INTRODUCTION
Chitin is an insoluble linear polymer of β-1, 4-linked N-acetyl glucosamine (GlcNAc) residue and is a major con-stituent not only of fungal cell walls (22% to 44%) but also of insect exoskeletons and crustacean shells (25% to 58%) (Muzzarelli 1999; Patil et al. 2000). Cell wall-degrading enzymes including chitinases are involved in the biologic control of phytopathogenic fungi by rhizobacteria. The pro-duction of chitinases by plants is considered to be a part of their defence mechanism against fungal pathogens. It is suggested that chitinolytic microorganisms or chitinolytic
enzymes have potential applications in the biocontrol of plant pathogenic fungi and insects (Shapira et al. 1989; Lorito et al. 1993a), as a target for biopesticides (Sakuda 1996), and in many other biotechnological areas (Shaikh and Desh-pande 1993; Patil et al. 2000). Biological control of plant pathogens provides an attractive alternative means for man-agement of plant disease without the negative impact of chemical fungicides that are usually costly, can cause envi-ronmental pollution, and may induce pathogen resistance. Several mycoparasites that grow on rust fungi produce lytic enzymes (Saksirirat and Hoppe 1991; Mathivanan et al. 1997). Among these, chitinase plays a vital role in the biolog-ical control of many plant diseases by degrading the chitin polymer in the cell walls of fungal pathogens (Barrows-Broadders and Kerr 1981; Haran et al. 1993). It affects fun-gal growth through the lysis of cell walls (Elad et al. 1982;
─ ─ 65 ──
Gamma Radiation Induced Mutagenesis of
Lysobacter enzymogenes for Enhanced Chitinolytic Activity
Young-Keun Lee, Kyoung Youl Kim and M. Senthilkumar*Advanced Radiation Technology Institute, Korea Atomic Energy Research Institute, Sinjeong, Jeongeup 580-185, Korea
Abstract -- Two chitinase producing strains CHI2 and CHI4 were isolated from soybean rhizos-phere soil. Both the strains belonged to Lysobacter enzymogenes as indicated by 16S rDNA sequence analysis. Though strain CHI2 and CHI4 produced extracellular chitinase, they differ in their chiti-nolytic activity. CHI4 produced approximately three times the higher amounts of enzyme than that of CHI2 under specified conditions. CHI2 produced 535.67 U l--1of chitinase after 48 h incubation with a specific activity of 3.91 U mg--1of protein while strain CHI4 produced 1584.13 U l--1of chiti-nase with a specific activity of 10.88 U mg--1protein. SDS-PAGE analysis indicated that the molecu-lar weight of chitinase enzyme was approximately 45 kDa. A faint band with a molecumolecu-lar weight of 55 kDa reveals the possibility for the presence of another kind of chitin binding protein. Mutant library was developed by exposing the isolates to gamma rays at their LD99value (0.23 kGy). Totally, 11 mutants of CHI2 and CHI4 are reported to have enhanced chitinase activity. Several leaky mutant clones with decreased enzyme activity and a defective mutant (CHI2-M16) with complete loss of chitinase activity were also identified. CHI4-M18, CHI4-M8 and CHI4-M29 showed 78.8, 41.5, and 31.9% increased chitinase activity over wild type CHI4.
Key words : Biocontrol, Gamma radiation, Chitinase, Lysobacter
* Corresponding authors: M. Senthilkumar, Tel. +82-63-570-3304, Fax. +82-63-570-3309, E-mail. [email protected]
germ tubes (Lorito et al. 1993b; Gunaratna and Balasubra-manian 1994). Chitinase has been purified from various fungi (Ulhoa and Peberdy 1992; Gunaratna and Balasubra-manian 1994; Sakurada et al. 1996) and its direct involve-ment in controlling plant diseases has also been demonstrated (Haran et al. 1993; Park et al. 1995; Madi et al. 1997). Bac-teria produce chitinase to digest chitin, primarily to utilize it as a carbon and energy source (Roberts and Selitrennikoff 1988; Gooday 1990; Leah et al. 1991). Chitinases are con-stituents of several bacterial species; some of the best known include the Aeromonas, Serratia, Myxobacter, Vibrio, Strep-tomyces, and Bacillus genera. Strains of Serratia marcescens, Bacillus, and Vibrio have been shown to produce a high level of chitinolytic enzymes (Cody et al. 1990). Enzymatic hydrolysis of chitin to free β-1, 4-linked N-acetylglucosamine (GlcNAc) is performed by a chitinolytic system consisting of chitinase and chitobiase, the actions of which are syner-gistic and consecutive (Deshpande 1986; Shaikh and Desh-pande 1993; Patil et al. 2000). The chitinases are classified as endochitinase, exochitinase (EC.3.2.1.14), β-N-actylglu-cosaminidase, and chitobiase (EC.3.2.1.30), which degrade chitin and its derivatives. The objective of this study was to isolate chitinase producing bacteria with potential antifungal activity and to improve the chitinase production through gamma radiation induced mutagenesis.
MATERIALS AND METHODS
1. Isolation and characterization of chitinaseproducing bacteria
Soil samples from soybean rhizosphere were serially dilut-ed and platdilut-ed on nutrient agar mdilut-edium supplementdilut-ed with 0.5% colloidal chitin (Wiwat et al. 1999). The colonies with large clear zones were purified and stored in 25% glycerol at 80�C as well as under refrigerated conditions. The colloidal chitin was obtained by a modification of the published method (Hsu and Lockwood 1975). Chitin (20 g) from crab shell (Sigma Co., practical grade) was dissolved in 200 ml of concentrated HCl with stirring for 3 min at 40�C. The chitin was precipitated as a colloidal suspension by slowly adding water (2 l) adjusted to 5�C. Colloidal suspensions were col-lected by filtering through coarse filter paper; then the filtered
pH of the suspension was about 7.0.
Chitinase producing isolates were identified based on 16S rDNA sequence analysis. Genomic DNA was extracted by using Qiagen DNeasy Plant Mini Kit (Qiagen, Valencia, CA). 16S rDNA was amplified by using the primers 27F (5′-AG AGTTTGATCCTGGCTCAG-3′) and 1492R (5′-GGTTA CCTTGTTACGACTT-3′) (Lane 1991). The sequences were determined by fluorescent dye terminators method using ABI prismTMBigdyeTMterminator cycle sequencing ready
reaction kit V.3.1. Products were run on ABI 3730XL capil-lary DNA sequencer (ABI prism 310 Genetic analyzer, Tokyo, Japan). Sequences were subjected to BLASTn analy-sis with the NCBI data base and phylogenetic trees were gen-erated with NJPLOT programme of CLUSTAL X (Thomp-son et al. 1997, Fig. 1).
2. Determination of chitinase activity
Chitinase activity was assayed by the method of Yanai et al. (1992). The reaction mixture containing 250μl of 0.5% colloidal chitin, 250μl of 0.2 M sodium acetate buffer (pH 4), and 500μl of enzyme solution was incubated for 2h at 37�C. After centrifugation, 500μl from the supernatant fluid was mixed with 100μl of 0.8 M boric acid, and the pH of this mixed solution was adjusted to 10.2 with KOH. The solution was heated for 3 min in boiling water. After the mixture was cooled, 3 ml of p-dimethyl aminobenzaldehyde (DMAB) solution (1 g of DMAB dissolved in 100 ml of glacial acetic acid containing 1% v/v hydrochloric acid) was added and the mixture was incubated for 20 min at 37�C. Absorbance at 585 nm was measured against water as a blank. One unit of chitinase activity was defined as the amount of enzyme which produced sugars equivalent to 1μmol of N-acetyl glucosamine per min under the above condition. Protein con-centration was determined according to Bradford (1976) using bovine serum albumin as a standard.
3. Radiation sensitivity
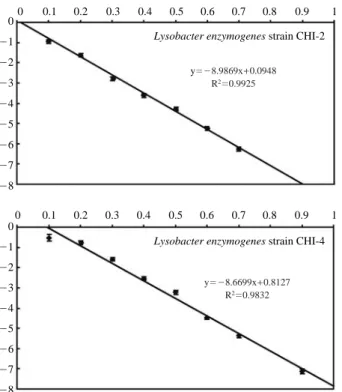
Cell pellets were collected by centrifuging 20 ml of log phase bacterial isolate cultured in LB broth. Pellets were washed twice with sterile distilled water and suspended (107~109CFUml-1). 500μl of cell suspensions were trans-ferred to 1.5 ml micro-centrifuge tubes and irradiated at dif-ferent doses expressed in kGy as shown in Fig. 2. Samples
were irradiated in cobalt-60 irradiator (capacity: 250000 Ci, dose rate 920 Gy hr-1, AECL) at Advanced Radiation
Tech-nology Institute, Korea Atomic Energy Research Institute, Republic of Korea. Bacterial population in irradiated samples was determined by serial dilution and plate count method. The D10-value was determined by plotting radiation dose
(kGy) on X-axis and survival ratio on Y-axis. Negative rec-iprocal of slope indicated the D10-value. LD99value-radiation
dose required to kill 99% of viable bacterial cells was deter-mined by using the formula LD99==log (0.01) slope-1.
4. Radiation mutagenesis and selection
To induce the mutagenesis, bacterial cell suspensions were
prepared as described above and exposed to gamma irradia-tion at the dose of LD99. Irradiated samples were serially
diluted and plated on chitin agar and incubated at 28±2�C for 48 h. Master plates were prepared from the mutant clones with enhanced as well as leaky or defective chitinase activity and stored under refrigerated conditions. Chitinase activity of mutant clones was determined by colorimetric method as described earlier and compared with wild type. Over produc-ing as well as defective mutants for chitinase activity were selected and stored as pure cultures.
5. SDS-PAGE analysis
The molecular weight of the chitinase was determined by Fig. 1. 16S rDNA sequence based phylogenetic tree showing the relationship of chitinase producing strains with other related genera. Bootstrap
1970). 25μl of protein sample (1 mg ml ) was applied to 12% polyacrylamide gel and electrophoresed at 20 mA along with molecular weight markers. Before electrophoresis,
pro-(pH 7) containing 2-mercaptoethanol. The gels were stained with Coomassie Brilliant Blue R-250 in methanol-acetic acid-water (4 : 1 : 5, v/v) and decolorized in 7% acetic acid
RESULTS AND DISCUSSION
Two chitinase producing strains CHI2 and CHI4 were iso-lated from the rhizosphere soil of soybean. They produced chitinase in the presence of 0.5% colloidal chitin and pro-duced clear zones around the colonies on chitin agar plates. Chitinase of CHI2 and CHI4 appeared to be extracellular, since the supernatant from the cell lysate prepared by sonica-tion did not show a detectable amount of chitinase activity. Both isolates are Gram negative bacteria, producing yellow pigmented colonies on nutrient agar plates. 16S rDNA analy-sis indicated that they belong to Lysobacter enzymogenes with sequence similarity of 99.59%. These isolates were tested for their inhibitory effect on hyphal growth of phyto-pathogens like Alternaria alternata, Alternaria solani, Rhi-zoctonia solani, and Botrytis cinerea. Both isolates inhibited the growth of tested fungi, and CHI2 recorded the largest zone of inhibition (12 mm) against B. cinerea. Extracellular proteins of CHI2 and CHI4, grown on 0.5% colloidal chitin Fig. 2. Radiation sensitivity of CHI2 and CHI4.
0 -1 -2 -3 -4 -5 -6 -7 -8 0 -1 -2 -3 -4 -5 -6 -7 -8 0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1 0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1
Lysobacter enzymogenes strain CHI-2
Lysobacter enzymogenes strain CHI-4 y=-8.9869x++0.0948
R2=0.9925
y=-8.6699x++0.8127 R2=0.9832
Table 1. Chitinase activity of mutant clones and wild type of strain CHI2
Bacteria Chitinase activity (U l-1) Protein (mg l-1) Specific activity (U mg-1protein)
CHI2 535.87±31.30l 137.04±8.70ef 3.91±0.05i CHI2-M1 505.65±09.71jk 140.25±3.19efg 3.61±0.04h CHI2-M2 859.04±14.03o 151.69±2.84h 5.66±0.03l CHI2-M3 806.89±22.52n 153.42±2.86h 5.26±0.06k CHI2-M4 366.82±07.27de 124.12±5.29d 2.96±0.07d CHI2-M5 341.19±17.16cd 111.14±4.04b 3.07±0.04e CHI2-M6 295.05±15.97a 114.78±4.79bc 2.57±0.03b CHI2-M7 487.49±12.56j 135.36±4.69e 3.60±0.05gh CHI2-M8 392.03±11.66ef 141.06±5.15efg 2.78±0.04c CHI2-M9 416.83±15.64fg 119.57±8.94bcd 3.49±0.13fg CHI2-M10 517.40±19.96kl 148.95±6.35gh 3.48±0.03f CHI2-M11 421.68±10.93gh 145.30±4.48fgh 2.91±0.06d CHI2-M12 363.91±11.75d 139.16±3.56ef 2.62±0.02b CHI2-M13 445.52±13.98hi 140.67±4.23efg 3.17±0.19e CHI2-M14 489.27±12.33j 133.62±3.41e 3.67±0.01h CHI2-M15 312.23±10.87ab 120.90±4.93cd 2.58±0.02b CHI2-M16 - 89.16±3.51a -CHI2-M17 591.13±14.67m 132.79±4.45e 4.45±0.04j CHI2-M18 434.75±13.75ghi 133.12±4.63ef 3.16±0.03e CHI2-M19 329.80±16.10bc 130.27±4.76e 2.44±0.04a CHI2-M20 451.46±14.97i 148.56±4.24h 2.95±0.03d
supplemented growth medium showed the inhibitory effect on hyphal growth of Botrytis cinerea. Acetone precipitated extracellular proteins were capable of degrading chitin and produced clear zone at the site of application on chitin agar plates. Though the strains shared a high level of 16S rDNA sequence similarity, they differed in their chitinolytic activ-ity. CHI2 produced 535.67 U l-1of chitinase on 48 h
incuba-tion with a specific activity of 3.91 U mg-1of protein while
strain CHI4 produced 1584.13 U l-1of chitinase with a
spe-cific activity of 10.88 U mg-1protein.
Gamma radiation induced mutagenesis was carried out in order to modify the level of chitinase activity. D10and LD99
of strain CHI2 and CHI4 were initially determined as 0.11, 0.22 kGy and 0.12, 0.23 kGy respectively. Mutant library was developed by exposing the isolates to gamma rays at LD99 value (0.23 kGy). Mutant clones were screened for
their altered chitinase activity. Of 20 mutants from CHI2, 3 mutants are reported to have enhanced chitinase activity.
Several leaky mutant clones with a decreased enzyme activ-ity and a defective mutant (CHI2-M16) with loss of chitinase activity were also identified. CHI2-M2 showed the highest chitinase activity, 859.04 U l-1i.e. 60.3% over the wild type
CHI2 with a specific activity of 5.66 U mg-1protein. Another
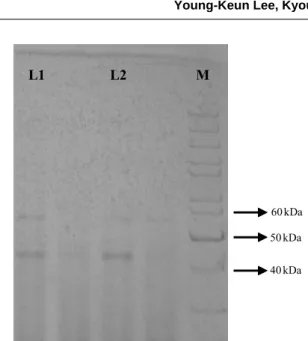
mutant CHI2-M3 recorded 50.6% increased chitinase activ-ity over wild type (Table 1). In the case of CHI4, 30 mutant clones were identified with altered chitinase activity. Eight mutant clones were reported to have enhanced chitinase activity while others are leaky mutants with a decreased enzy-me activity. CHI4-M18, CHI4-M8 and CHI4-M29 showed 78.8, 41.5, and 31.9% increased chitinase activity over the wild type CHI4 (Table 2). SDS PAGE analysis indicated that the molecular weight of chitinase enzyme was 45 kDa (Fig. 3). Chitinase with similar molecular weight was purified from Alcaligenes xylosoxydans (Vaidya et al. 2003) and Bacillus circulans No. 41 (Wiwat et al. 1999). Microbial chitinases varied in their molecular weight ranging from 20 Table 2. Chitinase activity of mutant clones and wild type of strain CHI4
Bacteria Chitinase activity (U l-1) Protein (mg l-1) Specific activity (U mg-1protein)
CHI4 1584.13±27.70k 145.71±4.40m 10.88±0.36efgh CHI4-M1 1683.68±14.49m 44.31±1.72a 38.03±1.13m CHI4-M2 1105.72±25.34g 78.37±3.29cd 14.12±0.27ij CHI4-M3 821.56±19.73bc 76.02±7.94c 10.87±0.86efgh CHI4-M4 940.04±23.83e 81.84±7.47cdef 11.54±0.77gh CHI4-M5 1144.66±20.18h 109.74±8.14jkl 10.46±0.68defgh CHI4-M6 1017.34±27.01f 82.00±3.10cdef 12.41±0.19hi
CHI4-M7 858.44±24.75cd 98.28±8.91hij 8.77±0.54abcde
CHI4-M8 2240.70±25.85p 44.56±4.37a 54.39±1.74p
CHI4-M9 936.10±26.41e 97.88±7.89hij 9.59±0.51cdefg
CHI4-M10 1229.58±32.88l 100.96±8.98hijk 12.23±0.95hi CHI4-M11 806.72±20.71b 113.38±7.73l 7.13±0.31a CHI4-M12 1027.99±23.40f 91.34±6.43efgh 11.28±0.54fgh CHI4-M13 918.89±32.83e 111.14±7.97kl 8.28±0.32abc CHI4-M14 1626.82±24.26l 84.74±5.39cdefg 19.24±0.95k CHI4-M15 881.06±14.70d 76.25±5.65c 11.59±0.66gh CHI4-M16 1224.39±13.87i 112.32±6.10kl 10.92±0.46efgh
CHI4-M17 828.39±11.44bc 89.50±4.96defgh 9.27±0.38bcdef
CHI4-M18 2832.10±32.73q 63.15±4.18b 44.96±2.51o
CHI4-M19 1023.48±21.90f 81.61±6.21cde 12.58±0.71hi
CHI4-M20 1130.36±22.30g 104.65±9.09ijkl 10.84±0.72efgh
CHI4-M21 1619.16±17.95kl 50.57±3.30a 32.10±1.78l
CHI4-M22 2108.11±24.31o 50.73±2.04a 42.19±2.11n
CHI4-M23 1864.58±20.57n 98.45±9.73hij 19.06±1.70k
CHI4-M24 989.46±15.77f 92.07±4.66efgh 10.76±0.38efgh
CHI4-M25 798.00±23.11b 55.16±6.30ab 14.57±1.26j
CHI4-M26 788.67±14.40b 93.86±8.25fghi 8.44±0.59abcd
CHI4-M27 1542.31±16.11j 50.46±5.07a 30.74±2.64l
CHI4-M28 734.26±17.62a 99.39±8.71hij 7.42±0.53ab
CHI4-M29 2089.70±18.76o 48.44±2.45a 43.20±1.78no
CHI4-M30 1146.60±16.08h 96.28±6.96ghi 11.94±0.69h
kDa to 120 kDa with little consistency (Wang and Chang 1997). A faint band appeared at 55 kDa may represent the contaminated protein or another form of chitinase. Similarly, B. circulans WL-12 was reported to secrete six distinct chiti-nases into the cultural supernatant when the bacterium was grown in a medium containing chitin as an inducer substrate. Chitinase A1 showed a strong affinity to chitin and was suggested to play a major role in the degradation of chitin in the chitinase system of B. circulans WL-12 (Watanabe et al. 1992). Further studies will focus on the purification of chitinase enzyme for their homogeneity and characterization with respect to isoelectric point, pH and temperature stability.
ACKNOWLEDGMENT
This study was carried out under the Nuclear R&D Pro-gram of the Ministry of Education, Science and Technology, Republic of Korea.
REFERENCES
Barrows-Broaddus J and Kerr TK. 1981. Inhibition of Fusarium moniliforme var. subglutinans, the causal agent of pitch canker, by the soil bacterium Arthrobacter sp. Can. J. Micro-biol. 27:20-27.
Cody RM, Davis ND, Lin J and Shaw D. 1990. Screening micro-organisms for chitin hydrolysis and production of ethanol
Deshpande MV. 1986. Enzymatic degradation of chitin and its biological applications. J. Sci. Ind. Res. 45:273-281. Elad Y, Chet I and Henis Y. 1982. Degradation of plant
patho-genic fungi by Trichoderma harzianum. Can. J. Microbiol. 28:719-725.
Gooday GW. 1990. The ecology of chitin degradation. Adv. Microbiol. Ecol. 11:387-430.
Gunaratna KR and Balasubramanian R. 1994. Partial purification and properties of extracellular chitinase produced by Acre-monium obclavatum, an antagonist to groundnut rust, Puc-cinia arachidis. World J. Microbiol. Biotechnol. 10:342-345.
Haran S, Schickler H, Pe’er S, Longemann S, Oppenheim A and Chet I. 1993. Increased constitutive chitinase activity in transformed Trichoderma harzianum. Biol. Control Theory Appl. Pest Manange. 3:101-108.
Hsu SC and Lockwood JL. 1975. Powdered chitin agar as a selective medium to enumeration of actinomycetes in water and soil. Appl. Microbiol. 29:422-426.
Kunz C, Ludwig A, Bertheau Y and Boller T. 1992. Evaluation of the antifungal activity of the purified chitinase I from the filamentous fungus Aphanocladium album. FEMS Microbi-ol. Lett. 90:105-109.
Laemmli UK. 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227:680-685.
Leah R, Tommerup H, Svendsn IB and Mundy J. 1991. Bio-chemical and molecular characterization of three barley seed proteins with antifungal properties. J. Biol. Chem. 266:1564-1573.
Lorito M, Di Pietro A, Hayes CK, Woo SL and Harman GE. 1993a. Antifungal, synergistic interaction between chitinoly-tic enzymes from Trichoderma harzianum and Enterobacter cloacae. Mol. Plant Pathol. 83:721-728.
Lorito M, Harman GE, Hayes GK, Broadway RM, Tronsmo A, Woo SL and Di Pitro A. 1993b. Chitinolytic enzymes pro-duced by Trichoderma harzianum: antifungal activity of purified endochitinase and chitobiosidase. Phytopathology, 83:302-307.
Mathivanan N, Kabilan V and Murugesan K. 1997. Production of chitinase by Fusarium chlamydosporum, a mycoparasite to groundnut rust, Puccinia arachidis. Indian J. Exp. Biol. 35:890-893.
Madi L, Katan T, Katan Y and Henis Y. 1997. Biological control of Sclerotium rolfsii and Verticillium dahliae by Talaromyces flavus is mediated by different mechanisms. Phytopathology, 87:1054-1060.
Muzzarelli RAA. 1999. Native, industrial and fossil chitins. pp. 1-6. In: Chitin and chitinases (Jolls P and Muzzarelli
50 kDa 40 kDa 60 kDa
Fig. 3. SDS-PAGE analysis of chitinase from CHI2 and CHI4.
RAA eds.), Basel, Switzerland: Birkh_user Verlag. Ordentlich A, Elad Y and Chet I. 1988. The role of chitinase of
Serratia marcescens in biocontrol of Sclerotium rolfsii. Phytopathology, 78:84-88.
Park SK, Lee HY and Kim KC. 1995. Role of chitinase produced by Chromobacterium violaceum in the suppression of Rhi-zoctonia damping-off. Korean J. Plant Pathol. 11:304-311. Patil RS, Ghormade V and Deshpande MV. 2000. Chitinolytic
enzymes: An exploration. Enzyme Microb. Technol. 26:473-483.
Roberts WK and Selitrennikoff CP. 1988. Plant and bacterial chitinases differ in antifungal activity. J. Gen. Microbiol. 134:169-176.
Shaikh SA and Deshpande MV. 1993. Chitinolytic enzymes: Their contribution to basic and applied research. World J. Microbiol. Biotechnol. 9:468-475.
Saksirirat W and Hoppe HH. 1991. Secretion of extracellular enzymes by Verticillium psalliotae Treschow and Verticilli-um lecanii (Zimm.) Viegas during growth on uredospores of the soybean rust fungus (Phakopsora pachyrhizi Syd.) in liquid cultures. J. Phytopathol. 131:161-173.
Sakuda S. 1996. Studies on the chitinase inhibitors. Allosami-dins, In: Muzzarelli RAA (ed.) Chitin enzymology, Vol. 2. Grottammare: Eur. Chitin Soc., pp 203-212.
Sakurada M, Morgavi DP, Komatani K, Tomita T and Onodera R. 1996. Purification and characteristics of cytosolic chiti-nase from Piromyces communis OTS1. FEMS Microbiol. Lett. 137:75-78.
Shapira R, Ordentlich A, Chet I and Oppenheim AB. 1989. Control of plant diseases by chitinases expressed from cloned DNA in Escherichia coli. Phytopathology 79:1246-1249. Thompson JD, Gibson TJ, PleWniak F, Jeanmougin F and
Higgins DG. 1997. The clustalx windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 24:4876-4882.
Ulhoa CJ and Peberdy JF. 1992. Purification and some proper-ties of the extracellular chitinase produced by Trichoderma harzianum. Enzyme Microb. Technol. 14:236-240. Vaidya RJ, Macmil SLA, Vyas PR and Chhatpar. 2003. The
novel method for isolating chitinolytic bacteria and its application in screening for hyperchitinase producing mutant of Alcaligenes xylosoxydans. Lett. Appl. Microbiol. 36:129-134.
Watanabe T, Oyanagi W, Suzuki K, Ohnishi K and Tanaka H. 1992. Structure of gene encoding chitinase D of Bacillus circulans WL-12 and possible homology of the enzyme to other prokaryotic chitinase and class III plant chitinases. J Bacteriol. 174:408-414.
Wang SL and Chang WT. 1997. Purification and characteriza-tion of two bifunccharacteriza-tional chitinases/lysozymes extracellularly produced by Pseudomonas aeruginosa K-187 in a shrimp and crab shell powder medium. Appl. Environ. Microbiol. 63:380-386.
Wiwat C, Siwayaprahm P and Bhumiratana A. 1999. Purification and characterization of chitinase from Bacillus circulans No.4.1. Curr. Microbiol. 39:134-140.
Yanai K, Takaya N, Kojima M, Horiuchi H, Ohta A and Takaki M. 1992. Purification of two chitinases from Rhizopus oligosporus and isolation and sequencing of the encoding genes. J. Bacteriol. 174:7398-7406.
Manuscript Received: February 26, 2010 Revision Accepted: March 12, 2010