측정을 위해 비누적 농 도 -반응곡선을 얻었다. 아세틸콜린 10 µM을 반복적으로 2 ~3회 투여하여 안정적인 수축을 얻은 후 각 근절편에 단일 농도의 모틸린을 투여하여 얻은 수축의 크기를 아 세틸콜린에 의한 수축력으로 나누어 표준화하였다 . 모틸 린에 의한 수축이 내재신경 혹은 콜린성 신경전달물질의 개입 여부를 알기 위해 복어독 및 아트로핀을 모틸린(10 nM) 투여 전후에 처리하고 수축력의 변화를 관찰하였으 며 이 때 아세틸콜린 혹은 카바콜 (carbachol, CCh) 10 µM 에 의한 수축력과 비교하여 표준화한 후 도시하였다 . 아 세틸콜린에 의한 농도 -반응 곡선은 한 조직절편에서 아 세틸콜린의 농도를 올리면서 수축을 유발한 때 얻어진 수축정도를 농도-반응곡선이 얻어지기 전에 얻어진 아세 틸콜린 10 µM에 의한 수축력의 크기로 나누어 표준화한 후 도시하였다 .
2 . 유리 미세전극을 이용한 세포막 전압 측정
길이 10~12 mm, 폭 2~3 mm의 근절편을 만들어 용량 2 mL의 수평형 organ bath에 위치시켰다 . Organ bath 바닥에 부착된 실리콘 판에 가는 금속 핀을 이용하여 근절편의 한쪽 끝을 고정하여 근육이 움직이지 못하도록 하고 다른 한쪽 끝은 실크 실을 이용하여 고리를 만든 후 등장성 수 축력측정기에 연결하여 나머지 근절편 부위에서 일어나는 근육의 수축을 측정할 수 있도록 설계하였다. 근절편은 펌프를 이용하여 2~3 mL/ min의 속도로 95% O2, 5 % CO2
혼합가스가 공급되는 Tyrode용액을 관류하였으며 온도는 36.5℃로 유지하였다. 수평형 puller (PUL- 1, WPI, FL, USA) 를 이용하여, 전극 끝에 40~60 MΩ의 저항을 갖는 가는 유리 미세전극을 뽑은 후에 3 M KCl을 채우고 미세조정 기를 이용하여 핀으로 고정된 절편 부위의 평활근세포에 찔러 넣어 세포막 전압을 측정하고 증폭기로 증폭한 후 피지오그래프에 기록하였다 . 이때 근 수축력의 변화를 막 전압의 변화와 시간적으로 동시에 기록하여 막전압의 변 화가 근 수축에 미치는 효과를 관찰하였다 .
3 . 단일 평활근세포의 분리 (s ing le s m o o t h m u s c le c e ll is o la t io n )
칼슘이 함유되지 않은 Tyrode용액에 papain (1 mg/mL), 우혈청 알부민(bovine serum albumin, 2 mg/mL) 및 dith- iothreitol (DTT, 1 mg/mL)를 녹여 효소용액을 만든 후 근 조직절편을 넣고 37℃에서 5분간 소화시켰다. 위의 효소 용액을 이용하여 1차 소화시킨 조직절편을 3.0~3.5 mg/mL 의 콜라겐분해효소(Wako, Osaka, Japan), 우혈청 알부민(3 mg/mL) 및 DTT (1 mg/mL)가 함유된 무칼슘 Tyrode용액을 이용하여 2차 소화시켰다. 2차 소화된 조직을 Kraft-Brche
5
The Korean Journal of Gastroenterology : Vol. 39. No. 1. 2002
(KB)용액에 옮기고 끝이 부드러운 파스퇴르 전극으로 부 드럽게 진탕하여 단일 평활근세포를 분리하였다. 분리된 세포는 KB 용액에 넣어 4℃에 보관하였으며 세포 분리 후 10시간 내에 실험하였다.
4 . 칼슘전류의 측정
효소 처리를 통해 분리된 단일 평활근세포를 도립 현 미경(Diaphot 300, Nikon, Tyokyo, Japan) 위에 설치된 기록 chamber 내에 뿌린 후 중력을 이용하여 bath용액을 2 mL/min의 속도로 흘려주면서 whole-cell patch clamp 방법2 0 으로 칼슘전류를 기록하였다. 이온전류는 실온(24℃)에서 기록하였다 . 수직형 puller (PP-83, Narishige, Tokyo, Japan) 를 이용하여 피펫 끝의 저항이 3~4 MΩ되는 patch clamp용 유리 미세전극을 제작하였다 . 유리 미세전극에 이온전류 기록에 필요한 내부 용액을 채운 후 patch clamp용 증폭기 (Axopatch- 1D, Axon Ins., Foster City, CA, USA)와 연결된 headstage에 연결한 후 미세조정기를 이용하여 세포에 접 근시켰다 . 피펫의 끝 부분에 4 mg/mL의 nystatin이 함유된 피펫 용액을 먼저 채운 후 나머지 부분에 nystatin이 없는 피펫 용액을 마저 채운 후 nystatin-perforated whole-cell mode에서 칼슘전류를 기록하였다. 발생된 이온전류는 증 폭기에 내장된 5 kHz (-3 dB frequency) 여과기를 통하여 여 과 후 기록하였으며 오실로스코프 (PM 3350A, Phillips, Eindhoven, Netherland)와 컴퓨터를 통해 관찰 및 저장하였 다 . 이온전류의 기록 및 분석을 위하여 pCLAMP 6.0.2 프 로그램(Axon Ins.)과 Origin 5.0 프로그램(Microcal Software, Inc ., Northhampton, MA, USA)을 이용하였다 .
5 . 실험용액 및 약물
근절편의 제작 및 수축력 측정에 이용된 bicarbonate- buffered 용액은 NaCl 116 mM, KCl 5.4 mM, NaHCO3 24 mM, NaH2PO4 1 mM, MgCl2 1 mM, CaCl2 2 mM, 포도당 5.6 mM이 함유되었으며, pH는 HCl을 이용하여 7.35로 맞 추었다. 효소용액의 제작에 필요한 생리적식염수(physiolo- gical salt solution, PSS)의 조성은 HEPES (hydroxyethylpiper- azineethanesulfonic acid) 10 mM, NaCl 135 mM, KCl 5 mM, MgCl2 1 mM, 포도당 10 mM이 함유되었으며, pH는 tris (tris aminomethane base)를 이용하여 7.35로 맞추었다 . 칼슘 전류 기록을 위해 Ca2 +-free PSS에 BaCl2 10 mM을 첨가하 였고 피펫 용액은 EGTA (ethyleneglycol-bis-aminoethyle- thertetraacetic acid) 10 mM, CsCl 110 mM, TEA (tetraethy- lammonium chloride) 20 mM, HEPES 10 mM, Na2ATP 3 mM, MgCl2 3.5 mM이 함유되었으며, pH는 tris를 이용하여 7.25로 맞추었다. KB 용액은 L-glutamic acid 50 mM, KCl 40 mM, taurine 20 mM, KH2PO4 20 mM, MgCl2 3 mM, 포도당 10 mM,
HEPES 10 mM, EGTA 0.5 mM 함유되었으며, pH는 KOH를 이용하여 7.35로 맞추었다. 실험에 사용된 시약 중 복어독 과 nifedipine은 Research Biochemical International (Natick, MA, USA)에서, 아세틸콜린, 카바콜, 돼지 모틸린, 아트로 핀 그리고 기타 시약들은 Sigma (St. louis, MO, USA)로부 터 구입하여 사용하였다.
6 . 자료 분석
데이터는 평균 ± 표준오차(n=검체수)로 표시하였으며 통 계적 유의성은 Student's t test를 이용하였다(p<0.05). 아래 logistic sigmoid function을 이용하여 약물의 농도-반응곡선 으로부터 EC5 0(half effective concentration)을 구하였다.
F/Fm a x = {1+([EC5 0]/ [A]h)- 1
F는 투여한 약물에 의해 발생된 근육의 수축력 크기를,
Fm a x는 약물에 의한 최대 수축을 표시한다. EC5 0는 Fm a x의
50 % 크기의 수축을 일으키는 약물 A의 농도이며 h는 곡 선의 기울기를 나타낸다.
결 과
1 . 모틸린에 의한 평활근 수축기전
Fig. 1. Motilin-induced contractions of human antral smooth muscle and dose-response curve. (A-D) Each concentration of motilin (1, 10, 50 and 100 nM) contracted the muscle.
6
Shim, et al. Mechanisms of Motilin Action on Smooth Muscle of the Human Stomach
모틸린 1nM에서 평활근의 수축반응이 거의 나타나지 않 았으나 10, 50, 100 nM에서는 강력한 위상성 수축을 보였다 (Fig. 1). 농도-반응곡선에서 모틸린은(1~100 nM) 농도 증가에 비례하여 수축력이 증가하였으며 EC5 0는 5.63 nM였다(Fig. 2).
복어독 1µM과 아트로핀 1µM을 전처리(A) 또는 후처리
Fig. 4. Summary of effects of TTX and atropine on the motilin-induced contraction. Data were collected from Fig. 2.
Pre- (A, B) or post-treatment (C) of atropine or TTX did not significantly change the amplitudes of motilin-induced contraction (p>0.05). The number above each bar indicates the sample size.
(B, C)한 경우 모틸린 10 nM에 의한 근 수축의 변화는 관 찰되지 않았다(Fig 3). Fig. 4는 Fig. 3를 그래프로 요약한 Fig. 2. Motilin-induced contractile forces. It was normalized by
10 µM acetylcholine (ACh), and plotted against motilin concentration. From the fitted line, EC5 0 value of motilin was 5.6 nM. The number in the parenthesis indicates the number of tested muscle strips.
Fig. 3. Effects of tetrodotoxin (TTX, 1 µM) and atropine (1 µM) on the motilin-induced contraction. TTX and atropine were treated before (A, B) and after (C) the administration of motilin. TTX and atropine did not block the motilin-induced contraction.
7
대한소화기학회지 :제39권 제 1호, 2002
것이며 모틸린과 대조군 간 수축 정도는 유의한 차이가 없었고, 복어독과 아트로핀 후처리하여도 두 군 간 수축
크기의 변화는 없었다 .
평활근세포에 존재하는 것으로 증명된19 L-형 막전압의 존성 칼슘통로를 차단하는 nifedipine을 처리하거나 세포 밖 칼슘을 제거하고 그에 따른 모틸린 수축반응의 변화를 관 찰하였다(Fig. 5). 모틸린 10 nM에 의한 근수축은 nifedipine 1 µM의 투여로 완전히 억제되었으나 지속적 긴장성 수축은 억제되지 않았다. Nifedipine 전처치 후 모틸린의 수축 효과 는 위상성 수축 없이 일과성의 수축만을 보였다. 세포밖 칼슘을 제거한 후 아세틸콜린을 투여하여 세포내 칼슘 저 장고로부터 유리될 수 있는 칼슘을 고갈시킨 다음 모틸린 을 투여하면 아세틸콜린은 위상성 수축이 없이 일과성의 단일 수축만 유도하였다.
2 . 저농도 모틸린에 의한 아세틸콜린 수축의 항진 모틸린 1 nM에 의해 수축반응이 없는 근조직에서 아세 틸콜린을 추가 투여하면 아세틸콜린에 의한 수축이 현저히 항진되고 농도의존적으로 수축력이 증가하였다(Fig. 6). 대 조군과 모틸린 투여군의 EC5 0는 각각 4.1 µM, 1.37 µM였 다 .
아세틸콜린은 위 평활근에 위상성 수축과 긴장성 수축 을 모두 일으키는데 , 그 양상은 지속되는 긴장성 수축 위 에 위상성 수축이 겹쳐서 나타난다 (Fig. 7). 아세틸콜린에 의한 위상성 수축은 L-형 칼슘통로 차단제인 nifedipine에 의해 완전히 억제되었으나 지속적인 긴장성 수축은 nifedipine 에 의해 전혀 차단되지 않았다 . 모틸린 1 nM의 전처리에 의 Fig. 5. Blocking effects of nifedipine and removal of
extracellular Ca2 + on the motilin-induced contraction. (A) Motilin- induced contraction was almost completely inhibited by 1 µM nifedipine. (B) Application of 10 nM motilin in the presence of nifedipine produced only a small transient contraction. (C) After the removal of extracellular Ca2 +, a transient contraction by 10 µM ACh was recorded, but motilin did not generate any appreciable contraction.
Fig. 6. Potentiation of acetylcholine (ACh)-induced contraction by 1 nM motilin. (A) ACh contracted the muscle dose-dependently (left panel), and the contractions were augmented by pre-treatment with 1 nM motilin (right panel). This muscle strip was not contracted by 1nM motilin. (B) Dose-response curves of ACh- induced contraction in the absence (filled circles) and/or presence of 1 nM motilin (filled squares). The estimated EC5 0 values from the two curves were 4.1 µM and 1.37 µM, respectively (n=17, p<0.05).
Fig. 7. Potentiation of ACh-induced contraction by 1 nM motilin . (A) The ACh-induced, nifedipine-sensitive phasic contraction was potentiated by 1 nM motilin (left trace).
Nifedipine (1 µM) selectively inhibited the phasic con- traction produced by 10 µM ACh (middle trace). (B) The effects of 1 nM motilin on the phasic and tonic contractions that are nifedipine-sensitive. n=5, *p<0.05.
8
심상군 외 9인 . 모틸린에 의한 사람 위 평활근의 수축기전에 관한 연구
해 위상성 수축은 1.26 ± 0.17 g에서 1.62 ± 0.22 g으로 증가 하였으나(P<0.05), nifedipine에 의해 차단되지 않는 긴장성 수축은 대조군(0.2 ±0.10 g)과 모틸린 처리(0.18 ± 0.07 g)로 유의한 차이가 없었다 .
3 . 칼슘통로에 대한 모틸린의 효과 ( F ig . 8 )
단일 평활근세포를 이용하여 patch clamp 방법으로 칼 슘전류를 직접 기록하고 칼슘전류에 대한 모틸린의 항진 효과를 관찰하였다 . 모틸린 1 nM 투여 그룹에서, +10 mV 에서 얻어진 최대 칼슘전류 값은 투여 모틸린 1 nM 투여 전 -168.6 ± 14.4 pA, 투여 후 -152.0 ± 9.3 pA로서 유의한 차이가 없었다(p>0.05). 또한 모틸린 10 nM 투여 그룹은 + 10 mV에서 얻어진 최대 칼슘전류 값은 투여 전 -125.2
± 9.0 pA, 모틸린 10 nM 투여 후 -116.7 ± 9.0 pA로 통계적 유의성이 없었다 .
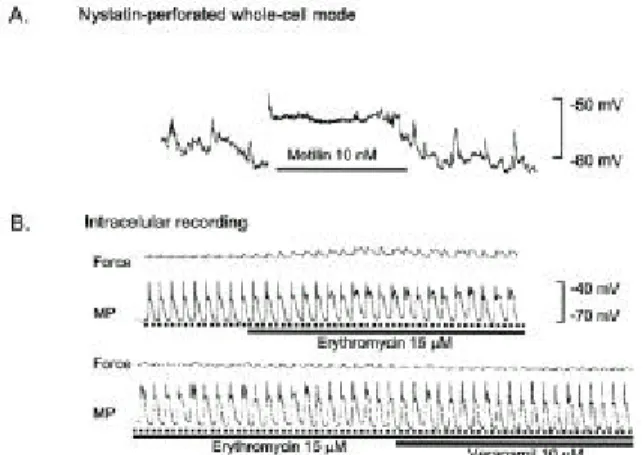
4 . 평활근 세포막 전압에 대한 모틸린과 e ryt h ro myc in ( EM)의 효과 ( Fig . 9 )
단일 평활근세포에서 nystatin-perforated법을 이용하여 세포막 전압을 측정한 결과 모틸린 10 nM은 막전압을 현 저하게 탈분극(depolarization)시켰다. 근조직에서 세포내 전 극기록법을 이용하여 EM 15 µM 투여는 평활근 세포막의 탈분극과 고원기(plateau)에서 가시전압(spike potentials)이 보였다. 이는 통상 칼슘전류에 기인하므로 EM 투여 후 탈분극 및 칼슘전류의 항진을 시사한다. 탈분극 및 가시 전압의 발생과 일치하여 근수축의 증가가 관찰되었으며 이런 변화는 L-형 칼슘통로 차단제인 verapamil 10 µM에 의해 효과적으로 억제되었다 .
고 찰
본 연구는 시험관 내에서 사람의 평활근을 이용하여 모틸 린의 근수축 및 그 기전에 대해 알아보았다 . 모틸린은 농 도의존적으로(1~100 nM) 사람 위 평활근의 수축을 증가시 켰고 농도 측면(EC5 0)에서 아세틸콜린에 비해 약 1000배 정도 강력한 수축을 유발함으로써 생리적으로 매우 중요 하고 강력한 위장관운동조절호르몬임을 알 수 있었다 . 세 부적인 근육 수축기전을 알기 위해 먼저 근절편 내에 함 유된 위장관 내재신경의 자극을 통한 효과인지 혹은 콜린 성 신경전달물질의 분비에 의한 것인지를 검증하기 위해 복어독이나 아트로핀을 투여하였지만 모틸린에 의한 수축 Fig. 8. Effects of motilin on the L-type Ca2 + current. (A) Current-
voltage relationships (I-V curves) of the Ca2 + current obtained from the cells without 1nM motilin treatment (open circles), with 1 nM motilin (filled circles), and after the wash-out of motilin (open squares). No significant changes in current amplitudes were detected among the treatments (n=15~16, p>0.05). (B) I-V curves showing the effects of 10 nM motilin on the Ca2 + current of the cells treated with the same ways. Open circles, control; filled circles, 10 nM motilin; open squares, wash-out of motilin. No significant changes in current amplitudes were detected among the treatments (n=14~15, p>0.05).
Fig. 9. Membrane depolarization by motilin and erythromycin.
(A) Motilin (10 nM)-induced depolarization. (B) A motilide, erythromycin (15 µM) depolarized resting membrane potential (MP) with firing spikes on the plateau of slow waves. In accordance to the excitatory changes in slow waves, the contractile forces of the muscle were increased. Verapamil (10 µM) inhibited both the changes in electrical activities and forces.
Upper and lower traces indicate a continual recording from the same strip. The dotted lines under the slow waves indicate the resting membrane potential.
9
The Korean Journal of Gastroenterology : Vol. 39. No. 1. 2002
력 및 그 정도에는 변화가 없었다. 이는 모틸린에 의한 수축 효과는 흥분성 평활근세포에 직접 작용한다는 것 을 시사하며 수용체결합 분석 등 다른 연구에서 위장관 평활근 세포막에 모틸린 수용체가 존재하며 모틸린이 평 활근세포에 직접 작용하여 근수축을 유발한다는 보고와 일치한다 .2 1- 2 3 그러나 생체실험에서 모틸린에 의한 수축이 아트로핀에 의해 차단된다는 결과17 ,1 8도 있어 논란이 있는 실정이다 .
위장관의 운동은 위장관 벽의 내인성 신경계에 의해 기본적인 리듬과 연동운동의 방향이 조절되고 있으며 위 장관을 생체로부터 분리해내더라도 기본적인 수축리듬 및 연동운동이 속도조정자(pacemaker)에 의해 평활근의 주기 적 수축 및 내인성 신경계에 의해 조절되고 있다고 알려 져 있다 .2 4 ,2 5 그러나 생체 내에서 위장관운동은 내재신경 계가 고위중추 및 자율신경계로부터 오는 신호를 수용하 고 통합함으로써 전체적인 운동성을 조절하고 있다.2 5 즉, 동일한 위장관 부위를 대상으로 한 생체실험과 시험관내 실험의 결과가 서로 다르다는 것은 모틸린이 외인성 신 경계(고위중추 및 자율신경계)에 작용하여 그 하부에 있 는 내인성 신경계를 조절한다고 생각할 수 있다 . 그러므 로 생체와 시험관 내에서 얻어진 결과들이 상반된 것이라 기보다는 상호 보완적이거나 혹은 둘 다 모두 일어날 수 있는 현상으로 해석할 수도 있을 것이다.
평활근의 수축이 일어나기 위해서는 평활근세포내 칼 슘농도의 증가가 선행되어야 하므로 모틸린이 어떠한 과 정을 통하여 근세포내 칼슘농도를 증가시켜 근수축을 유 도하는지 검증하였다. 본 연구에서 nifedipine 전처리 후 지속적인 위상성 수축 없이 일과성의 수축만 관찰되었고 또한 세포밖 칼슘의 제거로 근수축이 억제됨으로써 L-형 막전압의존성 칼슘통로 활성화를 통한 칼슘 유입이 모틸 린에 의한 지속적인 위상성 수축에 관여하고 일부의 수축 은 세포내 칼슘저장고의 칼슘이온을 동원함으로써 나타난 다는 것을 말해준다 . 그러나 patch clamp실험에서 모틸린 에 의한 칼슘통로의 증가가 없다는 것을 확인하여 모틸 린이 칼슘통로를 직접 항진시켜 그 효과를 나타내는 것 이 아니라고 판단하였다. 현재까지 생체연구에서 모틸린 (유도체)은 칼슘통로를 통한 전류를 증가시킨다는 혹은 감소시킨다는 보고가 있었으며 이런 효과는 동물의 종 및 위장관 부위 또는 세포내 환경을 어떻게 유지하는가에 따 라 매우 다양하게 나타날 수 있다는 것이다 .2 6 ,2 7
모틸린이 칼슘전류를 직접 항진시키지 않음에도 모틸 린에 의한 근수축이 L-형 칼슘통로 차단제에 의해 억제된 다는 사실은 모틸린이 평활근세포막의 전압을 탈분극시켜 서 간접적으로 칼슘통로를 활성화시켜 세포내 칼슘농도의 변화를 초래함으로써 근수축이 나타날 가능성이 제기되었
다 .2 6 본 실험에서 모틸린과 EM이 평활근세포를 탈분극시 켰다 . Yamada 등2 6은 모틸린 유도체인 GM611를 이용하여 유사한 결과를 보였고 또한 이들은 그 이유로 GM611이 비 선택성 양이온통로를 활성화시킴으로써 평활근의 탈분극을 유도한다고 하였으며 patch clamp를 이용하여 직접적으로 모틸린에 의한 내향성 양이온 전류가 발생함을 보였다 . 따라서 본 실험에서 기록된 모틸린에 의한 막전압의 탈분 극도 토끼의 십이지장 평활근에서 기록된 양이온전류와 유사한 이온전류의 발생을 통하여 나타났을 가능성이 있 다 . 사람의 위 평활근세포에서도 토끼의 십이지장에서와 같은 모틸린 유도성 내향전류가 발생한다면 막전압의 탈 분극을 유도할 것이며 그에 따라 간접적으로 막전압의존 성 칼슘통로의 활성이 증가할 것으로 생각된다 . 향후 이 에 대한 추가 실험이 필요할 것이며 이런 가정이 사실로 확인된다면 모틸린에 의한 사람 위 평활근의 수축이 nifedipine에 의해 억제되는 사실을 쉽게 설명할 수 있을 것 이다. 한편, 이들은 GM611이 세포내 칼슘이온 농도에 민감 하게 반응하여 활성화되는 칼슘의존성 칼륨(Ca2 +-activated K+)통로의 활성을 기록하고 모틸린이 이 통로를 일과성으 로 항진시킨다는 사실로부터 평활근세포내 칼슘저장고로 부터 모틸린에 의한 칼슘 유리를 주장하였다 . 본 실험에 서도 모틸린에 의한 세포내 칼슘 유리가 간접적으로 확인 되었다. 즉 세포 밖의 칼슘을 제거한 상태에서 고농도의 아세틸콜린을 전처리하여 세포내 칼슘저장고를 고갈시킨 후에는 모틸린에 의한 일과성 수축이 없었다 . 그러나 사 람 위 평활근을 이용한 본 실험에서는 모틸린에 의한 세 포내 칼슘 유리 과정이 phospholipase C inositotriphosphate (PLC-IP3)경로를 통하여 일어나는지를 정확히 검증하지 못 하였으며 향후 연구에서 그 사실이 밝혀질 것으로 생각한 다 .
본 실험에서 흥미로운 결과는 비교적 저농도(1 nM)의 모틸린이 존재하는 상황에서 아세틸콜린에 의한 근수축이 현저히 항진되며 주로 nifedipine에 의해 차단되는 위상성 수축만이 선택적으로 증가된다는 것이다 . 이는 생리적으 로 매우 중요한 현상으로 보인다 . 아세틸콜린은 부교감신 경계 및 위장관 내인성 신경계에서 유리되는 신경전달물 질로서 위장관의 강력한 수축 효과를 나타낸다 . 생리적으 로 부교감신경계는 항시 활성화되어 있으며 위장관의 기 본적인 연동운동 역시 내인성 신경으로부터 유리되는 아 세틸콜린에 의해 조절받고 있다 .2 4 ,2 5 즉, 생체 내에서 위장 관 평활근은 항상 아세틸콜린에 의한 흥분성 신호를 받고 있는 것이다 . 따라서 위장관 평활근에 대한 아세틸콜린의 흥분성 신호는 공복기에도 나타날 것이며, 위장관 점막의 장크롬친화세포에서 모틸린이 분비되면 그에 따라 혈중 모틸린 농도가 증가하는데 그 농도는 지극히 낮다고 보고 1 0
Shim, et al. Mechanisms of Motilin Action on Smooth Muscle of the Human Stomach
되었다.2 8 따라서 자연발생적인 혹은 외부에서 투여된 모 틸린 및 그 유도체에 의한 MMC가 아트로핀이나 미주신 경 절단에 의해 소실된다는 사실에서 저농도의 모틸린에 의한 아세틸콜린 수축 항진 효과는 생리적으로 의미가 있 다고 판단된다 . 다시 말해 생리적인 MMC의 발생은 저농 도의 모틸린에 의해 지속적으로 콜린성 흥분 자극을 받고 있는 위장관 평활근의 흥분도가 크게 증폭됨으로써 나타 날 가능성이 있다는 것이다. 최근의 한 연구2 9에서, 토끼의 십이지장 평활근세포에서 저농도(1 nM)의 모틸린이 무스 카린 수용체 자극에 의해 유도되는 내향성 양이온전류를 크게 항진시킨다고 하였다 . 무스카린 수용체 자극에 의해 내향성 양이온전류가 발생하면 평활근 세포막 전압의 탈 분극이 유도됨으로써 간접적으로 칼슘통로의 열림이 증가 되고 그에 따라 근수축이 유도되는데, 저농도의 모틸린이 아세틸콜린에 의한 양이온전류를 크게 증폭할 경우 더욱 큰 막전압의 탈분극이 유도될 것이므로 아세틸콜린에 의 한 근수축이 크게 증가할 것이다. 따라서 이들의 결과를 본 실험의 결과에 적용해 보면, 저농도의 모틸린이 어떠 한 기전에 의해 아세틸콜린에 의한 수축을 증가시키는가 를 추측해 볼 수 있다 . 향후 사람의 위 평활근에서 토끼 의 평활근에서와 같은 모틸린에 의한 양이온전류의 발생 과 무스카린성 양이온전류의 항진 현상이 나타날 것인지 는 추가 실험 및 증명이 필요할 것이다.
요 약
목 적 : 모틸린은 위장관호르몬으로서 십이지장과 공장 의 점막에 있는 장크롬친화세포에서 합성되어 공복시에 주기적으로 혈중으로 분비되며 위장관 수축기능(MMC)에 관여한다 . 현재까지 모틸린의 근수축기전이 생체실험에서 는 흥분성 신경을 자극함으로써 위장관 운동을 유도한다 는 보고와 시험관 내에서는 모틸린이 직접 평활근세포에 자극 효과를 일으킨다는 보고가 있어 논란이 있다 . 본 연 구는 사람의 위유문부 평활근을 이용하여 모틸린에 의한 근수축 및 그 기전에 대해 시험관내실험을 통하여 알아보 고자 하였다 . 대 상 및 방 법 : 수술장에서 위암 등으로 수 술받는 사람의 위 평활근조직을 얻어 모틸린에 의한 근수 축 및 수축 크기를 측정하고 아세틸콜린 , 아트로핀 , 복어 독 등의 약물과 patch clamp법과 세포막 전압측정법을 이용하여 모틸린에 의한 근수축기전을 관찰하였다 . 결 과 : 모틸린은 농도의존적으로(1-100 nM) 위 평활근의 수축 을 증가시켰고 그 수축력은(EC5 0: 5.63 nM) 아세틸콜린(EC5 0: 4.1 M)에 비해 훨씬 강력하였다. 모틸린에 의한 수축 및 그 정도는 신경 차단제인 복어독이나 무스카린성 수용체 차 단제인 아트로핀에 의해 변화가 없었다 . 모틸린에 의한
근수축은 L-형 칼슘통로 차단제인 nifedipine 처리나 세포 밖 칼슘 제거로 억제되었다 . 유의한 수축을 유발하지 않 는 저농도(1 nM) 모틸린을 전처리하면 아세틸콜린에 의한 평활근 수축력이 크게 항진되었는데, 특히 L-형 칼슘통로 에 의해 차단되는 위상성 수축을 선택적으로 항진시켰다 . L-형 칼슘통로를 통한 칼슘전류의 측정에서는 모틸린 1 nM (- 168.6 ± 14.4 pA vs. - 152.0 ±9 .3 pA)과 10 nM (- 125.2
± 9.0 pA vs. -116.7 ± 9.0 pA)을 투여하기 전과 투여 후 칼슘전류는 모두 유의한 차이가 없었다 . 세포막 전압과 근수축을 동시에 기록한 실험에서 모틸린(10 nM)은 막전 압을 현저하게 탈분극시켰고 EM (15 µM)에 의해 세포막 의 탈분극과 서파의 고원기 (plateau)에서 가시전압이 발 생되었고 동시에 근축의 증가가 나타났으나 이런 변화는 verapamil ( 10 µM)에 의해 효과적으로 억제되었다 . 결 론 : 이상의 결과는 모틸린이 사람 위유문부 평활근에 존재하 는 모틸린 수용체의 자극을 통해 직접 수축을 유발한다는 것을 시사하며 또한 간접적으로 칼슘통로를 통한 칼슘 유 입을 증가시켜 평활근세포막을 탈분극시킴으로써 수축반 응이 나타날 것으로 생각된다.
색 인 단 어 : 모틸린, 아트로핀, L형 칼슘통로, 탈분극
참 고 문 헌
1. Ito Z, Takeuchi S, Aizawa I, Takayanagi R. Effect of synthetic motilin on gastric motor activity in conscious dogs.
Am J Dig Dis 1977;22:813-819.
2. Itoh Z, Aizawa I, Honda R, Hiwatashi K, Couch EF.
Control of lower-esophageal-sphincter contractile activity by motilin in conscious dogs. Am J Dig Dis 1978;23:341-345.
3. Peeters TL, Vantrappen G, Janssens J. Fasting plasma motilin levels are related to the interdigestive motility complex. Gastroenterology 1980;79:716-719.
4. Jennewein HM, Hummelt H, Siewert R, Waldeck F. The motor-stimulating effect of natural motilin on the lower esophageal sphincter fundus, antrum and duodenum in dogs.
Digestion 1975;13:246-250.
5. Vantrappen G, Janssens J, Peeters TL, Bloom SR, Chris- tofides ND, Hellemans J. Motilin and the interdigestive migrat- ing motor complex in man. Dig Dis Sci 1979;24:497-500.
6. Holloway RH, Blank E, Takahashi I, Dodds WJ, Layman RD. Motilin: a mechanism incorporating the opossum lower esophageal sphincter into the migrating motor complex.
Gastroenterology 1985;89:507-515.
7. Inatomi N, Satoh H, Maki Y, Hashimoto N, Itoh Z, Omura S. An erythromycin derivative, EM-523, induces motilin-like gastrointestinal motility in dogs. J Pharmacol Exp Ther 1 1
대한소화기학회지 :제39권 제 1호, 2002
1989;25 1:707-712.
8. Hall KE, Greenberg GR, El-Sharkawy TY, Diamant NE.
Relationship between porcine motilin-induced migrating motor complex- like activity, vagal integrity, and endogenous motilin release in dogs. Gastroenterology 1984;87:76-85.
9. Mizumoto A, Sano I, Matsunaga Y, Yamamoto O, Itoh Z, Ohshima K. Mechanism of motilin-induced contractions in isolated perfused canine stomach. Gastroenterology 1993;105:
425-432.
10. Ohtawa M, Mizumoto A, Hayashi N, Yanagida K, Itoh Z, Omura S. Mechanism of gastroprokinetic effect of EM523, an erythromycin derivative, in dogs. Gastroenterology 1993;
104:1320-1327.
11. Strunz U, Domschke W, Mitznegg P, et al. Analysis of the motor effects of 13-norleucine motilin on the rabbit, guinea pig, rat, and human alimentary tract in vitro. Gastroenterology
1975;68:1485-1491.
12. Adachi H, Toda N, Hayashi S, et al. Mechanism of the excitatory action of motilin on isolated rabbit intestine.
Gastroenterology 1981;80:783-788.
13. Ludtke FE, Muller H, Golenhofen K. Direct effects of motilin on isolated smooth muscle from various regions of the human stomach. Pflugers Arch 1989;414:558-563.
14. Kitazawa T, Ishii A, Taniyama K. The Leu 13-motilin (KW- 5139)-evoked release of acetylcholine from enteric neurones in the rabbit duodenum. Br J Pharmacol 1993;109:94-99.
15. Poitras P, Dumont A, Cuber JC, Trudel L. Cholinergic regulation of motilin release from isolated canine intestinal cells. Peptides 1993;14:207-213.
16. Kitazawa T, Taneike T, Ohga A. Excitatory action of [Leu 13] motilin on the gastrointestinal smooth muscle isolated from the chicken. Peptides 1995;16:1243-1252.
17. Kitazawa T, Ichikawa S, Yokoyama T, Ishii A, Shuto K.
Stimulating action of KW-5139 (Leu13-motilin) on gastrointes- tinal motility in the rabbit. Br J Pharmacol 1994;111:288-294.
18. Boivin M, Pinelo LR, St-Pierre S, Poitras P. Neural mediation of the motilin motor effect on the human antrum.
Am J Physiol 1997;272:G71-G76.
19. Van Assche G, Depoortere I, Thijs T, Janssens JJ, Peeters TL. Concentration-dependent stimulation of cholinergic motor nerves or smooth muscle by [Nle13]motilin in the isolated rabbit gastric antrum. Eur J Pharmacol 1997;337:267-274.
20. Hamill OP, Marty A, Neher E, Sakmann B, Sigworth FJ.
Improved patch-clamp techniques for high-resolution current recording from cells and cell-free membrane patches.
Pflugers Arch 1981;391:85-100.
21. Bormans V, Peeters TL, Vantrappen G. Motilin receptors in rabbit stomach and small intestine. Regul Pept 1986;15:
143-153.
22. Peeters TL, Bormans V, Vantrappen G. Comparison of motilin binding to crude homogenates of human and canine gastrointestinal smooth muscle tissue. Regul Pept 1988;23:171- 182.
23. Depoortere I, Peeters TL, Vantrappen G. Distribution and characterization of motilin receptors in the cat. Peptides 1993;14:1153-1157.
24. Horowitz B, Ward SM, Sanders KM. Cellular and molecular basis for electrical rhythmicity in gastrointestinal muscles.
Annu Rev Physiol 1999;61:19-43.
25. Kunze WA, Furness JB. The enteric nervous system and regulation of intestinal motility. Annu Rev Physiol 1999;61:
117-142.
26. Yamada K, Chen S, Abdullah NA, Tanaka M, Ito Y, Inoue R. Electrophysiological characterization of a motilin agonist, GM611, on rabbit duodenal smooth muscle. Am J Physiol 1996;271:G1003-G1016.
27. Farrugia G, Macielag MJ, Peeters TL, Sarr MG, Galdes A, Szurszewski JH. Motilin and OHM-11526 activate a calcium current in human and canine jejunal circular smooth muscle.
Am J Physiol 1997;273:G404-G412.
28. Bloom SR, Mitznegg P, Bryant MG. Measurement of human plasma motilin. Scand J Gastroenterol Suppl 1976;39:47-52.
29. Yamada K, Yanagida H, Ito Y, Inoue R. Postsynaptic enhancement by motilin of muscarinic receptor cation currents in duodenal smooth muscle. Am J Physiol 1998;
274:G487-G492.
1 2