A thesis submitted to the Ulsan National Institute of Science and Technology in partial fulfillment of the requirements for the degree of Master of Science. While Kramers theory has been successfully applied to barrier crossing reactions occurring in the high friction limit, challenges arise when applying this theory to the complex environment of protein interiors. To address these challenges, we study the rearrangement reaction occurring within the chromophore pocket of green fluorescent protein (GFP) using single-molecule 2D fluorescence lifetime correlation spectroscopy (sm-2D-FLCS).
We compare the solvent-viscosity dependence of the internal reaction with Kramers' rate theory and the non-Markovian expansion (Grote-Hyne theory). We report strong deviations from Kramers' behavior showing a power-law dependence of reaction time with solvent viscosity , med. The results are consistent with 'internal friction' previously observed in protein folding experiments, suggesting a universal description of the friction coefficient for reaction processes occurring in the protein interior.
List of Tables
List of Abbreviations
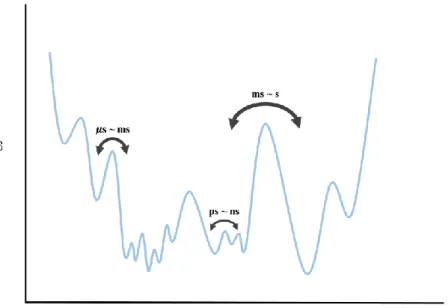
Interestingly, despite the difference in time scales of the movements studied, both studies observed the same power law scaling of transition time with friction, 𝜏 ∝𝛼, with 𝛼 = 0.5. This is in contrast to the well-known Kramers scale coefficient of 𝛼 = 1. Here is the observed transition time and is the friction coefficient of the bath. These observations led to the introduction of 'internal friction', a term used to describe the apparent friction observed in protein movements.
-24) Despite the consistency of the observation of internal friction, the microscopic origin of the phenomenon is not well understood. Furthermore, the technique offers one of the few possible approaches to directly characterize the dynamics occurring under equilibrium conditions. Single-molecule fluorescence resonance energy transfer (sm-FRET) is perhaps the most widely used technique for studying the detailed kinetic information of biological macromolecules.
This energy transfer process depends critically on the distance between the donor and acceptor chromophore. Therefore, protein movements that lead to a change in the distance between the two probe chromophores can be measured through sm-FRET experiments. While this approach has provided valuable information for a number of experimental systems, there are several drawbacks to the technique.
Recent years have seen significant advances in single-molecule two-dimensional fluorescence lifetime correlation spectroscopy (sm-2D-FLCS). This approach uses small changes in fluorescence lifetime to identify distinct conformational states of a biological macromolecule. Since the technique monitors small changes in fluorescence lifetime, a single probe chromophore is capable of resolving many degrees of freedom, corresponding to the ability to monitor the exchange between many distinct conformations.
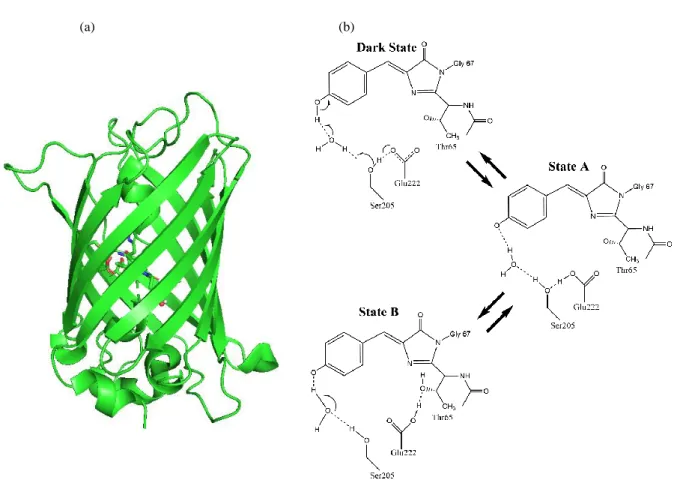
Green Fluorescent Proteins
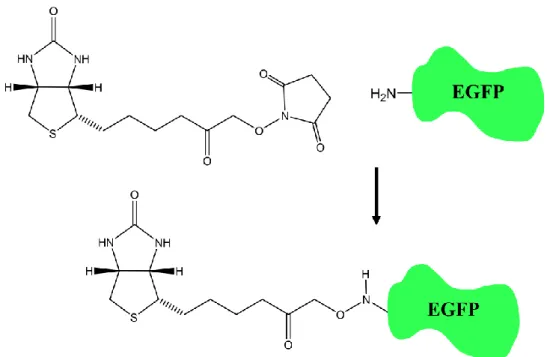
This work aims to characterize the internal reaction dynamics that occur in the eGFP chromophore pocket, with a particular interest in understanding 1) the coupling of the internal reaction coordinate to the external bath, and 2) the role of site-specific mutations in modulate the free energy surface of the chromophore pocket. To create an immobilized, sparsely localized, single-molecule microscopy sample, we use biotin-streptavidin coupling. Thus, drop 200 biotin-labeled protein solutions onto the streptavidin-coated solution and then incubate for 1.5 hours in a dark condition to prevent the photoactivation of eGFP.
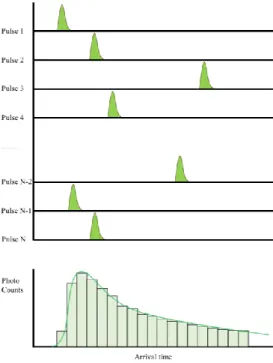
Single-molecule fluorescence lifetime data is collected using an inverted confocal microscope equipped with time-correlated single-photon counting (TCSPC) detector. TCSPC is one of widely used for fluorescence correlation spectroscopy (FCS) fields. REF)TCSPC resolves the time delay between the arrival of an excitation pulse at the sample and the arrival time of an emitted photon at the detector. Fitting the emission delay histogram to a decay function (typically exponential or bi-exponential) allows for the extraction of the fluorescence lifetime of the molecule of interest.
The excitation and detection wavelengths are chosen according to the absorption and emission spectra of the system of interest. To use the fluorescence lifetime, two independent time scales are needed: (i) the lifetime, the measurement of the time gap between the photon arrival and the excitation pulse, and (ii) the FCS-scale measurement time between the photon arrival and the onset of the experiment. The 𝑝𝑗(𝑘) decay patterns can be obtained either experimentally or the mathematical decomposition of the total intensity decay.
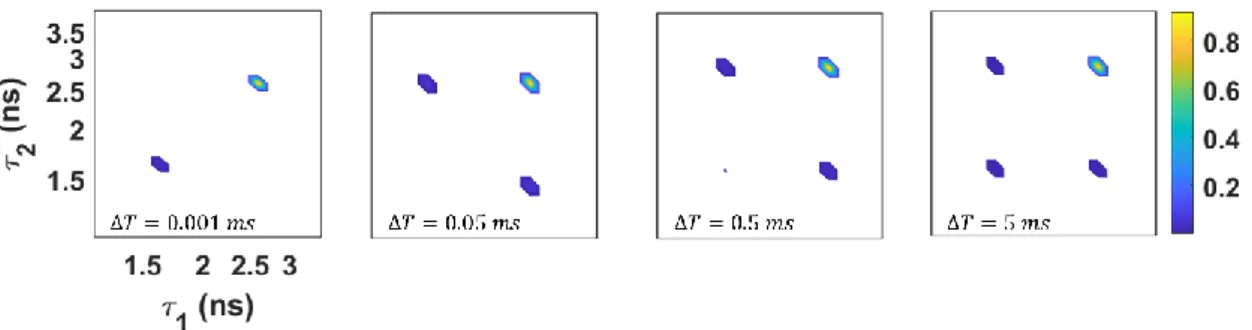
A histogram including multiple values is separated using the filter value 𝑓𝑘(𝑘) calculated by matrix algebra. 59-61) This value separates the contribution of each fluorescence state with a weight depending on the combining number of the TCSPC photon channel 𝑗. The aim of the technique is to detect conformational heterogeneity and dynamics in the system to realize their functions. In the 2D-FLCS spectrum, the population of unique conformations of the system appears as peaks along the diagonal.
Measuring a set of 2D-FLCS spectra allows the dynamic exchange occurring in the system to be directly time-resolved. In the histogram map, each photon detected using TCSPC is labeled with a "microtime (tx)", and each histogram map produced is divided by a "macrotime (ΔT). The macrotime (ΔT) of an experiment varies systematically from s ~ s time scales, while when the microtime (tx) is in the ps~ns range.
Photon pairs were recorded based on delayed macrotime (ΔT) from detected photon and within a time window (ΔΔT). Recorded photon pairs as indicated in histogram with micro time, 'tx' means a micro time of photon's arrival dividing in sequence, x-axis first detected is baseline photon and y-axis is another photon's arrival micro time within time window (∆ ∆𝑇) indicating in the y-axis.
Result
To identify multiple states present in the system, we first measure the 1D fluorescence relaxation spectrum. The spectrum also reveals the relative population of the two conformations as 0.3 and 0.7 for state A and state B, respectively (Table 4.1). Exchange between state A and state B is revealed through sm-2D-FLCS spectroscopy, which can directly resolve conformational exchange on time scales from 1 s to 1 s.
In the 2D spectrum, distinct conformations of the protein are observed as peaks along the diagonal of the spectrum. The exchange between the two modes can be monitored directly by plotting the peak amplitude of the cross peaks as a function of time. Fitting the exponential growth of the cross-peak amplitude allows the exchange time of the A-B and B-A transitions to be extracted.
Kramers theory is based on transition state theory involving particle diffusion on a free energy surface. In large-scale motion dictated by Brownian walks along a free energy profile, the rate constant 𝑘𝑜𝑏𝑠 depends on −1, where is the bath friction. Assuming that is given by Stokes' law, we can write the Kramers rate expression in the high friction regime as, .
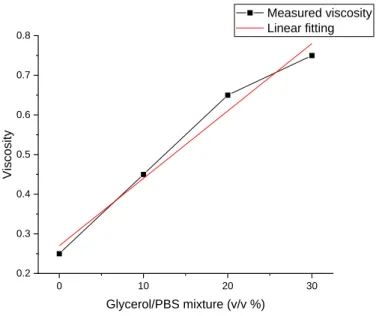
Although Kramers' theory has been widely applied to condensed-phase reactions, it is often experimentally observed that processes including enzyme kinetics (66-68), conformational fluctuations (69, 70), and protein folding (71-73) follow a modified rate expression , . This fractional value explained by the. internal friction' that reduces the effective viscosity of the bath. To observe the scaling of kobs with bath viscosity, we measure the chemical exchange occurring in eGFP as a function of water-glycerol mixtures.
So, in light of these data, the reaction rate decreases with an increase in solvent viscosity as Kramers expected, regardless of the reaction type. To derive the protein-bath coupling, we plot the observed reaction time rxn = kobs-1 as a function of the solvent viscosity (Figure 4.6). This power-law scaling is consistent with 'internal friction' observed in protein folding (71-73) and conformational fluctuation (69, 70) dynamics.
Discussion
However, at high viscosity, the relaxation time increases in the energy landscape pools, where it is independent of level 2 of the solvent viscosity. (77, 78). As the reaction time is determined by the local fluctuations, a deviation from Kramers' theory is expected in the high viscosity regime. It is an extension of Kramers' theory, based on the Markovian to non-Markovian system of Grote and Hynes.
In this experiment we confirmed this system, the link between eGFP chromophore and bath system. This coupling is also expressed by a power law dependence of the reaction time with a solvent viscosity scale of 0.5. For this experiment, we confirmed isolated photophysical chromophore motions and their weak bath coupling based on a non-Markovian system.
And explaining this with a hierarchical free energy landscape for protein provides an experimental agreement with a theoretical model.
Conclusion
Soranno et al., Quantification of internal friction in unfolded and intrinsically disordered proteins with single molecule spectroscopy. Ha et al., Probing the interaction between two single molecules: fluorescence resonance energy transfer between a single donor and a single acceptor. Eaton, Characterization of the unfolded states of proteins using single-molecule FRET spectroscopy and molecular simulations.
King, Fast and robust two-dimensional inverse Laplace transform of single-molecule fluorescence lifetime data. Jones, Crystal structure of enhanced green fluorescent protein to 1.35 Å resolution reveals alternative conformations for Glu222. Zimmer, Photophysics and dihedral freedom of the chromophore in yellow, blue, and green fluorescent protein.
Schmitt et al., eGFP-pHsens as a highly sensitive fluorophore for cellular pH determination by fluorescence lifetime imaging microscopy (FLIM). Kay, Viscosity-dependent kinetics of protein conformational exchange: microviscosity effects and the need for a small viscogen. Marques et al., Subjective versus objective, polymer cage-based selective removal of caries tissue: 1-year interim analysis of a randomized clinical trial.
I would first like to thank my thesis supervisor, Professor John Thomas King, for his help and support during my thesis. I am also grateful to Professor Oh-Hoon Kwon and Professor Hyuk-Kyu Pak for being part of the supervisory committee and for constant encouragement.