저작자표시-비영리-변경금지 2.0 대한민국 이용자는 아래의 조건을 따르는 경우에 한하여 자유롭게 l 이 저작물을 복제, 배포, 전송, 전시, 공연 및 방송할 수 있습니다. 다음과 같은 조건을 따라야 합니다: l 귀하는, 이 저작물의 재이용이나 배포의 경우, 이 저작물에 적용된 이용허락조건 을 명확하게 나타내어야 합니다. l 저작권자로부터 별도의 허가를 받으면 이러한 조건들은 적용되지 않습니다. 저작권법에 따른 이용자의 권리는 위의 내용에 의하여 영향을 받지 않습니다. 이것은 이용허락규약(Legal Code)을 이해하기 쉽게 요약한 것입니다. Disclaimer 저작자표시. 귀하는 원저작자를 표시하여야 합니다. 비영리. 귀하는 이 저작물을 영리 목적으로 이용할 수 없습니다. 변경금지. 귀하는 이 저작물을 개작, 변형 또는 가공할 수 없습니다.
A THESIS
FOR THE DEGREE OF MASTER OF SCIENCE
Juvenile hormone antagonistic activity of actinobacteria and their
insecticidal activity against Plutella xylostella
방선균의 곤충 유약호르몬 길항제 탐색 및
배추좀나방 살충활성 검정
By
Jun Young Kim
Major in Entomology
Department of Agricultural Biotechnology
Seoul National University
Juvenile hormone antagonistic activity of actinobacteria and their
insecticidal activity against Plutella xylostella
방선균의 곤충 유약호르몬 길항제 탐색 및
배추좀나방 살충활성 검정
UNDER THE DIRECTION OF ADVISER YEON HO JE
SUBMITTED TO THE FACULTY OF THE GRADUATE SCHOOL OF SEOUL NATIONAL UNIVERSITY
By
Jun Young Kim
Major in Entomology
Department of Agricultural Biotechnology
Seoul National University
February, 2020
APPROVED AS A QUALIFIED THESIS OF Jun Young Kim
FOR THE DEGREE OF MASTER OF SCIENCE
BY THE COMMITTEE MEMBERS
CHAIRMAN Joon-Ho Lee _________________________
VICE CHAIRMAN Yeon Ho Je _________________________
MEMBER Jun-Hyung Tak _________________________
II
Juvenile hormone antagonistic activity of actinobacteria and their
insecticidal activity against Plutella xylostella
Major in Entomology
Department of Agricultural Biotechnology
Seoul National University
February 2020
Jun Young Kim
ABSTRACT
Insect growth regulators (IGRs) are promising alternatives to conventional chemical insecticides because they are specific to target insects and low toxic to non-target organisms. Actinobacteria produce a wide range of secondary metabolites with insecticidal and insect growth regulatory activities, and therefore, these bacteria could be potential sources of novel IGR compounds. In this study, to identify novel IGR compounds from actinobacteria, culture media of 1,904 actinobacteria isolated from Korean soil samples were investigated for their juvenile hormone (JH)-based IGR activities. Whereas there were no actinobacteria
III
isolates with JH agonist (JHA) activity, 25 isolates showed high JH antagonist (JHAN) activity. Among them, 4 isolates exhibited high level of insecticidal activity against 3rd instar larvae of Plutella xylostella. Two isolates IMBL-1412 and IMBL-1823 showing relatively higher insecticidal activities were identified as Streptomyces lactacystinicus based on culture characteristics on various ISP media and nucleotide sequence of 16S rRNA gene. Although the two isolates showed 100% of similarity in 16S rRNA nucleotide sequence, nucleotide sequence of another marker, recA gene, was different each other, demonstrating that these two isolates could belong to different subspecies. When the culture media of these two isolates were extracted sequentially using hexane, ethyl acetate and butyl alcohol, ethyl acetate fractions of both isolates showed high JHAN and insecticidal activities against P. xylostella larvae at a concentration of 100 ppm. These results suggested that secondary metabolites of actinobacteria could be efficiently applied as novel IGR insecticides.
Key words: Actinobacteria, Streptomyces lactacystinicus, insect growth regulator, Juvenile
hormone antagonist, Plutella xylostella
IV
TABLE OF CONTENTS
ABSTRACT ... II
TABLE OF CONTENTS ... IV
LIST OF TABLES ... VI
LIST OF FIGURES ... VII
INTRODUCTION ... 9
LITERATURE REVIEW ... 12
1.
Actinobacteria ... 12
2.
Juvenile hormone ... 14
3.
Insect growth regulator (IGR) ... 16
METERIAL AND METHODS ... 20
1.
Insects ... 20
2.
Actinobacteria ... 20
3.
Yeast two-hybrid β-galactosidase assay ... 21
4.
Yeast growth inhibition tests ... 22
5.
Insect bioassay ... 22
V
7.
Isolation of JHAN compounds from the selected actinobacteria ... 23
RESULTS ... 26
1.
Screening of actinobacteria for IGR activity
...26
2.
Insecticidal activity of actinobacteria with JHAN activity ... 29
3.
Concentraiton-dependent activities of the selected actinobacteria
strains ... 35
4.
Taxonomic identification of the selected actinobacteria strains .... 36
5.
Activities of the selected actinobacteria culture extracts ... 43
DISCUSSION ... 48
LITERATURES CITED ... 50
VI
LIST OF TABLES
VII
LIST OF FIGURES
Figure 1. Schematic diagram for extraction of actinobacteria culture media. ... 25
Figure 2. Screening of actinobacteria culture media for their IGR activites. ... 27
Figure 3. JHAN activity of actinobacteria strains. ... 28
Figure 4. Insecticidal activity of actinobacteria strains against P. xylostella. ... 30
Figure 5. Assay method-dependent insecticidal activities of actinobacteria strains against P. xylostella. ... 31
Figure 6. Time-course insecticidal activities of actinobacteria strains against P. xylostella. ... 32
Figure 7. Insecticidal activity of actinobacteria strains against A. albopictus. ... 33
Figure 8. Insecticidal activity of actinobacteria strains against B. tabaci. ... 34
Figure 9. Concentration-dependent JHAN activities of actinobacteria strains. ... 36
Figure 10. Anti-yeast activity tests of actinobacteria strains with JHAN activities. ... 37
Figure 11. Concentration-dependent insecticidal activities of actinobacteria strains against P. xylostella. ... 38
Figure 12. Phylogenetic relationship of the IMBL-1412 and IMBL-1823 strains based on their nucleotide sequence of 16S rRNA gene. ... 41
Figure 13. Phylogenetic relationship of the IMBL-1412 and IMBL-1823 strains based on their nucleotide sequence of recA gene. ... 42
Figure 14. Concentration-dependent JHAN activities of the strain IMBL-1412 culture extracts. ... 44
VIII
Figure 15. Concentration-dependent JHAN activities of the strain IMBL-1823 culture extracts. ... 45 Figure 16. Concentration-dependent insecticidal activity of the strain IMBL-1412 culture extracts against P. xylostella. ... 46 Figure 17. Concentration-dependent insecticidal activity of the strain IMBL-1823 culture extracts against P. xylostella. ... 47
9
INTRODUCTION
Insects not only cause severe economic damage to agricultural products but also act as vectors of human diseases by carrying various pathogens (Boyer, Zhang, and Lempérière 2012; Hill et al. 2005). Chemical insecticides have been commonly used to minimize the losses to agriculture and to protect human health. However, due to their toxicity to the environment and the development of insect resistance, the demands for environmentally benign insecticides are on the rise.
Actinobacteria are filamentous gram-positive bacteria with a high G+C content and widely distributed in various terrestrial and aquatic environments (Sharma, Dangi, and Choudhary 2014). Actinobacteria represent one of the most studied classes of bacteria because they produce a broad spectrum of biologically active compounds (Mahajan and Balachandran 2014; Solecka et al. 2012). Approximately 10,000 bioactive secondary metabolites are reported to be produced by actinobacteria, which represents 45% of all bioactive microbial metabolites discovered (Jackson et al. 2018).
Actinobacteria have also been identified as potential biological pest control agents (Omura 2011; Montesinos 2003). It has been reported that secondary metabolites derived from actinobacteria show insecticidal activities against a variety of insect pests belonging to Lepidoptera and Diptera (Vijayabharathi et al. 2014; Ababutain, Abdul Aziz, and AL-Meshhen 2012; Karthik et al. 2011; El-Khawagh, Hamadah, and El-Sheikh 2011; Dhanasekaran et al. 2010). Furthermore, recent studies have shown that actinobacteria produce secondary metabolites with insect growth regulatory activities (Samri et al. 2017; Kaur et al. 2014; Arasu et al. 2013).
10
Insect growth regulators (IGRs) are promising alternatives to conventional chemical insecticides because of their selectivity and relatively low toxicity (Pener and Dhadialla 2012; Dhadialla, Retnakaran, and Smagghe 2009). Depending on their mode of action, commercially available IGRs have been divided into three classes: juvenile hormone (JH) agonists, ecdysone agonists and chitin synthesis inhibitors (Dhadialla, Retnakaran, and Smagghe 2009). As JH is insect specific hormone and regulates development, reproduction, diapause and many other aspects of insect physiology, JH-based IGRs, including JH agonists (JHAs) and antagonists (JHANs), fatally disrupt the endocrine system, which causes abnormal development and larval death (S.-H. Lee et al. 2015; Pener and Dhadialla 2012).
It has been reported that methoprene-tolerant (Met) functions as the JH receptor (Jindra, Palli, and Riddiford 2013; Charles et al. 2011). Met is a member of a protein family known as basic-helix-loop-helix (bHLH)-Per-Arnt-Sim (PAS) transcription factors, which have significant roles in JH activity and downstream transcriptional activation. This family of transcription factors must dimerize to regulate transcription (Kewley, Whitelaw, and Chapman-Smith 2004). Met of Aedes aegypti dimerizes with other bHLH-PAS transcription factors, such as Ftz-F1-interacting steroid receptor coactivator (FISC) or Cycle (CYC), in a JH-dependent manner (Shin et al. 2012; M. Li, Mead, and Zhu 2011). Previously, this JH-mediated interaction of Met and its binding partners have been replicated in vitro using yeast cells transformed with the Met and FISC/CYC genes of A.
aegypti (S.-H. Lee et al. 2015). Using this in vitro yeast two-hybrid β-galactosidase ligand
binding assay, various compounds with JHA or JHAN activity have been identified from plant essential oils and chemical libraries (S.-H. Lee, Lim, et al. 2018; S.-H. Lee, Ha, et al.
11 2018; S.H. Lee, Choi, et al. 2018).
In this study, it was assumed that actinobacteria could be another source of natural JHAs and JHANs because they produce a variety of secondary metabolites with insecticidal and insect growth regulatory activities. In order to explore novel IGR compounds from actinobacteria, culture media of 1,904 actinobacteria isolates were screened for their JHA/JHAN activities, and insecticidal activities of culture media with high JHAN activity were evaluated.
12
LITERATURE REVIEW
1. Actinobacteria
Actinobacteria are Gram-positive filamentous bacteria with a high guanine-plus-cytosine (GC) content in their genomes. They grow by a combination of tip extension and branching of the hyphae. This is what gave them their name, which derives from the Greek words for ray (aktis or aktin) and fungi (mukes). Traditionally, actinobacteria were considered transitional forms between fungi and bacteria. Indeed, like filamentous fungi, many Actinobacteria produce a mycelium, and many of these mycelial actinobacteria reproduce by sporulation. However, the comparison to fungi is only superficial: like all bacteria, actinobacteria’ cells are thin with a chromosome that is organized in a prokaryotic nucleoid and a peptidoglycan cell wall; furthermore, the cells are susceptible to antibacterial agents. Physiologically and ecologically, most Actinobacteria are aerobic, but there are exceptions. Further, they can be heterotrophic or chemoautotrophic, but most are chemoheterotrophic and able to use a wide variety of nutritional sources, including various complex polysaccharides (Zimmermann 1990; Lechevalier and Lechevalier 1965). Actinobacteria may be inhabitants of soil or aquatic environments (e.g., Streptomyces, Micromonospora, Rhodococcus, and Salinispora species), plant symbionts (e.g., Frankia spp.), plant or animal pathogens (e.g., Corynebacterium, Mycobacterium, or Nocardia species), or gastrointestinal commensals (e.g., Bifidobacterium spp.).
Actinobacteria are of great importance in the field of biotechnology, as producers of a plethora of bioactive secondary metabolites with extensive industrial, medical, and
13
agricultural applications. In particular, Actinobacteria produce the majority of the naturally occurring antibiotics. The first antibiotics discovered in Actinobacteria were actinomycin from a culture of Streptomyces antibioticus in 1940 (Waksman and Woodruff 1940), streptothricin from Streptomyces lavendulaein 1942 (Waksman and Woodruff 1942), and streptomycin from Streptomyces griseus in 1944 (Schatz and Waksman 1944), all of which were discovered by Waksman and colleagues. Streptomycetes have been the major source of clinical antibiotics and are responsible for over 80% of all antibiotics of actinobacterial origin (Ilic et al. 2007). That actinomycin, streptomycin, and streptothricin were the first to be found is not surprising, as these molecules occur at much higher frequencies than many other antibiotics. The producing capacity of individual actinobacteria can also vary enormously. Some Streptomyces species produce a single antibiotic, while others produce a range of different compounds and compound classes. Besides antibiotics, Actinobacteria also produce a wide variety of other secondary metabolites with activity as herbicides (Tindall et al. 2006), antifungals, antitumor or immunosuppressant drugs, and anthelmintic agents (Běhal 2000). Examples are given below.
Macrotetrolides are active against mites, insects (Jizba et al. 1991; Sagawa et al. 1972; Oishi et al. 1970), coccidia (Sakamoto et al. 1978), and helminths, and they also show immunosuppressive effects (Shichi, Tanouchi, and Kamada 1989). They are produced by a variety of Streptomyces species. However, with regard to the composition of the macrotetrolide complex, only S. aureus S-3466 (Ando et al. 1971), which produces a mixture of tetranactin (the most active member of the compound group) with dinactin and trinactin (Ando et al. 1971; Oishi et al. 1970), has been utilized for commercial purposes. Tetranactin, a cyclic antibiotic produced by Streptomyces aureus with a molecular structure
14
related to cyclosporine, is used as emulsion against carmine mites of fruits and tea. A true success story in terms of anthelmintics is ivermectin (Ōmura and Crump 2014), which is a dehydro derivative of avermectin produced by Streptomyces avermitilis. After its appearance in the late 1970s, ivermectin was the world’s first endectocide, which at the time was a completely novel class of antiparasitic agents, with strong and broad-spectrum activity against both internal and external nematodes and arthropods. Recently, the Nobel Prize for Physiology or Medicine 2015 was awarded to Satoshi Omura and William C. Campbell for their discovery of avermectin, jointly with Youyou Tu for the discovery of the antimalarial drug artemisinin.
2. Juvenile hormone
It has been almost two centuries since studies relating juvenile hormone (JH) were started. Müller described specific organs in the cockroach which were renamed as the corpora allata (CA) in 1899. However, until then, the CA was described as sympathetic ganglia concerned with the innervation of the digestive system. Although Police (1910) suggested that the CA is endocrine organs concerned with nervous function, it had remained to be proved that. In 1934, Wigglesworth (1934) began historical studies on insect JH, making efficient use of surgical techniques. He assumed at first that the CA is the source of the molting hormone, an “inhibitory factor” which prevents the first four larval stages from molting directly into adults in Rhodnius. In 1936, he showed that the CA is the source of the inhibitory hormone that prevents metamorphosis in young larvae and that the CA from young larvae when implanted into fifth instars caused them to undergo a
15
supernumerary molt (V.B. Wigglesworth 1936). Wigglesworth concluded that the concentration of the inhibitory hormone from CA determines the extent of metamorphosis at the next molt. Then, there have been many studies on JH function. The modern era of JH research began with the critical finding by Carroll Williams (1956) that he discovered a natural repository for juvenile hormone, “golden oil”, and JH was extracted and diluted in peanut oil or mineral oil to conduct hundreds or even thousands of experiments from male
Hyalophora cecropia. About 10 years after, the structure of JH was identified using gas
chromatographic analysis. Röller and colleague (Röller et al. 1967) identified the first juvenile hormone from lipid extracts of the wild silk moth, H. cecropia. This JH, methyl (2E, 6E, 10-cis) -10,11-epoxy-7-ethyl-3,11-dimethyl-2,6-tridecadienoate, was termed cecropia JH or C18 JH in older literature, but it is now recognized as JH I. Meyer (Meyer et al. 1968) identified a minor component that is called JH II, which differed from JH I by a methyl group at C7 in the H. cecropia extracts. A third JH homologue, JH III, methyl 10, 11–epoxy–farnesoate, was identified from media in which the CA of the tobacco hornworm,
Manduca sexta, had been contained (Judy et al. 1973). JH III differs from the other
homologues in that all three branches of the carbon skeleton, at C3, C7, and C11, are methyl groups. JH III appears to be the most common homologue among the species studied (Schooley et al. 1984). The trihomosesquiterpenoids JH 0 and its isomer 4–methyl JH I (iso–JH 0) were identified in M. sexta eggs (Bergot, Schooley, and De Kort 1981), but nothing is currently known of their functions. JH III bisepoxy (JHB3) was identified from
in vitro cultures of larval ring glands of Drosophila melanogaster (Richard, Applebaum,
and Gilbert 1989). These historical studies formed the basis of many JH-related studies today. JHs are a group of acyclic sesquiterpenoids that secreted from endocrine glands
16
called CA. Main role of JHs were first recognized and described by Wigglesworth in the blood sucking bug, Rhodnius prolixus (V.B. Wigglesworth 1934). JHs roles mainly in various physiological functions including molting, metamorphosis, reproduction, polyphenism, caste differentiation, and various physiological functions in insects (Hartfelder and Emlen 2012; Raikhel, Brown, and Belles 2005; Nijhout 1998; Riddiford 1994). Although JHs are very important for insect physiology, their regulatory mechanisms have remained elusive (Riddiford 2008).
