317
목 적 :티타늄 입자에 의한c-fos 유전자 발현에 관련된 세포내 신호전달을 이해하기 위해, Rac, cytosolic phospholipase A2, eicosanoids (e.g., leukotriene B4 and prostaglandin E2) 그리고, c-fos의 관련에 대해 분석 하였다.
대상 및 방법 : Rat-2 섬유 아세포에 티타늄 입자를 첨가 하여c-fos serum response element의 활성도 변화를 조사 하였다.
c-fos serum response element의 측정을 위해 luciferase reporter system을 이용하였으며, luciferase는 scintillation spec- trophotometer로 활성도를 측정하였다. 다음, c-fos로의 활성화 경로 내에서 Rac의 연관성과 그 하류 전달체로서의 eicosanoids 합성 역할을 분석하였다.
결 과 :티타늄 입자에 의해c-fos serum response element의 활성화가 일어나는 것이 확인되었으며, 또한 RacN17, cytoso- lic phospholipase A2, cyclooxygenase, 5-lipoxygenase의 억제제 투여에 의해 선택적으로 억제 되었다. 또한, eicosanoids의 합성이 Rac 의존성 방식으로 티타늄 마모편에 의해 증가 하였다.
결 론 :티타늄에 의한 신호 전달계에서는 G-단백의 일종인 Rac과 eicosanoids 합성이 필수적인 역할을 하고 있다. 또한 이러한 Rac-cytosolic phospholipase A2-eicosanoids-c-fos 신호전달계가 인공 삽입물 주위의 골 용해에서, 티타늄 입자에 의한 eicosanoids합성을 일으키는 기작으로 관여할 가능성을 생각할 수 있다.
색인 단어 : 섬유 아세포, 티타늄, Rac, Eicosanoid, c-fos
317
티타늄 입자에 의한 G-단백 Rac과 Eicosanoids의 활성화 경로
이상수∙허재영∙우창훈*∙김재홍*∙장준동**
포항성모병원 정형외과, 고려대학교 생명공학원*, 한림의대 정형외과학교실**
서 론
세포는 외부 환경의 변화나 자극에 대하여, 신호 전이(signal transduction)라는 세포 내 기작을 통해 빠르게 반응한다21). 세 포자극물질(agonist)은 세포를 통과하지 못하기 때문에 세포막 에 존재하는 수용체와 결합한 후 세포내부에 있는 신호전달체계 를 활성화 시키기 위한 이차신호전달물질(second messenger) 을 생성한다. 이차신호전달물질에는 Ca2+, cyclic AMP, cyclic GMP, inositol, 1,4,5-triphosphate (IP3), diacylglyc- erol, phosphatidic acid 등 다양한 종류가 알려져 있다. 최근에 는 nitric oxide (NO)나 reactive oxygen species (ROS) 등 의 활성물질도 세포의 신호전달에 중요하게 작용하는 것으로 알 려지고 있다21,28). 인공 관절 소재로부터 형성되는 마모편 입자 들도 역시 삽입물 주위의 여러 가지 종류의 세포들을 자극하게 된다. 여기에는 섬유 아세포, 파골 세포, 골 모세포, 대식 세포 들이 포함된다21,24,26). 특히 삽입물 주위의 섬유 아세포에서는 정 상의 활막 조직이나 피부의 섬유 아세포 보다 metalloproteinase 들의 농도가 증가 되어 있다30).
c-fos는 최조기 유전자(immediate early gene)의 일종이며, 성장인자, cytokine, 환경적 스트레스 같은 여러 가지 세포외적 신호에 의해 즉각적으로 발현이 된다. c-fos 유전자의 지속적 발현은 특정 세포에 제한되어 있지만, 거의 모든 세포에서 다양 한 자극에 대하여 일시적으로 발현이 될 수가 있다. 특히, c-fos 는 세포의 분할과 분화 조절의 중요한 기능을 한다고 할 수 있 다. c-fos에 존재하는 Serum Response Element (SRE)는 세 포외적 자극의 일차적 핵 표적(nuclear target)으로 알려져 있 으며, c-fos 유전자 전사의 급속유도에 관여하는 것으로 알려져
있다18,20,25). 본 연구에서는 섬유 아세포에서 인공 관절 소재인
티타늄 입자를 자극물질로 하여, G-단백 중 Rho family GTPase의 일종인 Rac과 cytosolic phospholipase A2(cPLA2), eicosanoids, 그리고 c-fos SRE를 중심으로 한 신호 전달계를 분석 하였다.
재료 및 방법
본 연구의 개요는 아래와 같다. 첫째, 티타늄이c-fos SRE를 활성화시키는 여부를 확인하였다. 이를 위해, Rat-2 섬유 아세 포를 luciferase 부호 서열이 결합된c-fos minimal promoter의 상부에 c-fos SRE oligonucleotides가 삽입된 지시 plasmid인 pSRE-Luc (3 g)와 과도이입 하였다14)(Fig. 2A). 티타늄 입
317 317 통신저자 : 장 준 동
서울특별시 영등포구 영등포동 94-200
한림대학교 의과대학 부속 한강성심병원 정형외과 TEL: 02-2639-5301∙FAX: 02-2631-9337
E-mail: [email protected]
자 처리 전, 이입된 Rat-2 세포를 36시간동안 0.5% 우태아혈청 을 포함한 Dulbecco’s modified eagle’s medium (DMEM)에 서 배양하여 혈청 기아(serum starvation)처리 하였다. c-fos SRE 활성도는 luciferase 활성도를 측정하여 분석 하였다.
둘째, c-fos SRE 활성화를 위한 신호계는, Elk-1p62TCF의존 성 신호 경로 뿐만 아니라, 비의존성 신호 경로계가 존재 한다
2,4,12,27). 따라서 Elk-1p62TCF가 결손된 변이형 oligonucleotide
(AGG to TGT)를 포함하는 pSREmt-Luc을 사용하여 티타 늄 입자로 유발되는c-fos SRE 활성화에는 어떠한 신호 전달 계가 작용하는지 분석하였다14). 또한 MEK 억제제와 MEK1- dominant negative (DN) mutant를 사용하여 Ras-mitogen activated protein (MAP) kinase 경로의 관련을 확인하였다.
셋째, 티타늄 입자의 c-fos SRE로의 신호계에서 G-단백중 Rho family의 small GTPase인 Rac이 관여할 가능성에 대해 분석하였다. 이를 위하여 pSRE-Luc (3 g)을 대조군인 pEXV와 Rac1의 우성결손변이형으로 부호화한 pEXV- RacN17으로 공동이입 하였다. 또한 Rat-2와 Rat2-RacN17에 서 SRE-luciferase 활성도를 비교하여 다시 분석 하였다.
넷째, 일반적으로, Rac-c-fos SRE의 신호 전달계에서 cPLA2가 Rac의 중요한 하류 전달체로서 기여한다고 알려져 있
다15,16). 그러므로, 티타늄-c-fos SRE 신호계에서 cPLA2가 하
류전달체로 관여할 가능성을 생각할 수 있다. 이를 규명하기 위 해서 cPLA2, 5-lipoxygenase (5-Lo), cycloxygenase (COX) 에 대한 억제제들을 함께 투여하여 분석하였다.
다섯째, eicosanoids의 합성이 유발되는 기전을 규명하기 위
해, 티타늄 입자에 의한 leukotriene B4 (LTB4)와 prosta- glandin E2 (PGE2)의 세포내 변화를 측정하였다. 또한 합성 유 발 경로가 Rac-cPLA2에 의존하는지 알기 위해 Rat-2와 Rat2- RacN17에서c-fos SRE-luciferase 활성도를 비교하였다.
1. Chemicals and Plasmids
Commercially pure Titanium 입자(#000681, 280 mg/0.2 mL, 1-3 m; Alpha chemicals, USA)를 사용하여 멸균된 phosphate buffered saline (PBS; pH7.2)에 저장 하였다. 우 태아혈청(fetal bovine serum)과 DMEM은 Gibco-BRL사의 제품을, Indomethacine, MAFP, MK886, Aspirin, NS-398, PD098059, Wortmannin은 Sigma chemical사와 Calbiochem.
사의 제품을 사용 하였다. 다른 모든 시약들도 분자 생물학적 등급내지는 그 이상의 표준화된 것을 사용하였다. pSRE-Luc은 pFos-lcf에서 분리하여, -53에서 +45의 서열을 포함하는c-fos 촉진자(promoter)의 -53 위치에 삽입하였다(Fig. 2A). c-fos 촉진 유전자에는 luciferase 유전자가 함께 결합되었다14). 야생 형(pSREwt-Luc)과 변이형(pSREmt-Luc) SRE oligonu- cleotide의 염기 서열은 각각, AGGATGTCCATATTAG- GACATCT와 tGtATTCCATATTAGGACATCT이다.
PEXV, pEXV-RacN17 plasmids는 Dr. A. Hall (London, UK)이 제공하였다. 각 실험들은 모두 두 개의 독립된 격리 세 트에서 각 3회 시행하였고, 결과들의 평균 값을 구하였다.
Relative Luciferase Activity
2
1
0
0 0.05 0.075 0.1 0.25 0.5
Relative Luciferase Activity
2
1
0
0 1 1.5 2 2.5 3 4
Fig. 1.Titanium-particles activate c-fos SRE in a dose and time-dependent manner. After transient transfections with wild-type pSREwt- Luc (3 g) plasmids, Rat-2 cells were serum starved in DMEM/0.5% FBS for 36 h before adding Titanium-particles. (A) Dose-dependent response of Titanium-particles on c-fos SRE activation. Serum starved cells were treated with various amounts of Titanium-particles (0.0, 0.05, 0.075, 0.1, 0.25, 0.5 mg/mL) for 3 h. (B) Time-dependent response of Titanium-particles (0.1 mg/mL) on c-fos SRE activation.
Serum-starved Rat-2 cells were treated with Titanium-particles for the lengths of time indicated. Luciferase activities were measured and normalized to the co-transfected -galactosidase activities. Values are representative of triple transfections.
HOURS
Titanium-particle (mg/mL) A B
2.
세포 배양, DNA
이입과Luciferase
분석Rat-2 섬유 아세포주는 American Type Culture Collection (CRL 1764)에서 입수 하였다. 세포는 non-essential amino acids 0.1 mM (Gibco-BRL, Gaithersburg, MD, USA)과 10%의 우태아혈청, 50 units/mL의 penicillin, 50 g/mL의 streptomycin을 포함한 DMEM에서 5% 이산화탄소, 95% 공 기의 비율(v/v)과 37℃ 온도 조건으로 배양하였다. Rat2- RacN17 안정세포주는 Rac1의 우성결손변이형(dominant neg- ative mutant)인 RacN17의 발현형을 가지며, pSV-Neo� plasmid를 pEXV-myc-RacN17 plasmid와 공동이입(co- transfection)한 후 2-3주간 G418 항생제(400 g/mL)를 투여 하여 분리 하였다. RacN17 단백의 발현은 anti-myc 항원결정
기(epitope)항체를 이용한 Western blot hybridization으로 확 인하였다. 과도이입(transient transfection)은 100 mL 배양접 시 당 5×105개의 세포를 분주한 후 인산화 칼슘(calcium phosphate)을 첨가하여 24시간 동안 이루어졌다. DNA 침전물 은 배양접시 당 20 g의 DNA를 사용하여 만들어 졌으며, 사 용된 plasmid의 양은 reporter gene (e.g. pSRE-Luc)은 3 g 이며 small GTPase 발현형 plasmid (e.g. pEXV-RacV12)는 5 g이다.
유전자 이입시의 오차를 보정하기 위해, 모든 clone은 1 g의 pCMV- GAL을 이입 하였다. pCMV- GAL은 E.coli - galactosidase (lac Z) structural gene이 CMV 촉진자(pro- moter)의 전사 제어 조절하에 있는 진핵 발현 벡터(eukaryotic expression vector)이다. 인산화 칼슘으로 DNA 침전물 세포를 PBS로 2회 세척한 후 0.5% 우태아혈청을 포함하는 신선 DMEM에서 36시간 동안 배양하였다. 각 배양접시의 세포를 우 태아혈청으로 2회 세척하였고, 0.2 mL의 용해액[0.2 M Tris (pH7.6)+0.1% Triton X-100]으로 용해하여, 채집하였다. 상 층액의 luciferase와 -galactosidase 활성도를 분석하였다.
Luciferase의 활성도는 10 L의 추출물을 사용하여, tritium channel을 사용한 섬광 분광계(Beckman liquid scintillation spectrometer)로 계산 하였다.
Relative Luciferase Activity
7
6
5
4
3
2
1
0
Control Titanium EGF LPA
Relative Luciferase Activity
7
6
5
4
3
2
1
0
Control Titanium EGF LPA
Fig. 2.Elk-1/p62TCF-independent pathway to c-fos SRE by Titanium-particles. (A) Diagram of the pSRE-luciferase reporter gene plasmid.
The structures of the constructs containing the wild-type or mutant SRE oligonucleotide sequences (23-mers) that are inserted into the - 53 position of the truncated c-fos promoter which is fused to the luciferase gene are shown. The methylation interference pattern for the SRF ternary complex with Elk1/p62TCFis indicated by solid circles. The mutant SRE has two point mutations (AGG to TGT) that abolish the SRF-Elk1/p62TCFformation in the Elk/p62TCF-binding region. (B) Elk-1/p62TCF-independent pathway of Titanium to c-fos SRE. Rat-2 cell cultures that were not confluent only were transiently transfected with 3 g of pSREwt-Luc plasmids and cultured for 36 h in DMEM con- taining 0.5%FBS. Titanium-particles (0.1 mg/mL) was treated for 3 h before harvest, and the luciferase and -galactosidase activities were measured as described in the Materials and Methods section. EGF (50 ng/mL) and LPA (2 m) were treated for 2 h before the cell harvest. Values are representative of triple transfections. (C) No inhibition of Titanium-induced SRE activation by pCGN1-MEK1/DN. A reporter gene plasmid, pSRE-Luc (3 g) was transiently co-transfected with 5 g of pCGN1-MEK1/DN, which is a dominant negative MEK1 expression plasmid as described in the Materials and Methods. Transfected cells were serum-deprived in DMEM containing 0.5%
FBS for 36 h before treatment with the Titanium-particles (0.1 mg/mL), EGF (50 ng/mL), LPA (2 M). After the treatment, the luciferase activities were measured and normalized to the co-transfected -galactosidase activities. Values are representative of triple transfec- tions.
insert
Luciferase Gene
SRE-wt AGGATGTCCATATTAGGACATCT SRE-mt tGtATGTCCATATTAGGACATCT
-53 TATA +1 +45
A
C B
pSRE-Luc pSREmt-Luc
pCGN1 MEK1/DN
3. LTB
4 와PGE
2의 측정Rat-2 또는 Rat-2RacN17Asp17 세포를 60 mm 배양접시에 분주하여(3×105 ea/well), 10% 우태아혈청이 첨가된 DMEM 에서 24시간 배양하였다. 이후 24시간 동안 배양매체를 0.5%
우태아혈청을 포함하는 DMEM으로 교환하였고, 지정된 시간대 로 0.1 mg/mL의 티타늄 마모편 입자로 처리하였다. 세포내 LTB4의 양을 측정하기 위해서, 평판을 저온 PBS로 2회 세척 후, 4배 용적의 무수에탄올과 혼합하여, 4℃에서 30분간 방치하 였다. 발생한 침전물을 4℃에서 30분간 10,000 rpm으로 원심분 리하여 제거한 후, LTB4을 포함한 ethanolic 상층액을 C2 reverse phase column (Amersham Pharmacia Biotech, RPN 1903)을 통해 채집하였다. 진공에서 ethanol을 증발시킨 후, 표본을 assay buffer 내에서 재구성하여, -50℃의 argon 분 석완충액에 저장한 후 specific ELISA kit (Amersham Phar- macia Biotech, RPN 222)로 분석하였다. 결과를 LTB4 농도 로그 함수 표준곡선에 의해 계산하였다. PGE2는, Rat-2 또는 Rat-2RacN17Asp17 세포를 standard 96-well microtitre plate 에 분주 시킨후(1×104 ea/well), LTB4와 같은 방법으로 처리 하여 측정 하였다. LTB4와 PGE2 분석의 통계학적 유의성은 각 각 ANOVA test (p<0.05)와 unpaired t-test (p<0.01)로 평가 하였다.
결 과
1.
티타늄 입자의 핵 표적 서열로서의c-fos SRE
티타늄의 신호 전달계 분석의 첫 단계로, 티타늄이 c-fos SRE를 활성화시키는지에 대하여 실험하였다. 티타늄 입자의 첨 가에 의해 luciferase 활성도가 증가 하였다(Fig. 1). 티타늄 입 자(0.1 mg/mL)의 첨가 3시간 후 luciferase 활성도가 2.2배로 증가 하였으며, 그 후 다시 감소하였다. 티타늄 입자는 0.1 mg/mL의 농도에서 최대치의 luciferase 활성도가 관찰 되었다.
한편c-fos SRE가 제거된 vector인 pO-Luc으로 과도이입 한 Rat-2 세포주에서는, 티타늄 입자의 첨가에 의해 luciferase의 활성이 증가되지 않았다. 이 결과들은 Rat-2 섬유아세포에서의 c-fos SRE는 티타늄 입자에 의한 핵 표적 서열 중의 하나라는 것을 의미한다.
2.
티타늄 신호계와Ras/MAP Kinase
경로티타늄 입자로 유발되는 c-fos SRE 활성화에 Ras/MAP Kinase 경로가 작용하는지 알기 위해 Elk-1p62TCF가 결손된 변 이형 oligonucleotide (AGG to TGT)를 포함하는 pSREmt- Luc를 이입하여 실험하였다14). 그 결과 티타늄 입자는
pSREmt-Luc과 pSREwt-Luc을 동일한 정도로 자극하였다 (Fig. 2B). 유사하게, Elk-1p62TCF 비의존성으로c-fos SRE를 활성화시키는 lysophosphatidic acid (LPA; 10 M)는 pSREmt-Luc과 pSREwt-Luc를 동일한 수준으로 자극하였다 (Fig. 2B). 즉 티타늄에 의한 c-fos SRE로의 신호 전달은 Elk-1p62TCF 경로와는 별개로 작용한다는 것을 의미한다(Fig.
2B). 대조실험으로 EGF (50 ng/mL)는 pSRE-mt를 pSRE- wt 보다 적은 정도로 자극시켰다. 이것은 EGF가c-fos SRE 를 활성화하기 위해서는 Elk-1p62TCF 경로가 더 중요하다는 것 을 의미한다. 상기의 결과들은 Titanium 입자의c-fos SRE로 의 신호 전달에 Ras-MAP kinase 경로가 관여 하지 않는다는 것을 의미한다.
이점을 다시, MEK 억제제가 티타늄에 의한c-fos SRE 활성 을 억제할 수 있는지 실험 하였다. 그 결과, PD098059 (1 M;
MEK 억제제)에 의한 억제 효과는 나타나지 않았으므로, 본 cascade는 MAP kinase 신호 전달계와 다른 경로로 전달된다 고 볼 수 있다(Table 1). 또한, MEK1-DN mutant을 이입한 MEK-1의 우성결손변이형은 티타늄 신호 전달에 대해 억제 효 과를 가지지 않았다(Fig. 2C). 반면, 대조 실험으로서 EGF에 의한 신호 전달계에서는 억제가 발생하였다. 이러한 결과에 의 해, 티타늄의c-fos로의 신호전달계에서, Ras-MAP kinase 경 로가 관여할 가능성은 배제되었다고 볼 수 있다.
3.
티타늄 신호계에서Rac GTPase
의 역할Rho family의 small GTPase가 관여할 가능성에 대해 분석 하기 위하여 pSRE-Luc (3 g)을 대조군인 pEXV (5 g)와 Rac1의 우성결손변이형으로 부호화한 pEXV-RacN17 (5 g)
* Values in percentage.
A reporter gene plasmid, pSRE-Luc (3 g), was transiently transfect- ed. Various inhibitors were pretreated each time before the addition of Titanium-particles (0.1 mg/mL). MAFP (5 M), MK886 (50 nM), Indomethacin (10 M), Aspirin (1 M), NS-398 (10 M), PD098059 (10 M), Wortmannin (50 nM). Two hours (3 h for the Titanium-particles) later, the luciferase activities were measured and normalized to the co- transfected -galactosidase activities. The results are the means of ± S.D. obtained from three samples.
Relative Luciferase Activity Inhibitors
Dosage - (%) Titanium (%)
- - 1.0±0.1 (100)* 2.3±0.1 (100)
MAFP 5 M 1.0±0.1 (100) 1.0±0.1 (43) MK886 50 nM 1.0±0.0 (100) 1.3±0.2 (56) INDOMETHACIN 10 M 1.0±0.2 (100) 1.0±0.2 (43) ASPIRIN 1 M 1.0±0.2 (100) 1.1±0.2 (48) NS-398 10 M 1.0±0.0 (100) 1.3±0.1 (56) PD098059 10 M 1.0±0.1 (100) 2.2±0.4 (96) WORTMANNIN 50 nM 1.0±0.3 (100) 1.2±0.1 (52) Table 1.Effect of Various Inhibitors in Titanium-induced SRE Activation
로 공동이입 하였다. 그 결과, 티타늄 입자가 유발하는 c-fos SRE-luciferase의 활성이 pEXV-RacN17에서는 59%로 감소 되었다(Fig. 3A). 즉 티타늄 입자가 유발하는c-fos SRE 활성 화 경로에는 Rho family의 small GTPase 단백질의 일종인 Rac이 필요하다는 것을 알 수 있다. 또한 Rac의 연관성을 Rat- 2와 Rat2-RacN17에서 SRE-luciferase 활성도를 비교하여 다 시 분석한 결과, 티타늄 입자는 Rat-2-luciferase 보다, Rat-2- RacN17-luciferase 활성을 더 적게 유발하였다(Fig. 3B). 대조 실험에서, LPA (1 M)는 SRE-luciferase 활성을 Rat-2와 Rat-2 RacN17에서 모두 동일한 정도로 자극하였다. 이러한 결 과들로 볼 때, 티타늄-c-fos SRE의 신호계에서 Rac의 역할이 필요함을 알 수 있다.
4.
티타늄 신호계에서cPLA
2, Lipoxygenase
와Cyclooxy- genase
의 역할일반적으로, Rac-c-fos SRE의 신호 전달계에서 cPLA2가 Rac의 중요한 하류 전달체로서 기여한다고 알려져 있다15,16). 그 러므로, 티타늄과 Rac의 관계를 고려할 때, 티타늄-c-fos SRE 신호계에서 cPLA2가 하류전달체로 관여할 가능성을 생각할 수 있다. MAFP (5 M; cPLA2 특이적 억제제)는 티타늄에 의 한 c-fos SRE활성을 완전히 억제하였다(Table 1). 동일하게 MK886 (50 nM; 5-Lo 억제제), Aspirin (1 M ; COX 억 제제), NS-398 (10 M; COX-II 선택적 억제제) 모두가 c- fos SRE 활성을 억제 하였다. 이러한 결과들은, cPLA2 그리 고 lipoxygenase (Lo)와 cyclooxygenase (COX)에 의한 arachidonic acid (AA) 대사가 티타늄-c-fos SRE의 신호 전 이에 관여한다는 것을 나타낸다.
5. Rac-
의존성 신호전이계: Titanium
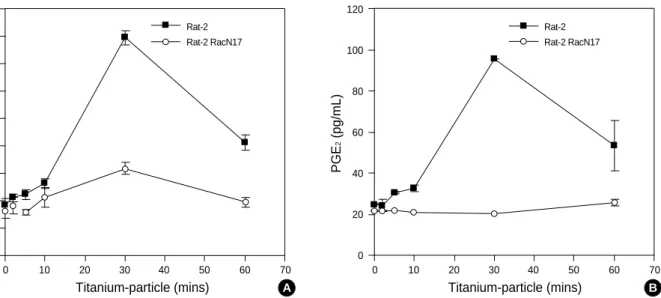
입자의eicosanoids
합성의 유도 경로티타늄 입자에 의해, 섬유 아세포에서도 eicosanoids의 합성이 유발되는지를 확인하기 위해, LTB4와 PGE2의 세포 내 변화를 측정하였다. 티타늄 입자(0.1 mg/mL)로 처리 후 30분 뒤에, LTB4와 PGE2의 양이 최대(기준치의 약 4배 이상)로 증가하였 다(Fig. 4). 그 다음 티타늄 입자에 의한 eicosanoids 합성 유발 경로가 Rac-cPLA2에 의존하여 작용하는지 분석하였다. 그 결 과 티타늄에 의한 PGE2의 증가는 Rat2-RacN17 clone에서는 일어나지 않았으며, 동일한 현상이 LTB4에서도 나타났다(Fig.
4). 이러한 결과들은 티타늄에 의한 eicosanoids의 합성 유발 기 전에는 Rac GTPase가 필요하다는 것을 의미한다. 유사하게 cPLA2의 특이적 억제제인 AACOCF3 (10 M)로 전처치 한 경우에도 LTB4와 PGE2의 합성이 현저히 감소 되었다(Fig. 5).
최근 Rac, cPLA2, phosphatidylinositol 3-kinase (PI3- kinase)의 활성이 tumor necrosis factor- (TNF- )에 의한 LTB4 합성의 경로에 관여한다는 것이 알려졌다16). 따라서, PI3-kinase가 티타늄에 의한 eicosanoids 합성에 연관되는지를 분석하였다. 그 결과 티타늄 입자에 의한 LTB4와 PGE2의 합성 은 PI3-kinase 억제제인 Wortmannin (50 nM)에 의해 현저 히 감소 되었다(Fig. 5). 이 결과는 PI3-kinase가 티타늄에 의 한 eicosanoids의 합성 기전에 연관됨을 나타낸다. 그러므로 이 상의 결과들은 PI3-kinase, Rac, 그리고 cPLA2가 티타늄에 의 한 eicosanoids의 합성 기전에 연관되어 되어 있으며, 이러한 eicosanoids의 합성은 티타늄에 의한c-fos SRE로의 신호전달 경로에 주요한 작용을 한다고 여겨진다.
Relative Luciferase Activity
7
6
5
4
3
2
1
0
Buffer Titanium LPA
Relative Luciferase Activity
7
6
5
4
3
2
1
0
Buffer Titanium LPA
Fig. 3.Role of Rac in the Titanium-induced signaling to c-fos SRE. A reporter gene plasmid, pSRE-Luc (3 g) was transiently co-trans- fected with 5 g of pEXV, pEXV-RacN17. (A) The effect of Rac was analyzed using Rat2-RacN17 cells. (B) Transfected cells were serum- deprived in 0.5% FBS/DMEM for 36 h before being treated with Titanium-particles (0.1 mg/mL), LPA (2 M). After the treatment, the luciferase activities were measured and normalized to the co-transfected -galactosidase activities. Values are representative of triple transfections.
Mock
pEXV Rat-2
Rat-2 RacN17 RacN17
B A
고 찰
본 실험에서 티타늄 입자가c-fos SRE의 활성을 증가시키는 것이 확인되었다. 이는 Rat-2 섬유 아세포에서 c-fos SRE는 티타늄 입자에 의한 핵 표적 서열 중의 하나라는 것을 의미한다.
또한, c-fos SRE로 전이되는 티타늄의 신호계에서 Rac, cPLA2, 5-Lo, 그리고 COX가 필수적인 하류 전달체로서 작용 한다는 것을 제시하였다. 또한 티타늄에 의한 본 ‘Rac-
cPLA2- eicosanoids- c-fos 신호전달계’는 LTB4와 PGE2 등 의 염증 매개체의 합성을 유발하는 신호기전이 될 수 있을 가능 성이 있다.
MAP kinase는 extracellular signal-regulated protein kinase (ERK)로도 알려져 있다. 이는 성장인자 등의 외부의 자극에 의한 분열촉진 신호전달의 중심역할을 수행하며, ser- ine/threonine kinase로서 세포의 생장과 분화에 중요한 유전자 발현을 조절한다1,21). 삼성분 복합체 인자(ternary complex
LTB4(pg/mL)
180 160 140 120 100 80 60 40 20
0
0 10 20 30 40 50 60 70
PGE2(pg/mL)
120
100
80
60
40
20
0
0 10 20 30 40 50 60 70
Fig. 4.Enhanced synthesis of LTB4and PGE2by Titanium in a Rac-dependent manner. (A) Rat-2 and Rat2-RacN17Asp17were exposed to 0.1 mg/mL Titanium-particles for the indicated time periods, and then the intracellular LTB4and PGE2were assayed using a specific ELISA. (B) Rat-2 and Rat2-RacN17Asp17cells were treated with Titanium-particles for the indicated time, after which the intracellular PGE2
was assayed as described in Materials and Methods section. Data are expressed as the means ±S.D. of three independent experi- ments. The statistical significance of LTB4and PGE2assays was assessed with an analysis of variance (p<0.01).
Rat-2 Rat-2 RacN17 Rat-2
Rat-2 RacN17
Titanium-particle (mins) Titanium-particle (mins)
LTB4(pg/mL)
120 100 80 60 40 20 0
PGE2(pg/mL)
300
250
200
150
100
50
0
Buffer Titanium
Fig. 5.Effects of Wortmannin and AACOCF3 on Titanium-particles-induced synthesis of LTB4and PGE2. (A) Rat-2 cells were exposed to 0.1 mg/mL Titanium-particle for 30 min in the presence or absence of Wortmannin (0.1 M) or AACOCF3 (10 M) and then intracellular LTB4was assayed using a specific ELISA. Inhibitors were added for 30 min prior to addition of the Titanium-particles. (B) Rat-2 cells were treated with 0.1 mg/mL Titanium-particles for 30 min, after which the intracellular PGE2was assayed as in the procudure A. Data are expressed as the mean ±S.D. of three independent experiments. The statistical significance of the LTB4assays and the PGE2
assays was assessed with the ANOVA test (p<0.05) and the unpaired t-tests (p<0.01), respectively.
Buffer Titanium Titanium+WortmanninTitanium+AACOCF3
Buffer Wortmannin AACOCF3
B A
B A
factors; TCFs)중 하나인 Elk-1p62TCF는 MAP kinase 경로 로 알려진 Ras-Raf-ERK cascade의 활성화에 반응하여c-fos SRE를 조절하는 것으로 알려져 있다2,4,27). Ras family G-단백 은 성장인자 등으로부터 들어오는 신호를, 세포분열의 주기를 조절하는 Ras/MAP kinase pathway로 전달하는 신호 전달 단백이다1). 최근 Rho family GTPases는, serum response factor (SRF)의 직접적인 활성화와 연관된 MAP kinase 비의 존성 경로를 통해c-fos SRE 활성화 신호 전달계에 역할을 하 는 것이 밝혀졌다12). 따라서, c-fos SRE는, Elk-1p62TCF 의존 성 경로 뿐만 아니라, 비의존성 신호 경로계를 통해 활성화 될 수 있다. 본 연구의 결과에 의해 티타늄의 c-fos로의 신호전달 계에서 Ras-MAP kinase 경로가 관여할 가능성은 배제되었다 고 볼 수 있다.
G-단백 superfamily 중에서 Rho family에 속하는 Rac GTPases는 Ras와 마찬가지로, GTPase activating protein (GAP)들에 의해 같은 방식으로 조절된다1). Rac은, 성장 인자 에 의한 actin filament의 재기질화, 즉 세포막 파상운동(mem- brane ruffling)을 조절한다10,22). 또한 세포 형질전환과 G세포 주기진행을 포함하는 넓은 범위의 생물학적 과정 조절에도 연관 되어져 왔다10). 최근에는 C2-ceramide, cytokines, environ- mental stresses 등에 의한 신호 전달에도 Rac이 관여 한다고 보고되고 있다2,12). 티타늄 입자는 세포에 대해서 이물질이므로, 일종의 agonist로 작용하여, Rac과 연관된 신호전달계를 자극할 수 있을 것이다. 예상대로, Rac1의 우성 결손 변이형으로 이입 한 pEXV-RacN17에서 티타늄 입자가 유발하는c-fos SRE 활 성화가 의미 있게 감소 하였다(Fig. 3). 이것은 c-fos SRE에 대한 티타늄의 신호전달계에서 Rac단백의 활성이 필수적이라는 것을 의미한다.
일반적으로, Rac의 활성화에 의해 AA가 유리된다는 것은 이 미 잘 알려져 있으며, 이러한 AA의 유리와 그 대사과정들이 바 로 Rac이 섬유 아세포의 actin 개작에 영향을 주는 분자생물학 적 기전으로 알려져 왔다15,16,19). 저자들도 Rac이c-fos SRE를 활성화 시키는 신호전달 경로에서‘cPLA2-AA’신호전달계가 매개하는 것을 보고한 바 있다29). 또한 최근에는‘Rac-cPLA2- AA’신호전달계가 hydrogen peroxide, ceramide, TNF- , Fas ligand, UV, 혹은 lipid second messenger 등에 의한 신 호 전달에 관여한다고 밝혀졌다9). 본 신호전달계가 파골 세포에 서도 작용할 경우 Rac과 c-fos의 활성화를 통해 actin 개작 (remodeling)과 세포막 파상운동을 조절하여 파골 세포의 분화 와 증식의 key regulator로서 작용할 가능성이 있다8). 그러나, 이점은 향후 파골 세포에서 직접 연구해야 할 내용이며, 세포 조작이 쉽지 않아 많은 어려움이 따를 것으로 생각 된다.
마모편에 의해 초래되는 삽입물 주위 골용해의 골-삽입물 경 계막(interfascial membrane)에서는 PGE2 농도가 증가되는 것 으로 알려져 있다5,6,7,23). 본 연구에 의한 티타늄의 신호전달계에 서는, Rac과 cPLA2에 아주 의존적인 양식으로 eicosanoids의
합성이 증가하였다(Fig. 4). 또한 PI3-kinase가 티타늄에 의한 eicosanoids의 합성 기전과c-fos SRE활성화에 연관됨을 고려 하면 PI3-kinase와 함께 Rac의 하류전달체로서 cPLA2, 5-Lo 와 COX에 의한 eicosanoids 합성 기전이 티타늄의c-fos SRE 로의 신호전달계에 주요한 작용을 한다고 여겨진다(Table 1)(Fig. 5).
마모편 입자들에 의한 골용해 과정에는, PGE2 이외의 다른 인자나 기전, 소위‘PG 비의존성 기전’으로서의 몇 가지의 신 호 전달계가 존재 할 것으로 생각할 수 있다3,5,7,8,17). TNF- , interleukin-1과 interleukin-6 같은 proinflammatory cytokine 들이 골용해 과정에 연관되어 있다11,13,23). 섬유 아세포에서c- fos SRE가 PI3-kinase, Rac, cPLA2의 순서를 거치며, LTB4 를 생성하면서 TNF- 의 표적 서열로 작용 할 수 있다9,16). 그 러므로 티타늄이 TNF- 등의 proinflammatory cytokine들과 함께 작용하면서‘Rac- cPLA2- eicosanoids-c-fos 신호전달 계’를 유도할 가능성이 입증되면 골 용해의 PG 비의존성 기전 은 물론, PG 의존성 경로의 일부가 밝혀질 수 있을 것이다. 또 한 본 신호전달계는 염증 반응을 유발하는 eicosanoids의 합성 기작의 하나일 가능성도 있다.
결 론
티타늄 입자의 자극에 의해 Rat-2 섬유 아세포의c-fos SRE 활성이 증가하며, 이 과정에서 Rac, cPLA2, 5-Lo, 그리고 COX의 작용이 필수적이다. 그러므로 티타늄에 의한 c-fos SRE의 활성화 경로에는‘Rac- cPLA2- eicosanoids- c-fos 신호전달계’가 존재한다.
참고문헌
1. Cho TJ, Choi IH, Chung CY, Sung HY, Sohn YJ and Kim DW:
Molecular characterization of congenital pseudoarthrosis of the tibia asso- ciated with neurofibromatosis. J. of Korean Orthop. Assoc. 35: 825-831, 2000.
2. Cosa OA, Chiariello M, Yu JC, et al:The small GTP-binding proteins Rac1 and CDC42 regulate the activity of the JNK/SAPK signaling path- way. Cell, 81: 1137-1146, 1995.
3. Eriksson AS and Thomsen P:Leukotrien B4, Interleukin-1 and leuko- cyte accumulation in titanium and PTEF chambers after implantation in the rat abdominal wall. Biomaterials, 12: 827-830, 1991.
4. Gille H, Sharrocks AD and Shaw PE:Phosphorylation of transcrip- tion factor p62TCF by MAP kinase stimulates ternary complex formation at c-fos promoter. Nature, 358: 414-417, 1992.
5. Glant TT, Jacobs JJ, Mikecz K, et al: Particulate-induced
prostaglandin and cytokine mediated bone resoption in an experimental system and in failed joint replacements. Am J Ther, 3: 27-41, 1996.
6. Goldring SR, Schiller A, Roelke M, O’’Neill D and Harris W: The synovial-like membrane at the bone-cement interface in loose total hip replacements and its proposed role in bone lysis. J Bone Joint Surg, 65(A):
575-584, 1983.
7. Goodman SB, Chin R and Magee F:Prostaglandin E2production by the membrane surrouding loose and fixed cemented tibial hemiarthroplas- ties in the rabbit knee. Clin Orthop, 284: 283-287, 1992.
8. Grigoriadis AE, Wang ZQ, Cecchini MG, et al:C-fos: a key regula- tor of osteoclast macrophage lineage determination and bone remodeling.
Science, 21; 266: 443-448, 1994.
9. Haliday EM, Ramesha CS and Ringold G:TNF induces c-fos via a novel pathway requiring conversion of arachidonic acid to a lipoxigenase metabolite. EMBO J, 10: 109-115, 1991.
10. Hall A: Small GTP-binding proteins and the regulation of the actin cytoskeleton. Annu Rev Cell Biol, 10: 31-54, 1994.
11. Haynes DR, Boyles SJ, Rogers SD, Howie DW and Vernon- Roberta B:Variation in cytokines induced by particles from different prosthetic materials. Clin Orthop, 352: 223-230, 1998.
12. Hill CS, Wynne J and Treisman R:The Rho family GTPase RhoA, Rac1 and CDC42Hs regulate transcriptional activation by SRF. Cell, 81:
1159-1170, 1995.
13. Jiranek W, Machado M, