INTRODUCTION
Soil bacteria with mineral phosphate solubilizing (MPS) ability could be efficiently used as biofertilizers in P-defi-cient soils. Phosphate solubilizing bacteria (PSB) exhibit several mechanisms including protein extrusion and secre-tion of low molecular weight organic acids (Rodriguez and Fraga 1999). Inorganic anions are also involved in phosphate solubilization but they are less effective (Kim et al. 1997). Mineral phosphate solubilization by organic acid may be due to the lowering of pH, chelation of cations and by
com-peting with P for adsorption sites in soils (Maliha et al. 2004). Production of gluconic acids and ketogluconic acid by PSB and their role in mineral phosphate solubilization was well established (Goldstein and Liu 1987; Gyaneshwar et al. 1999; Deubel et al. 2000).
Gluconic acid and ketogluconic acids are produced by Gram negative bacteria through the enzymes of direct oxi-dation pathway of glucose (DOPG) such as pyrroloquino-line quinone (PQQ) dependent glucose dehydrogenase (GDH) and gluconate dehydrogenase (GADH). Two types of GDH enzyme that differs in their substrate specificity and subcellular localization were identified. GDH type A is membrane bound enzyme while GDH type B is soluble in nature and has been demonstrated only in A. calcoaceticus
─ ─ 121 ──
Substrate Specificity of Glucose Dehydrogenase and
Carbon Source Utilization Pattern of Pantoea dispersa strain
P2 and Its Radiation Induced Mutants
Young-Keun Lee and Senthilkumar Murugesan*Radiation Application Research Division, Advanced Radiation Technology Institute, Korea Atomic Energy Research Institute, Jeongeup 580-185, Korea
Abstract-- Mineral phosphate solubilizing Pantoea dispersa strain P2 produced 5.5 mM and 42.6 mM of gluconic acid on 24 h and 72 h incubation, respectively. Strain P2 exhibited glucose de-hydrogenase (GDH) specific activity of 0.32 IU mg--1protein. We have studied the substrate
spe-cificity of GDH as well as carbon source utilization pattern of strain P2. GDH of strain P2 did not use ribose as substrate. Utilization of lactose with specific activity of 0.65 IU mg--1 protein
indi-cated that the enzyme belongs to GDH type B isozyme. Arabinose, galactose, ribose, sucrose and xylose did not induce the synthesis of GDH enzyme while mannose induced the synthesis of GDH with highest specific activity of 0.58 IU mg--1protein. Through radiation mutagenesis, the
sub-strate specificity of GDH was modified in order to utilize wide range of sugars available in root exudates. Ribose, originally not a substrate for GDH of strain P2 was utilized as substrate by mu-tants P2-M5 and P2-M2 with specific activity of 0.44 and 0.57 IU mg--1protein, respectively.
Spe-cific activity of GDH on the media containing lactose and galactose was also improved to 1.2 and 0.52 IU mg--1protein in P2-M5 and P2-M6 respectively. Based on the carbon source availability in
root exudate, the mutants can be selected and utilized as efficient biofertilizer under P-deficient soil conditions.
Key words : Glucose dehydrogenase, Pantoea dispersa, Substrate specificity
* Corresponding authors: Senthilkumar Murugesan, Tel. +82-63-570-3304, Fax. +82-63-570-3309, E-mail. [email protected]
(Cleton-Jansen et al. 1989). Rapid and efficient solubiliza-tion of P from rock phosphate by Pantoea agglomerans MMB051 was reported by Sulbaran et al. (2009). They also elucidated that a mechanism involving GA production and proton extrusion as a consequence of NH4++ uptake were responsible for phosphate solubilization.
In our laboratory, phosphate solubilizing potential of Pantoea strains and their mutant clones has been well established (Senthilkumar et al. 2009). Among the strains, Pantoea dispersa strain P2 was used in this study to deter-mine glucose dehydrogenase activity and its substrate spe-cificity. GDH enzyme also converts aldose sugars such as galactose, maltose and xylose to their corresponding aldo-nic acids, which can efficiently solubilize the mineral phos-phate. Versatile utilization of different sugars as carbon source for growth and substrate for GDH enzyme could have an added advantage over competitors that use limited carbon sources in natural environment. Mutant clones of strain P2 with altered MPS activity were also analysed for changes in substrate specificity of glucose dehydrogenase enzyme.
MATERIALS AND METHODS
1. Bacterial strains and their mutant clonesPantoea dispersa strain P2, originally isolated from the dry field soil (Senthilkumar et al. 2009) and their gamma radiation induced mutant clones were collected and stored at -80�C in nutrient broth with 25% glycerol. Bacterial strains were also maintained in nutrient agar (NA) and Pi-kovskaya (PVK) agar plates (PiPi-kovskaya 1948) under refri-gerated condition.
2. HPLC analysis of gluconic acid
Pantoea dispersa strain P2 was grown on PVK (Pikovs-kaya 1948) and national botanical research institute’s phos-phate solubilizing (NBRIP) liquid medium amended with 50 mM glucose (Nautiyal 1999). Samples were taken asep-tically at 24 h and 72 h of incubation at 28±2�C. Culture supernatants were prepared by centrifugation at 5,000 rpm for 10 min at 4�C and filtered through 0.22μm nylon filter. Gluconic acid produced by strain P2 was quantified by Agi-lent 1,200 high performance liquid chromatography (HPLC).
Gluconic acid was separated in RP-18 column with 0.1% phosphoric acid at a flow rate of 1 ml min-1and detected by
absorbance at 210 nm (Vikas et al. 2005).
3. Gluconic acid dehydrogenase (GDH) assay Strain P2 and its mutants were grown for 24 h on NBRIP liquid medium with 50 mM glucose or other carbon sources as listed in Table 1. The liquid medium was buffered with 100 mM Tris-HCl (pH 8.0). Cell pellets were derived by centrifugation of culture broth at 5,000 rpm, 4�C for 10 min, washed twice with 50 mM Tris-HCl (pH 8.75) and fol-lowed by suspension in known volume of the same buffer. This whole cell suspension was used as enzyme source in GDH assay carried out as described by Matsushita and Ame-yama (1982). One ml of reaction mixture contains 0.333μl of 50 mM Tris-HCl buffer (pH 8.75), 3.3μl of 6.7 mM 2,6-dichlorophenolindophenol (DCIP), 3.3μl of 20 mM phena-zine methosulfate (PMS), 4 mM sodium azide, glucose at final concentration of 100 mM and sample. The reaction was initiated by the addition of glucose. The extinction co-efficient of DCIP at pH 8.75 is taken as 15.1 mM-1. Protein
content of the sample was determined by using Bio-Rad protein assay kit as described by the manufacturers. Cells lysate prepared by sonication was used for total protein
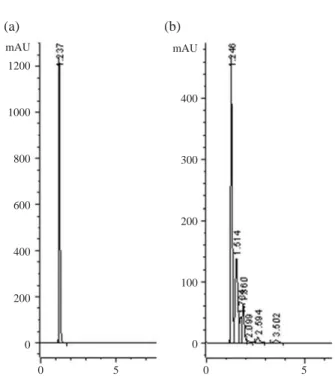
esti-Fig. 1. HPLC detection of gluconic acid (a) 100 mM standard
gluconic acid, (b) Gluconic acid in culture supernatant of
Pantoea dispersa strain P2. 1200 1000 800 600 400 200 0 mAU 400 300 200 100 0 mAU 0 5 0 5 (a) (b)
mation.
4. Substrate specificity and activity of GDH on different C sources
To study the utilization of different carbon sources, strain P2 and mutant clones were inoculated in NBRIP liquid me-dium with 50 mM of carbon source as listed in Table 1. The medium was buffered with 100 mM Tris-HCl (pH 8.0). Cell pellet from 24 h old liquid culture was used for GDH esti-mation as described above with 100 mM glucose as enzyme substrate. To determine the substrate specificity of GDH,
Pantoea strains and mutant clones were inoculated in
buf-fered NBRIP liquid medium with 50 mM of glucose as car-bon source. Cell pellet from 24 h old liquid culture were used for GDH estimation with 50 mM of different sugars as en-zyme substrate. Enen-zyme activity was expressed as IU mg-1
protein.
RESULTS AND DISCUSSION
1. Gluconic acid production
Among several plant growth promoting mechanisms ex-hibited by rhizobacteria, mineral phosphate solubilization plays a key role in the supply of phosphate in a soluble ionic form (Pi) by solubilizing the unavailable P from metal phos-phate complexes. Organic acid production by phosphos-phate solubilizing microorganisms (PSM) was linked to mineral phosphate solubilization (MPS) (Goldstein 1986). In most of the Gram negative bacteria, glucose is directly oxidized into gluconic acid. In our laboratory, production of gluconic acid by Pantoea dispersa strain P2 is confirmed by HPLC analysis. Gluconic acid was eluted with a retention time of 1.246 min, similar to that of pure standard gluconic acid. Strain P2 produced 5.5 mM and 42.6 mM of gluconic acid
on 24 h and 72 h incubation respectively on PVK broth (Table 1). Gluconic acid production was increased to 9.04 mM on 24 h incubation in buffered NBRIP liquid medium.
Previously, we developed mutants of P2 with enhanced as well as reduced mineral phosphate solubilizing activity by gamma radiation induced mutagenesis. In this study, selected mutants were analysed for acid production. P2-M1, radiation induced mutant of strain P2 with enhanced MPS activity acidified the culture supernatant to a pH of 3.81 and produced 10.04 mM of gluconic acid. In some cases, the pH value of culture supernatant and released P concentra-tions were negatively correlated (Banik and Dey 1983; Asea
et al. 1988; Kim et al. 1997). However, MPS activity of
Strain P2 and their mutants are directly correlated with acid production (Senthilkumar et al. 2009).
2. GDH enzyme activity
Membrane bound as well as soluble glucose dehydro-genase are involved in the direct oxidation of glucose in to Table 1. Gluconic acid production by Pantoea dispersa strain P2
and its mutants
Strain/mutant clone Gluconic acid production (mM)
P. dispersa strain P2 9.04 P2-M1 10.04 P2-M2 10.03 P2-M3 9.87 P2-M4 9.41 P2-M5 7.08 P2-M6 9.81
Table 2. Glucose dehydrogenase activity of radiation induced
mutants of Pantoea dispersa strain P2
Wild/mutant GDH activity Protein Specific activity
clones (IU) (mg L-1) (IU mg-1protein)
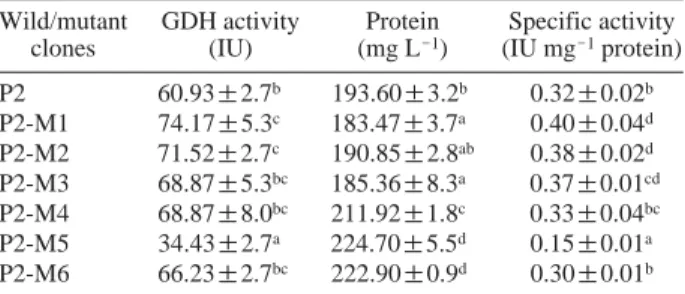
P2 60.93±2.7b 193.60±3.2b 0.32±0.02b P2-M1 74.17±5.3c 183.47±3.7a 0.40±0.04d P2-M2 71.52±2.7c 190.85±2.8ab 0.38±0.02d P2-M3 68.87±5.3bc 185.36±8.3a 0.37±0.01cd P2-M4 68.87±8.0bc 211.92±1.8c 0.33±0.04bc P2-M5 34.43±2.7a 224.70±5.5d 0.15±0.01a P2-M6 66.23±2.7bc 222.90±0.9d 0.30±0.01b
Treatment followed by same superscript letter do not differ significantly at p==0.05. The experiment was repeated thrice (n==3) with triplicates (r==3)
Table 3. Glucose dehydrogenase (GDH) activity of Pantoea
dis-persa strain P2 grown under different carbon sources
Sugar Protein GDH activity
(mg L-1) (IU mg-1protein) Arabinose 49.11±1.9b 0 Fructose 121.08±4.7f 0.23±0.01a Galactose 112.62±4.8e 0 Glucose 148.18±5.2h 0.32±0.02c Lactose 59.27±0.9c 0.28±0.02b Maltose 53.34±1.5b 0.20±0.05a Mannose 119.39±1.5f 0.58±0.02d Ribose 132.94±1.2g 0 Sucrose 39.80±2.2a 0 Xylose 64.35±1.5d 0
Treatment followed by same superscript letter do not differ significantly at p==0.05. The experiment was repeated thrice (n==3) with triplicates (r==3)
gluconic acid. Specific activity of GDH from strain P2 was determined with DCIP and PMS as electron acceptors. The strain P2 was grown for 20 h in buffered NBRIP liquid me-dium with 100 mM glucose as C-source. It exhibited the GDH specific activity of 0.32 IU mg-1protein. Although
the biomass production was lesser than that of the wild strain, P2-M1 showed higher enzyme activity (74.17 IU) with specific activity of 0.4 IU mg-1protein (Table 2).
3. Substrate specificity of GDH
Root exudates are the substantial carbon sources for rhizobacteria. Glucose is the main carbon source for PSM but other carbon sources can also be utilized (IIlmer and Schinner, 1992). Apart from glucose, root exudates contain many sugars such as arabinose, ribose, sucrose and fructose (Odunfa and Werner 1981; Kamilova et al. 2006). It has been suggested that rhizobacteria using different sugars as GDH substrate as well as carbon source could be very effec-tive under field conditions. We have studied the substrate specificity of GDH. Among the tested sugars, ribose was found to be a non-substrate of GDH produced by strain P2. Utilization of lactose as enzyme substrate with specific acti-vity of 0.65 IU mg-1protein indicated that the enzyme
be-longs to GDH type B isozyme.
Ability of strain P2 to utilize GDH enzyme substrate as sole carbon source was tested. Arabinose, lactose, maltose, sucrose and xylose were not utilized as efficiently as glu-cose which was indicated by total protein of cells grown with these sugars as sole carbon source (Table 2). Arabinose, galactose, ribose, sucrose and xylose did not induce the synthesis of GDH enzyme and reported zero specific acti-vity by the cells. Among the tested sugars, mannose
induc-ed the synthesis of GDH with highest specific activity of 0.58 IU mg-1protein.
It is possible to modify the substrate specificity of GDH by the mutagenesis. Norio et al. (2006) reported the narrowing down of substrate specificity of PQQGDH-B by biased mu-tation assembling method in order to develop glucose bio-sensors. In our case, mutant with broad GDH substrate spe-cificity and C source utilization was selected as suitable bio-fertilizing candidates under nutrient limiting conditions. Ribose, originally not a substrate for GDH of strain P2 was utilized as substrate by the mutants P2-M5 and P2-M2 with specific activity of 0.44 and 0.57 IU mg-1protein,
respec-tively (Table 4). Specific activity of GDH on lactose was improved to 1.2 and 0.8 IU mg-1protein in M5 and
P2-M4 mutants compared to 0.66 IU mg-1protein of the wild
type. Mutant P2-M1 recorded the higher specific activity on glucose (0.94 IU mg-1protein) than the wild type. Specific
activity of GDH on galactose was improved from 0.19 IU mg-1protein (wild type) to 0.52 IU mg-1protein in the
mu-tant P2-M6. Based on the carbon source availability in root exudates, the mutants can be selected and utilized as effi-cient biofertilizer under P- defieffi-cient soil conditions.
ACKNOWLEDGMENT
This study was carried out under the Nuclear R & D Program of the Ministry of Education, Science and Tech-nology, Republic of Korea.
REFERENCES
Asea PEA, Kucey RMN and Stewart JWB. 1988. Inorganic Table 4. Substrate specificity of GDH from Pantoea dispersa strain P2 and its mutants
Sugar GDH activity (IU mg
-1protein) P2 P2-M1 P2-M2 P2-M3 P2-M4 P2-M5 P2-M6 Arabinose 0 0 0.32±0.01c 0 0.17±0.02a 1.06±0.02g 0.07±0.01a Fructose 0.51±0.02b 0.55±0.01d 0.51±0.01d 0.49±0.01f 0.37±0.02b 0.54±0.02d 0.69±0.02g Galactose 0.19±0.03a 0.32±0.02b 0.25±0.01b 0.26±0.01c 0.41±0.02b 0.27±0.02ab 0.52±0.01e Glucose 0.69±0.03e 0.95±0.01g 0.80±0.01g 0.75±0.02h 0.78±0.02e 0.30±0.02b 0.61±0.01f Lactose 0.66±0.03de 0.46±0.02c 0.51±0.01d 0.34±0.02d 0.85±0.03f 1.20±0.01h 0.53±0.02e Maltose 0.79±0.02f 0.48±0.02c 0.49±0.02d 0.61±0.01g 0.53±0.03c 0.64±0.02e 0.72±0.01h Mannose 0.98±0.02g 0.80±0.02f 0.76±0.01f 0.80±0.02i 0.86±0.02f 0.71±0.03f 0.48±0.01d Ribose 0 0.05±0.01a 0.57±0.01e 0.19±0.02b 0 0.44±0.02c 0.08±0.01a Sucrose 0.62±0.02d 0 0.01±0.01a 0.07±0.02a 0.49±0.03c 0 0.32±0.01c Xylose 0.56±0.02c 0.59±0.02e 0.90±0.03h 0.44±0.01e 0.73±0.03d 0.21±0.01a 0.19±0.01b
phosphate solubilization by two Penicillium species in solution culture and soil. Soil Biol. Biochem. 20:459-464. Banik S and Dey BK. 1983. Alluvial soil microorganisms
capable of utilizing insoluble aluminium phosphate as a source of phosphorous. Zentralblatt Mikrobiol. 138:437-442.
Cleton-Jansen AM, Goosen N, Vink K and Putte P. 1989. Cloning, characterization and DNA sequencing of the gene encoding the Mr 50,000 quinoprotein glucose dehydroge-nase from Acinetobacter calcoaceticus. Mol. Gen. Genet.
217:430-436.
Deubel A, Gransee A and Merbach W. 2000. Transformation of organic rhizo-depositions by rhizosphere bacteria and its influence on the availability of tertiary calcium phosphate.
J. Plant Nutr. Soil Sci. 163:387-392.
Goldstein AH. 1986. Bacterial solubilization of mineral phos-phates: Historic perspective and future prospects. Am. J.
Altern. Agric. 1:57-65.
Goldstein AH and Liu ST. 1987. Molecular cloning and regu-lation of a mineral phosphate solubilizing gene from
Erwi-nia herbicola. Biotechnol. 5:72-74.
Gyaneshwar P, Parekh LJ, Archana G, Poole PS, Collins MD, Hutson RA and Nareshkumar G. 1999. Involvement of a phosphate starvation inducible glucose dehydrogenase in soil phosphate solubilization by Enterobacter asburiae.
FEMS Microbiol. Lett. 171:223-229.
Illmer P and Schinner F. 1992. Solubilization of inorganic phosphorous by microorganisms isolated from forest soils.
Soil Biol. Biochem. 24:389-395.
Kamilova F, Kravchenko LV, Shaposhnikov AI, Azarova T, Makarova N and Lugtenberg B. 2006. Organic acids, sugars and L-tryptophane in exudates of vegetables growing on stonewool and their effects on activities of rhizosphere bacteria. Mol. Plant Microbe Interact. 19:250-256. Kim KY, Jordan D and McDonald GA. 1997. Solubilization of
hydroxyapatite by Enterobacter agglomerans and cloned
Escherichia coli in culture medium. Biology and Fertility of Soils. 24:347-352.
Maliha R, Samina K, Najma A, Sadia A and Farooq L. 2004. Organic acids production and phosphate solubilization by
phosphate solubilizing microorganisms (PSM) under in vitro conditions. Pak. J. Biol. Sci. 7:187-196.
Matsushita K and Ameyama M. 1982. D-glucose dehydroge-nase from Pseudomonas fluorescens, membrane bound.
Methods Enzymol. 89:149-154.
Nautiyal CS. 1999. An efficient microbiological growth medium for screening phosphate solubilizing microorganisms. FEMS
Microbiol. Lett. 170:265-270.
Norio H, Akitoshi S, Yukiko N, Masaaki S, Takuyo A, Moto-wo N, Yuzuru H and Yasuhiko S. 2006. Modified substrate specificity of pyrroloquinoline quinine glucose dehydro-genase by biased mutation assembling with optimized ami-no acid substitution. Appl. Microbiol. Biotechami-nol. 73:607-617.
Odunfa SA and Werner D. 1981. Root exudates in relation to growth and nitrogenous activity of Rhizobium japonicum.
Z. Allg. Mikrobiol. 21:601-606.
Pikovskaya RI. 1948. Mobilization of phosphorus in soil con-nection with the vital activity of some microbial species.
Microbiologiya 17:362-370.
Rodriquez H and Fraga R. 1999. Phosphate solubilizing bacte-ria and their role in plant growth promotion. Biotechnol.
Adv. 17:319-339.
Senthilkumar M, Lee YK and Kim JH. 2009. Mineral phos-phate solubilization by wild type and radiation induced mutants of Pantoea dispersa and Pantoea terrea. J.
Radia-tion Industry 3:39-45.
Sulbaran M, Perez E, Ball MM, Bahsas A and Yarzabal LA. 2008. Characterization of the mineral phosphate solubili-zing activity of Pantoea agglomerans MMB051 isolated from an iron rich soil in south-eastern Venezuela (Bolivar State). Curr. Microbiol. 58:378-383.
Vikas S, Vikas K, Archana G and Nareshkumar G. 2005. Sub-strate specificity of glucose dehydrogenase (GDH) of
Enter-obacter asburiae PSI3 and rock phosphate solubilization
with GDH substrates as C sources. Can. J. Microbiol. 51: 477- 482.
Manuscript Received: June 18, 2009 Revision Accepted: June 22, 2009