3. Insect growth regulator (IGR)
Carol Williams (1967) suggested the use of insects own hormone to pest control, and he termed as “third-generation insecticides”. Schneiderman (1972) used the term insect growth regulators (IGRs) that regulate insect growth and development. Now IGRs are termed as chemicals that interfere with insect specific development, normal growth and reproduction. The first IGRs used for pest control were JH mimics or JH agonist. And chitin synthesis inhibitors and ecdysteroid agonists have been added later.
These insecticides possess relatively low environmental toxicity, such as low toxicity to off-target like man, wildlife, and environment. Furthermore, IGRs have high specificity cause effect against only targeted specific taxa (Pener and Dhadialla 2012).
Juvenile hormone agonists (JHAs)
JH regulates molting, metamorphosis and reproduction in insects. Due to its importance JH has long been considered as novel pesticides (Cusson, Sen, and Shinoda 2013). The first JH active compounds were sesquiterpenoid farnesol and farnesal (V. Wigglesworth 1961).
17
Later these compounds were announced as JH precursors and chemical structures of JHs were elucidated. But chemical properties of natural JHs are unstable and have vulnerable sites for degradation caused by lights, water, and temperature to use them into pest management (Sláma 1999; Judy et al. 1973; Meyer et al. 1968; Röller et al. 1967).
The first botanical JH agonist “The paper factor” containing Canadian balsam fir was first identified by Slama and Willians (Sláma and Williams 1966b). Pyrrhocoris apterus reared on the paper towels made from Canadian balsam fir cause abnormal development like metamorphosis failure and became nymphal-adult intermediate creatures or extra instar nymphs. Also eggs from adults that normally developed showed reduced hatch rate. Eventually, it was discovered that the balsam fir containing juvabione acts as a JHA (Slama 1971; Sláma and Williams 1966a).
After the discovery of juvabione, numerous plant derived sesquiterpenoids were screened for their JHA activities (W.S. Bowers and Bodenstein 1971). Despite extensive endeavors, however, only few JHAs have been identified as of now (W.S. Bowers 2012). But large number of synthetic sesquiterpenoid JHA was revealed and Zoecon Corporation registered hydroprene and methoprene (isopropyl 11-methoxy 3,7,11 trimethyldodeca-2,4-dienoate), which became first JHA commercialized IGR insecticide (HENRICK 1982). They have been successfully used against mosquitoes, ants and flies and they are still favored as the least toxic, environmentally safe insecticides.
In 1981, Hoffmann-LaRoche laboratories reported that juvenoid containing phenoxyphenyl group shows high JH activity. The most active molecule in 4-phenoxyphenyl series was fenoxycarb (Masner et al. 1981). As the one of the most successful JHA, the pyriproxyfen has been commercialized in 1986. It is also fenoxycarb
18
derivatives in which side chain has been replaced by pyridyl structure (HATAKOSHI, AGUI, and NAKAYAMA 1986).
Juvenile hormone antagonists (JHANs)
Since JHA discovered, inspired thoughts that the reverse principle, anti-juvenile hormone agent could be explored to complement the use of JHA (Staal 1986). And it could offer more attractive method of control because accelerate metamorphosis would shorten the larval lifetime (Quistad et al. 1981).
Fluoromevalonate (FMev), tetrahydro-4-fluoromethyl-4-hydroxy-2H-pyran-2-one, was previously known for its hypocholesteremic activity in mammalian systems, showed anti JH activity in Lepidoptera. FMev induced precocious metamorphosis in several Lepidoptera larvae. And later, it was discovered that FMev acts as a reversible inhibitor in JH biosynthesis (Quistad et al. 1981). Imidazole caused precocious metamorphosis in
Bombyx mori. Later, substituted imidazoles act as methyl farnesoate inhibitor in JH
synthesis (Unnithan et al. 1995; ASANO, KUWANO, and ETO 1986).
Bowers discovered prococene 1 and prococene 2 that showed anti JH activity in the extract of Ageratum houstonianum (W.S. Bowers 1977; W. Bowers 1976). These compounds induce precocious metamorphosis, inhibition of vitellogenic development in oocytes. These compounds were shown allactocidal activity by forming highly reactive epoxides in the CA (Hamnett et al. 1981; W.S. Bowers 1981; Barovsky and Brooker 1980; W.S. Bowers 1977).
Recent studies have identified JHANs from plants Lindera erythrocarpa and Solidago
19
and insecticidal activity to Aedes aegypti larvae were characterized. Also, topical application of these compounds caused a retardation of follicle development in female mosquito ovaries. The discovery of JHANs, along with plant derived JHAs like juvabione, indicates that plants produce IGRs, and that they use these substances as a part of their defense system against herbivores (S.-H. Lee et al. 2015).
20
METERIAL AND METHODS
1. Insects
The diamondback moth Plutella xylostella (L.) (Lepidoptera: Plutellidae) was reared on rape sprouts and maintained at 25°C and 70% relative humidity with a 16 h light/8 h dark cycles. Aedes albopictus (Skuse) (Diptera: Culicidae) was maintained in breeding chambers at 28°C and 70% relative humidity with a 12 h light/12 h dark cycles in aged tap water. Larvae were fed on a diet of TetraMin fish flakes, and adults were reared using 10% sucrose solution. Bemisia tabaci was reared on solanum melongena in plastic cages under a 16 h light /8 h dark cycles at 25°C and 70% relative humidity.
2. Actinobacteria
A total of 1,904 actinobacteria were isolated from soil samples collected from various sites in Korea. One gram of soil samples was serially diluted with distilled water and homogenized by vortexing for 10 min. An aliquot of the soil suspension was plated on humic acid-vitamin agar (1 g humic acid, 0.5 g Na2HPO4, 1.71 g KCl, 0.05 g MgSO4·7H2O,
0.01 g FeSO4·7H2O, 1 g CaCl2, B vitamins (0.5 mg each of thiamine-HCl, riboflavin,
niacin, pyridoxine, Ca-pantothenate, inositol and p-aminobenzoic acid, and 0.25 mg of biotin), 15 g agar and 1 L water, pH 7.2) and incubated at 28°C for 7 days. Selected isolates were maintained as a spore suspension in glycerol (20%, v/v) at -80°C. All 1,904 isolates were fermented in a 500 ml baffled Erlenmeyer flask containing 100 ml of M3 culture media (1 g soytone, 1 g glucose, 2 g soluble starch, 0.3 g CaCO3, 0.02 g FeSO4· 7H2O),
21
which were incubated on a rotary shaker (150 rpm) at 30°C for 7 days. After fermentation, the culture media were harvested by centrifugation at 10,000 rpm for 15 min at 4°C.
3. Yeast two-hybrid β-galactosidase
a ssayThe Y-187 yeast cells transformed with A. aegypti Met-FISC were incubated at 30°C in DDO (SD -Leu/-Trp) media until OD600 values reached 0.3-0.4. After harvest, the cells
were suspended in the fresh media at a concentration of 2.0×106 cells / ml and 100 μl of
the cells was distributed in 96-well plates. To estimate JHA activity, 1 μl of each actinobacteria culture media was added into each well, and the cells were incubated for 3 h and subjected to the β-galactosidase assays using the yeast β-galactosidase assay kit (Thermo Scientific, USA). A positive control treated with 0.033 ppm of pyriproxyfen and a negative control treated with 1 μl of M3 culture media was placed in each tested plate. The assay reaction mixtures in the 96-well plates were incubated at 30°C for 5 h, and the OD420 was measured using an iMarkTM microplate reader (BIO-RAD, USA). The obtained
OD420 values were converted to an arbitrary unit representing JHA activity.
JHA activity = OD420 of sample
OD420 of pyriproxyfen (0.033 ppm)
For JHAN activity, 100 μl of yeast cells (2.0×106 cells / ml) distributed in 96-well plates
was treated with 0.033 ppm of pyriproxyfen and 1 μl of each actinobacteria culture media. A negative control treated with 0.033 ppm of pyriproxyfen and 1 μl of M3 culture media was placed in each tested plate. The cells were incubated for a further 3 h and subjected to the β-galactosidase assays as described above. The obtained OD420 values were converted
22
JHAN activity = OD420 of pyriproxyfen (0.033 ppm) - OD420 of treated sample OD420 of pyriproxyfen (0.033 ppm)
4. Yeast growth inhibition tests
The transformed Y187 yeast cells with A. aegypti Met-FISC were incubated at 30°C in DDO (SD -Leu/-Trp) media until OD600 values reached 0.3-0.4. After harvesting, the cells
were suspended in the fresh media at a concentration of 2.0×106 cells / ml, and 200 μl of
the cells was treated with 2 μl of each actinobacteria culture media in 96 well plates. The treated cells were incubated at 30°C with shaking, and the OD600 of each sample was
measured every 3 h for 1 day.
5. Insect bioassay
Insecticidal activities of actinobacteria culture media against P. xylostella larvae were evaluated using two separate assay methods, Cabbage leaf dipping and larval immersion. For Cabbage leaf dipping assay, thirty 3rd instars were fed on Chinese cabbage leaf disc (60 mm diameter) soaked in serially diluted culture media or various concentrations (1, 10, 50, and 100 μg/ml) of culture extracts. In case of larval immersion, thirty 3rd instars were soaked in undiluted culture media for 30 sec and the treated larvae were provided Chinese cabbage leaf discs (60 cm diameter). Thirty A. albopictus 3rd instars were treated with actinobacteria culture media by adding 2.5 m1 of culture media in 2.5 ml tap water with food mixtures. Thirty adults of B. tabaci were fed on Solanum melongena leaf disc (15 mm diameter) treated with undiluted culture media. The number of dead larvae was checked at 24 h intervals and calculated after treatment for 5 days. All experiments were performed in
23 triplicate.
6. Taxanomic characteristics of the selected actinobacteria
The cultural characteristics of the strains IMBL-1412 and IMBL-1823 were determined on various International Streptomyces Project (ISP) media. The colony color was evaluated using the Inter-Society Color Council-National Bureau of Standards (ISCC-NBS) color chart (Kelly and Judd 1965). PCR amplification and nucleotide sequence analysis of the 16S rRNA and recA genes of the strains was performed as previously reported (Meyers 2015; W.-J. Li et al. 2007). The 16S rRNA sequences from closely related actinobacteria were retrieved from the National Center for Biotechnology Information (http://www.ncbi.nlm.nih.gov/GenBank/index.html) and aligned using the CLUSTAL X program (Thompson et al. 1997). Molecular phylogeny of the selected strains for 16S rRNA and recA, respectively, was inferred using the neighbor-joining method under the Jones-Taylor-Thornton (JTT) matrix-based model incorporated in the MEGA7 software (Jones, Taylor, and Thornton 1992).
7. Isolation of JHAN compounds from the selected actinobacteria
To separate JHAN compounds from the selected strains, the culture media of IMBL-1412 and IMBL-1823 was sequentially extracted with equivalent volumes of n-hexane, ethyl acetate and n-butanol (Fig.1). These extracts were concentrated in a rotary vacuum and redissolved in dimethyl sulfoxide (DMSO) for activity tests. The JHAN and
24
25
26
RESULTS
1. Screening of actinobacteria for IGR activites
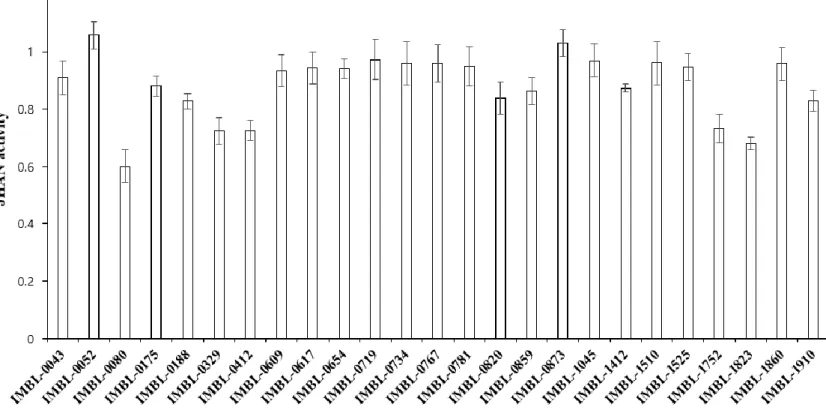
To isolate novel compounds with JHA or JHAN activity, culture media of 1,904 actinobacteria strains were tested using in vitro yeast two-hybrid β-galactosidase assay (Fig. 2). Among 1,904 actinobacteria strains, there were no culture media showing JHA activity. In contrast, culture media of 25 actinobacteria strains highly interfered with the binding of
A. aegypti Met-FISC, suggesting that these actinobacteria strains produce secondary
27
28
Figure 3. JHAN activity of actinobacteria strains. To estimate JHAN activity, 0.033 ppm of pyriproxyfen and 1 μl of each undiluted culture media were applied to yeast two-hybrid β-galactosidase assay.
29
2. Insecticidal activities of actinobacteria strains with JHAN activity
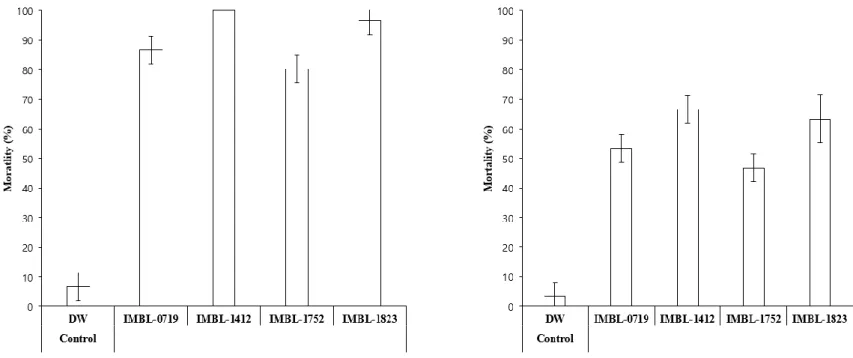
To evaluate insecticidal activities of actinobacteria with JHAN activity, 3rd instar larvae of P. xylostella and A. albopictus and adult of B. tabaci were treated with culture media of each actinobacteria strains, respectively. Among the 25 actinobacteria strains, culture media from strains IMBL-0719, IMBL-1412, IMBL-1752 and IMBL-1823 showed high levels of insecticidal activities against P. xylostella 3rd instar larvae, with mortalities greater than 80% (Fig. 4).
To investigate insecticidal activities of these 4 strains further, culture media of them were applied to 3rd instar larvae of P. xylostella using cabbage leaf dipping and larval immersion method, respectively. In all 4 strains tested, larval mortalities from cabbage leaf dipping assay were higher than those from larval immersion assay, demonstrating that JHAN compounds from these actinobacteria strains have both oral and topical toxicities (Fig. 5). Also, mortalities of P. xylostella larvae treated with these culture media were gradually increased with elapsed time (Fig. 6).
The insecticidal spectrum of actinobacteria was further investigated against A. albopictus larvae and B. tabaci adults (Fig. 7 and 8). All 4 culture media of actinobacteria showing high insecticidal activities against P. xylostella larvae exhibited very low insecticidal activities against both A. albopictus and B. tabaci, with mortalities smaller than 20%, demonstrating that JHAN compounds from these actinobacteria strains are highly specific to P. xylostella larvae.
30
Figure 4. Insecticidal activity of actinobacteria strains against P. xylostella. Third instar larvae of P. xylostella were treated with undiluted culture media of each actinobacteria strain and the mortality was calculated at 5 days after treatment.
31
Figure 5. Assay method-dependent insecticidal activities of actinobacteria strains against P. xylostella. Third instar larvae of P.
xylostella were treated with undiluted culture media of each actinobacteria strain using cabbage leaf dipping method (Left) and
32
Figure 6. Time-course insecticidal activities of actinobacteria strains against P. xylostella. Third instar larvae of P. xylostella were treated with undiluted culture media of each actinobacteria strain using cabbage leaf dipping method (Left) and larval immersion method (Right), and the mortality was calculated every 24 h for 5 days after treatment.
33
Figure 7. Insecticidal activity of actinobacteria strains against A. albopictus. Third instar larvae of A. albopictus were treated with 50% of each actinobacteria culture media and the mortality was calculated at 5 days after treatment.
34
Figure 8. Insecticidal activity of actinobacteria strains against B. tabaci. B. tabaci adults were treated with undiluted culture media of each actinobacteria strain and the mortality was calculated at 5 days after treatment.
35
3. Concentration-dependent activities of the selected actinobacteria strains
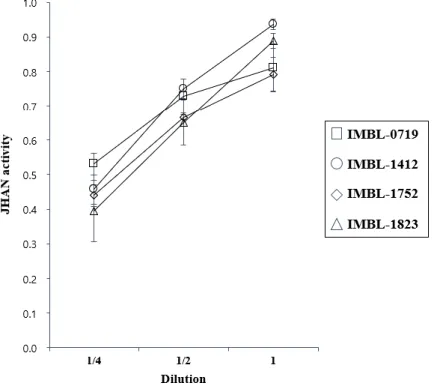
The IMBL-0719, IMBL-1412, IMBL-1752 and IMBL-1832 strains showing high insecticidal activities against P. xylostella were subjected to a concentration-dependent β-galactosidase assay to test the effects of increasing concentration on their JHAN activities. The absorbance of these actinobacteria culture media increased with regards to increasing concentration in the JHAN test (Fig. 9), while they showed no JHA activity even at high concentrations (data not shown).
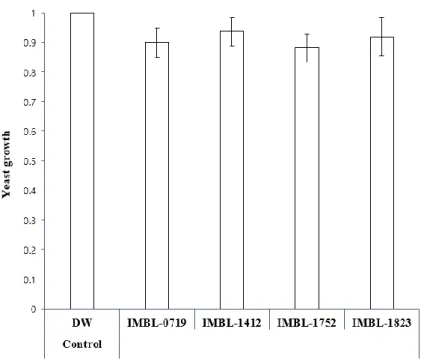
Yeast growth inhibition tests were conducted to investigate the possibility of false signals originating from the anti-yeast activities of actinobacteria (Fig. 10). The addition of actinobacteria culture media resulted in the normal growth of Y187 yeast cells transformed with Met and FISC in non-selective double dropout minimal (DDO, -Leu/-Trp) media, indicating that culture media of these actinobacteria strains directly disrupt the JH receptor complex and exhibit JHAN activity.
In addition, insecticidal activities of these actinobacteria strains against P. xylostella larvae were increased with increasing concentration of culture media (Fig. 11). Among 4 strains, the IMBL-1412 and IMBL-1823 strains showed superior insecticidal activities with mortalities greater than 60% at a concentration of 2-fold dilution, and were selected for further studies.
36
Figure 9. Concentration-dependent JHAN activities of actinobacteria strains. To estimate
the JHAN activity, 0.033 μg/ml of pyriproxyfen and serially diluted culture media of each strain were applied to a yeast two-hybrid β-galactosidase assay.
37
Figure 10. Anti-yeast activity tests of actinobacteria strains with JHAN activities. Culture media of each strain were tested for their anti-yeast activity to investigate whether the reduced β-galactosidase activity was resulted from JHAN activity or yeast toxicity.
38
Figure 11. Concentration-dependent insecticidal activities of actinobacteria strains against
P. xylostella. Third instar larvae of P. xylostella were treated with serially diluted culture
media of each actinobacteria strain using cabbage leaf dipping method and the mortality was calculated at 5 days after treatment.
39
4. Taxonomic identification of the selected actinobacteria strains
The cultural and physiological characteristics of the IMBL-1412 and IMBL-1823 strains observed at 14 days after growing on ISP media (Table 1). Aerial mycelium produced by the selected strains on all media were grey in color. Whereas substrate mycelium produced by the selected strains on ISP2 and ISP5 were brown in color, those on ISP4 were dark brown in color, suggesting that these strains belong to the genus Streptomyces.
For further phylogenetic profiling of these strains, the nucleotide sequence of their 16S rRNA gene was compared with those of representative Streptomyces strains. A phylogenetic tree constructed using the neighbor-joining method showed that both strains were most closely related to Streptomyces lactacystinicus OM-6519 with 99.5% 16S rRNA nucleotide sequence similarity (Fig. 12). Although nucleotide sequence of 16S rRNA were identical, these two strains showed differences in nucleotide sequence of another marker gene, recA, suggesting that they belong to different subspecies each other (Fig. 13).
40
41
Figure 12. Phylogenetic relationship of the IMBL-1412 and IMBL-1823 strains based on their nucleotide sequence of 16S rRNA gene. The nucleotide sequences of 16S rRNA gene from the genus Streptomyces were compared by the neighbor-joining method. Numbers at each branch node indicate the bootstrap percentage of 1,000 replications.
42
Figure 13. Phylogenetic relationship of the IMBL-1412 and IMBL-1823 strains based on their nucleotide sequence of recA gene. The nucleotide sequences of recA gene from the genus Streptomyces were compared by the neighbor-joining method. Numbers at each branch node indicate the bootstrap percentage of 1,000 replications.
43
5. Activities of the selected actinobacteria culture extracts
To purify JHAN compounds from the IMBL-1412 and IMBL-1823 strains, culture media of these strains were sequentially extracted with n-hexane, ethyl acetate and n-butanol, respectively. Whereas non-polar n-hexane and high-polar n-butanol fractions showed very low level of JHAN activity, ethyl acetate fraction of both strains displayed high JHAN activity at a concentration of 100 ppm (Fig. 14 and 15). Also, these fractions exhibited increasing JHAN activities with increasing concentrations, demonstrating that compounds extracted with ethyl acetate interfere with the pyriproxyfen-mediated binding of A. aegypti Met-FISC in a concentration-dependent manner.
In addition, insecticidal activities of the ethyl acetate fraction of 1412 and IMBL-1823 strains showing high JHAN activity were investigated against P. xylostella. Ethyl acetate fraction of both strains showed insecticidal activities in a concentration-dependent manner and the highest activity at a concentration of 100 ppm, with mortalities of 100% (Fig. 16 and 17).
44
Figure 14. Concentration-dependent JHAN activities of the strain IMBL-1412 culture extracts. To estimate the JHAN activity, 0.033 μg/ml of pyriproxyfen and corresponding
concentrations (5, 10, 50 and 100 ppm) of each fraction were applied to a yeast two-hybrid
45
Figure 15. Concentration-dependent JHAN activities of the strain IMBL-1823 culture extracts. To estimate the JHAN activity, 0.033 μg/ml of pyriproxyfen and corresponding
concentrations (5, 10, 50 and 100 ppm) of each fraction were applied to a yeast two-hybrid
46
Figure 16. Concentration-dependent insecticidal activity of the strain IMBL-1412 culture extracts against P. xylostella. Third instar larvae of P. xylostella were treated with
corresponding concentrations (1, 10, 50 and 100 ppm) of each fraction using cabbage leaf dipping method and the mortality was calculated at 5 days after treatment.
47
Figure 17. Concentration-dependent insecticidal activity of the strain IMBL-1823 culture extracts against P. xylostella. Third instar larvae of P. xylostella were treated with
corresponding concentrations (1, 10, 50 and 100 ppm) of each fraction using cabbage leaf dipping method and the mortality was calculated at 5 days after treatment.
48
DISCUSSION
During the last several decades, studies searching for eco-friendly insecticidal compounds have been extensively carried out to overcome the undesirable effects caused by synthetic insecticides such as toxicity to the environment and the development of resistant pests. Many natural products originating from plants and microorganisms have been successfully utilized as environmentally benign alternatives to synthetic insecticides (Cantrell, Dayan, and Duke 2012). Specifically, actinobacteria have been regarded as plentiful sources for natural products as they have been reported to produce various secondary metabolites with a diverse range of biological activities, including insecticidal, antifeedant, and insect growth inhibitory activities (Vijayabharathi et al. 2014; Arasu et al. 2013; Omura 2011). In this study, among 1,904 actinobacteria isolated from Korean soil samples, culture media of 25 strains showed high JHAN activities, with the culture media of IMBL-1412 and IMBL-1823 strains causing 100% mortality against P. xylostella larvae, demonstrating that these strains might produce novel JHAN compounds with high insecticidal activities. Several endophytic actinobacteria have been reported to play a role as beneficial symbionts with plants, and diverse bioactive metabolites from symbiotic actinobacteria could have been utilized by plants to endure environmental stresses, such as plant diseases and insect pests, over the course of plant and actinobacteria coevolution (Kinkel et al. 2012; Qin et al. 2011). Thus, potential JHAN compounds derived from endophytic actinobacteria might be adopted as effective defense mechanisms for symbiotic plants because IGRs are insect-specific, and insects may have difficulty acquiring
49
resistance to these IGRs (W.S. Bowers 2012). These results provided novel insights into the interactions between actinobacteria, plants, and insects.
Botanical precocenes isolated from Ageratum houstonianum and their synthetic analogues have also been reported to show JHAN activities (Banerjee et al. 2008). They cause precocious metamorphosis in several insect species by inducing irreversible degeneration of the corpora allata and inhibiting JH production (Azambuja and Garcia 1991). However, precocenes have not been commercialized as insecticides because of their low insecticidal activities and potential carcinogenic effects in mammals (Alzogaray and Zerba 2017). Because the JHAN compounds from actinobacteria identified in this study interrupt the formation of the JH receptor complex by competing with JH for binding to Met, their mode of action as JHANs is different from that of precocenes. Additionally, the high insecticidal activities of these compounds suggested that actinobacteria might be plentiful sources of novel IGR compounds, which could be exploited for the development of biopesticides.
50
LITERATURES CITED
Ababutain, Ibtisam M, ZK Abdul Aziz, and Nijla A AL-Meshhen. 2012. "Lincomycin antibiotic biosynthesis produced by Streptomyces sp. isolated from SAUDI ARABIA soil II-extraction, separation and purification of lincomycin." Canadian
Journal of Pure and Applied Sciences 6 (2): 1905-1911.
Alzogaray, Raúl A, and Eduardo N Zerba. 2017. "Rhodnius prolixus intoxicated." Journal
of insect physiology 97: 93-113.
Ando, Kunio, Hideo Oishi, Seiji Hirano, Tsuneo Okutomi, KOJI SUZUKI, HIROSHI OKAZAKI, MIKIO SAWADA, and TAKAO SAGAWA. 1971. "Tetranactin, a new miticidal antibiotic." The Journal of antibiotics 24 (6): 347-352.
Arasu, Mariadhas Valan, Naif Abdullah Al-Dhabi, Valsalam Saritha, Veeramuthu Duraipandiyan, Chinnasamy Muthukumar, and Sun-Ju Kim. 2013. "Antifeedant, larvicidal and growth inhibitory bioactivities of novel polyketide metabolite isolated from Streptomyces sp. AP-123 against Helicoverpa armigera and Spodoptera litura." BMC microbiology 13 (1): 105.
ASANO, Shoji, Eiichi KUWANO, and Morifusa ETO. 1986. "Anti-juvenile hormone activity of imidazole compound (kk-22) and its diminution by methoprene in the 4th instar silkworm, Bombyx mori L.: Lepidoptera: Bombycidae." Applied
Entomology and Zoology 21 (1): 63-69.
Azambuja, Patricia de, and Eloi S Garcia. 1991. "Effects of proallatotoxins (precocenes) on the development and reproduction of Rhodnius prolixus: some data." Memórias
51
Banerjee, Smita, GP Kalena, A Banerji, and AP Singh. 2008. "New synthetic precocenoids as potential insect control agents." J Environ Biol 95: 1-957.
Barovsky, K, and G Brooker. 1980. "(-)-[125I]-iodopindolol, a new highly selective radioiodinated beta-adrenergic receptor antagonist: measurement of beta-receptors on intact rat astrocytoma cells." Journal of cyclic nucleotide research 6 (4): 297-307.
Běhal, Vladislav. 2000. "Bioactive products from Streptomyces." Adv. Appl. Microbiol 47: 113-156.
Bergot, BJ, DA Schooley, and CAD De Kort. 1981. "Identification of JH III as the princpal juvenile hormone inLocusta migratoria." Experientia 37 (8): 909-910.
Bowers, WILLIAM S. 1977. "Anti-juvenile hormones from plants: chemistry and biological activity." GB Marini-Bettolo, Natural Products and the Protection of
Plants, Proceedings of the Pontifical Academy of Science, Vatican City: 129-142.
Bowers, WILLIAM S. 1981. "How anti-juvenile hormones work." American Zoologist 21 (3): 737-742.
Bowers, WILLIAM S. 2012. "Insect hormones and antihormones in plants." Herbivores:
Their Interactions with Secondary Plant Metabolites (GA Rosenthal and MR Berenbaum, eds.): 431-456.
Bowers, William S, and William G Bodenstein. 1971. "Sex pheromone mimics of the American cockroach." Nature 232 (5308): 259.
Bowers, WS. 1976. "Discovery of insect antiallatotropins." In The juvenile hormones, 394-408. Springer.
52
resistance mechanisms in stored-product insects." Bulletin of entomological
research 102 (2): 213-229.
Cantrell, Charles L, Franck E Dayan, and Stephen O Duke. 2012. "Natural products as sources for new pesticides." Journal of Natural Products 75 (6): 1231-1242. Charles, Jean-Philippe, Thomas Iwema, V Chandana Epa, Keiko Takaki, Jan Rynes, and
Marek Jindra. 2011. "Ligand-binding properties of a juvenile hormone receptor, Methoprene-tolerant." Proceedings of the National Academy of Sciences 108 (52): 21128-21133.
Cusson, Michel, Stephanie E Sen, and Tetsuro Shinoda. 2013. "Juvenile hormone biosynthetic enzymes as targets for insecticide discovery." In Advanced
Technologies for Managing Insect Pests, 31-55. Springer.
Dhadialla, Tarlochan S, Arthur Retnakaran, and Guy Smagghe. 2009. "Insect growth-and development-disrupting insecticides." In Insect development: morphogenesis,
molting and metamorphosis, 679-740. Academic Press.
Dhanasekaran, D, V Sakthi, N Thajuddin, and A Panneerselvam. 2010. "Preliminary evaluation of Anopheles mosquito larvicidal efficacy of mangrove actinobacteria."
Int J Appl Biol Pharm Technol 1 (2): 374-381.
El-Khawagh, MA, Kh Sh Hamadah, and TM El-Sheikh. 2011. "The insecticidal activity of Actinomycete metabolites against the mosquito Culex pipiens." Egypt. Acad. J.
Biol. Sci 4: 103-113.
Hamnett, Anthony F, Anthony P Ottridge, Grahame E Pratt, Richard C Jennings, and Katherine M Stott. 1981. "Kinetics and products of the hydrolysis of 3, 4‐ dihydroprecocene I 3, 4‐epoxide in aqueous organic solvents." Pesticide Science
53 12 (3): 245-254.
Hartfelder, Klaus, and DJ Emlen. 2012. "Endocrine control of insect polyphenism." In
Insect endocrinology, 464-522. Elsevier.
HATAKOSHI, Makoto, Noriaki AGUI, and Isamu NAKAYAMA. 1986. "2-[1-Methyl-2-(4-phenoxyphenoxy) ethoxy] pyridine as a new insect juvenile hormone analogue: induction of supernumerary larvae in Spodoptera litura (Lepidoptera: Noctuidae)."
Applied Entomology and Zoology 21 (2): 351-353.
HENRICK, CLIVE A. 1982. "Juvenile hormone analogs: Structure-activity relationships." In Insecticide mode of action, 315-402. Elsevier.
Hill, Catherine A, Fotis C Kafatos, Sally K Stansfield, and Frank H Collins. 2005. "Arthropod-borne diseases: vector control in the genomics era." Nature Reviews
Microbiology 3 (3): 262.
Ilic, SB, SS Konstantinovic, ZB Todorovic, ML Lazic, VB Veljkovic, N Jokovic, and BC Radovanovic. 2007. "Characterization and antimicrobial activity of the bioactive metabolites in streptomycete isolates." Microbiology 76 (4): 421-428.
Jackson, Stephen, Lisa Crossman, Eduardo Almeida, Lekha Margassery, Jonathan Kennedy, and Alan Dobson. 2018. "Diverse and abundant secondary metabolism biosynthetic gene clusters in the genomes of marine sponge derived Streptomyces spp. isolates." Marine drugs 16 (2): 67.
Jindra, Marek, Subba R Palli, and Lynn M Riddiford. 2013. "The juvenile hormone signaling pathway in insect development." Annual review of entomology 58: 181-204.
54
1991. "Macrotetrolide antibiotics produced by Streptomyces globisporus." Folia
microbiologica 36 (5): 437-443.
Jones, David T, William R Taylor, and Janet M Thornton. 1992. "The rapid generation of mutation data matrices from protein sequences." Bioinformatics 8 (3): 275-282. Judy, Kenneth J, David A Schooley, Loren L Dunham, MS Hall, B John Bergot, and John
B Siddall. 1973. "Isolation, structure, and absolute configuration of a new natural insect juvenile hormone from Manduca sexta." Proceedings of the National
Academy of Sciences 70 (5): 1509-1513.
Karthik, L, K Gaurav, KV Bhaskara Rao, G Rajakumar, and A Abdul Rahuman. 2011. "Larvicidal, repellent, and ovicidal activity of marine actinobacteria extracts against Culex tritaeniorhynchus and Culex gelidus." Parasitology research 108 (6): 1447-1455.
Kaur, Talwinder, Arti Vasudev, Satwinder Kaur Sohal, and Rajesh Kumari Manhas. 2014. "Insecticidal and growth inhibitory potential of Streptomyces hydrogenans DH16 on major pest of India, S podoptera litura (Fab.)(Lepidoptera: Noctuidae)." BMC
microbiology 14 (1): 227.
Kelly, Kenneth Low, and Deane Brewster Judd. 1965. "ISCC-NBS color-name charts illustrated with centroid colors."
Kewley, Robyn J, Murray L Whitelaw, and Anne Chapman-Smith. 2004. "The mammalian basic helix–loop–helix/PAS family of transcriptional regulators." The international
journal of biochemistry & cell biology 36 (2): 189-204.
Kinkel, Linda L, Daniel C Schlatter, Matthew G Bakker, and Brett E Arenz. 2012. "Streptomyces competition and co-evolution in relation to plant disease
55
suppression." Research in microbiology 163 (8): 490-499.
Lechevalier, H, and Mary P Lechevalier. 1965. "Classification des actinomycètes aérobies basée sur leur morphologie et leur composition chimique." ANNALES DE L INSTITUT PASTEUR.
Lee, Seok-Hee, Kyu Baik Ha, Dong Hwan Park, Ying Fang, Jong Hoon Kim, Min Gu Park, Ra Mi Woo, Woo Jin Kim, Il-Kwon Park, and Jae Young Choi. 2018. "Plant-derived compounds regulate formation of the insect juvenile hormone receptor complex." Pesticide biochemistry and physiology 150: 27-32.
Lee, Seok-Hee, Hee Nam Lim, Jae Young Choi, Dong Hwan Park, Byung Hoon Ahn, Ying Fang, Jong Hoon Kim, Min Gu Park, Ra Mi Woo, and Bo Ram Lee. 2018. "Mosquitocidal activity of penfluridol as juvenile hormone antagonist." Journal of
Asia-Pacific Entomology 21 (1): 130-133.
Lee, Seok-Hee, Hyun-Woo Oh, Ying Fang, Saes-Byeol An, Doo-Sang Park, Hyuk-Hwan Song, Sei-Ryang Oh, Soo-Young Kim, Seonghyun Kim, and Namjung Kim. 2015. "Identification of plant compounds that disrupt the insect juvenile hormone receptor complex." Proceedings of the National Academy of Sciences 112 (6): 1733-1738.
Lee, Seok‐Hee, Jae Young Choi, Bo Ram Lee, Ying Fang, Jong Hoon Kim, Dong Hwan Park, Min Gu Park, Ra Mi Woo, Woo Jin Kim, and Yeon Ho Je. 2018. "Insect growth regulatory and larvicidal activity of chalcones against Aedes albopictus."
Entomological research 48 (1): 55-59.
Li, Meng, Edward A Mead, and Jinsong Zhu. 2011. "Heterodimer of two bHLH-PAS proteins mediates juvenile hormone-induced gene expression." Proceedings of the
56
National Academy of Sciences 108 (2): 638-643.
Li, Wen-Jun, Ping Xu, Peter Schumann, Yu-Qin Zhang, Rüdiger Pukall, Li-Hua Xu, Erko Stackebrandt, and Cheng-Lin Jiang. 2007. "Georgenia ruanii sp. nov., a novel actinobacterium isolated from forest soil in Yunnan (China), and emended description of the genus Georgenia." International Journal of Systematic and
Evolutionary Microbiology 57 (7): 1424-1428.
Mahajan, Girish, and Lakshmi Balachandran. 2014. "Biodiversity in production of antibiotics and other bioactive compounds." In Biotechnological Applications of
Biodiversity, 37-58. Springer.
Masner, P, S Dorn, W Vogel, M Kalin, and O Graf. 1981. "Types of response of insects to a new IGR and to proven standards." Regulation of insect development and behavior: International Conference, Karpacz, Poland, June 23-28 1980/[editor-in-chief, Marian Kloza].
Meyer, Andre S, Howard A Schneiderman, Edith Hanzmann, and Jane H Ko. 1968. "The two juvenile hormones from the Cecropia silk moth." Proceedings of the National
Academy of Sciences of the United States of America 60 (3): 853.
Meyers, Paul R. 2015. "Analysis of recombinase A (recA/RecA) in the actinobacterial family Streptosporangiaceae and identification of molecular signatures."
Systematic and applied microbiology 38 (8): 567-577.
Montesinos, Emilio. 2003. "Development, registration and commercialization of microbial pesticides for plant protection." International Microbiology 6 (4): 245-252. Nijhout, H Frederik. 1998. Insect hormones. Princeton University Press.
57
HAYASHI, MIKIO SAWADA, and KUNIO ANDO. 1970. "Insecticidal activity of macrotetrolide antibiotics." The Journal of antibiotics 23 (2): 105-106.
Omura, Satoshi. 2011. "Microbial metabolites: 45 years of wandering, wondering and discovering." Natural Product Updates 67 (35): 6420-6459.
Ōmura, Satoshi, and Andy Crump. 2014. "Ivermectin: panacea for resource-poor communities?" Trends in parasitology 30 (9): 445-455.
Pener, Meir Paul, and Tarlochan S Dhadialla. 2012. "An overview of insect growth disruptors; applied aspects." In Advances in Insect Physiology, 1-162. Elsevier. Qin, Sheng, Ke Xing, Ji-Hong Jiang, Li-Hua Xu, and Wen-Jun Li. 2011. "Biodiversity,
bioactive natural products and biotechnological potential of plant-associated endophytic actinobacteria." Applied Microbiology and Biotechnology 89 (3): 457-473.
Quistad, Gary B, David C Cerf, David A Schooley, and Gerardus B Staal. 1981. "Fluoromevalonate acts as an inhibitor of insect juvenile hormone biosynthesis."
Nature 289 (5794): 176-177.
Raikhel, AS, M Brown, and X Belles. 2005. "3.9 Hormonal Control of Reproductive Processes." Comprehensive molecular insect science: 433-491.
Richard, David S, Shalom W Applebaum, and Lawrence I Gilbert. 1989. "Developmental regulation of juvenile hormone biosynthesis by the ring gland ofDrosophila melanogaster." Journal of Comparative Physiology B 159 (4): 383-387.
Riddiford, Lynn M. 1994. "Cellular and molecular actions of juvenile hormone I. General considerations and premetamorphic actions." In Advances in insect physiology, 213-274. Elsevier.
58
Riddiford, Lynn M. 2008. "Juvenile hormone action: a 2007 perspective." Journal of insect
physiology 54 (6): 895-901.
Röller, H, KH Dahm, CC Sweely, and BM Trost. 1967. "The structure of the juvenile hormone." Angewandte Chemie International Edition in English 6 (2): 179-180. Sagawa, Takao, Seiji Hirano, Hironobu Takahashi, Nobuhiko Tanaka, Hideo Oishi, Kunio
Ando, and Kunihiko Togashi. 1972. "Tetranactin, a new miticidal antibiotic. III. Miticidal and other biological properties." Journal of economic entomology 65 (2): 372-375.
Sakamoto, K, T Asano, K Mizuochi, K Sasaki, and K Hasegawa. 1978. "Makrotetrolid-Antibiotikum zur Bekämpfung der Geflügelkokzidiose." Chem Abstr.
Samri, SE, M Baz, I Ghalbane, S El Messoussi, A Zitouni, A El Meziane, and M Barakate. 2017. "Insecticidal activity of a Moroccan strain of Streptomyces
phaeochromogenes LD-37 on larvae, pupae and adults of the Mediterranean fruit
fly, Ceratitis capitata (Diptera: Tephritidae)." Bulletin of entomological research 107 (2): 217-224.
Schatz, Albert, and Selman A Waksman. 1944. "Effect of Streptomycin and Other Antibiotic Substances upon Mycobacterium tuberculosis and Related Organisms."
Proceedings of the Society for Experimental Biology and Medicine 57 (2):
244-248.
Schooley, DA, FC Baker, LW Tsai, CA Miller, and GC Jamieson. 1984. "Juvenile hormones O, I, and II exist only in Lepidoptera." In Biosynthesis, metabolism and mode of
action of invertebrate hormones, 373-383. Springer.
59
identification, and their applications." Int J Curr Microbiol App Sci 3 (2): 801-832. Shichi, Hitoshi, Yoshihito Tanouchi, and Yoshio Kamada. 1989. Immunosuppressive agent.
Google Patents.
Shin, Sang Woon, Zhen Zou, Tusar T Saha, and Alexander S Raikhel. 2012. "bHLH-PAS heterodimer of methoprene-tolerant and Cycle mediates circadian expression of juvenile hormone-induced mosquito genes." Proceedings of the National Academy
of Sciences 109 (41): 16576-16581.
Slama, KAREL. 1971. "Insect juvenile hormone analogues." Annual review of
biochemistry 40 (1): 1079-1102.
Sláma, KAREL. 1999. "The history and current status of juvenoids." Proceedings of 3rd International Conference on Urban Pests.
Sláma, Karel, and Carroll M Williams. 1966a. "The juvenile hormone. V. The sensitivity of the bug, Pyrrhocoris apterus, to a hormonally active factor in American paper-pulp." The Biological Bulletin 130 (2): 235-246.
Sláma, Karel, and Carroll M Williams. 1966b. "‘Paper factor’as an inhibitor of the embryonic development of the European bug, Pyrrhocoris apterus." Nature 210 (5033): 329-330.
Solecka, Jolanta, Joanna Zajko, Magdalena Postek, and Aleksandra Rajnisz. 2012. "Biologically active secondary metabolites from Actinomycetes." Open Life
Sciences 7 (3): 373-390.
Staal, GB. 1986. "Anti juvenile hormone agents." Ann. Rev. Entomol. 31: 391-429. Thompson, Julie D, Toby J Gibson, Frédéric Plewniak, François Jeanmougin, and
60
strategies for multiple sequence alignment aided by quality analysis tools." Nucleic
acids research 25 (24): 4876-4882.
Tindall, Brian J, Peter Kämpfer, Jean P Euzéby, and Aharon Oren. 2006. "Valid publication of names of prokaryotes according to the rules of nomenclature: past history and current practice." International Journal of Systematic and Evolutionary
Microbiology 56 (11): 2715-2720.
Unnithan, Gopalan C, John F Andersen, Tomomi Hisano, Eiichi Kuwano, and René Feyereisen. 1995. "Inhibition of juvenile hormone biosynthesis and methyl farnesoate epoxidase activity by 1, 5‐disubstituted imidazoles in the cockroach, Diploptera punctata." Pesticide science 43 (1): 13-19.
Vijayabharathi, Rajendran, Bhimineni Ratna Kumari, Arumugam Sathya, Vadlamudi Srinivas, Rathore Abhishek, Hari C Sharma, and Subramaniam Gopalakrishnan. 2014. "Biological activity of entomopathogenic actinomycetes against lepidopteran insects (Noctuidae: Lepidoptera)." Canadian Journal of Plant
Science 94 (4): 759-769.
Waksman, Selman A, and H Boyd Woodruff. 1940. "Bacteriostatic and Bactericidal Substances Produced by a Soil Actinomyces." Proceedings of the society for
Experimental Biology and Medicine 45 (2): 609-614.
Waksman, Selman A, and H Boyd Woodruff. 1942. "Selective antibiotic action of various substances of microbial origin." Journal of bacteriology 44 (3): 373.
Wigglesworth, VB. 1961. "Some observations on the juvenile hormone effect of farnesol in Rhodnius prolixus Stål (Hemiptera)." Journal of Insect Physiology 7 (1): 73-78. Wigglesworth, Vincent B. 1934. "Memoirs: The physiology of ecdysis in Rhodnius
61
prolixus (Hemiptera). II. Factors controlling moulting and ‘metamorphosis’."
Journal of Cell Science 2 (306): 191-222.
Wigglesworth, Vincent B. 1936. "Memoirs: the function of the Corpus Allatum in the growth and reproduction of Rhodnius Prolixus (Hemiptera)." Journal of Cell
Science 2 (313): 91-121.
Zimmermann, W. 1990. "Degradation of lignin by bacteria." Journal of biotechnology 13 (2-3): 119-130.
62
ABSTRACT IN KOREAN
방선균의
곤충 유약호르몬 길항제 탐색 및
배추좀나방
살충활성 검정
서울대학교
농생명공학부 곤충학전공
김준영
초 록
곤충생장조절제 (insect growth regulator: IGR)은 화학 살충제에 비해 상대
적으로 높은 특이성과 환경에 대해 낮은 독성 등의 장점을 가지기 때문에 화 학 농약의 대안으로 떠오르고 있다. 그램 양성에 속하는 토양 세균인 방선균 은 배양 과정 중에 살충 및 곤충 생장 조절 활성을 가지는 다양한 2차 대사 산물을 생산하는 것으로 알려져 있어서 새로운 IGR 활성 물질의 잠재적인 source로 활용될 수 있다. 본 연구에서는 방선균으로부터 새로운 IGR 물질을 탐색하기 위하여, 국내토양으로부터 분리된 1,904개 방선균 균주 배양액의 IGR 활성을 평가하였다. 그 결과, 25개의 균주가 0.4 이상의 높은 JHAN
63
(juvenile hormone antagonist) 활성을 보였다. 우수한 JHAN 활성을 보인 방
선균 균주 중, 4개의 균주 배양액이 배추좀나방 3령 유충에 대하여 우수한 살
충 활성을 보여주었다. 그 중, 상대적으로 높은 JHAN 및 살충 활성을 보인
IMBL-1412와 IMBL-1823 균주는 ISP 배지 상에서의 배양 생리학적 특성과
16S rRNA 유전자 염기서열을 이용한 분자생물학적 동정을 통하여
Streptomyces lactacystinicus 균주인 것으로 나타났다. 또한, 이 두 균주는
recA 유전자 염기서열에서 차이를 보임으로써 Streptomyces lactacystinicus
에 속하는 서로 다른 subspecies임을 확인할 수 있었다. IMBL-1412와
IMBL-1823 균주가 생산하는 JHAN 활성 물질을 분리하기 위하여, 두 균주의
배양액을 다양한 용매를 이용하여 순차적으로 추출하고 각각의 추출물에 대하
여 IGR 및 살충 활성을 조사하였다. 그 결과, ethyl acetate 추출물이 높은
JHAN 활성과 배추좀나방 유충에 대한 살충 활성을 가지는 것을 확인할 수
있었다. 이러한 결과는 방선균 유래의 2차 대사산물이 새로운 IGR 살충제로
유용하게 이용될 수 있음을 시사하였다.
Key words : 방선균, Streptomyces lactacystinicus, 곤충생장조절제, 유충
호르몬 길항제, 배추좀나방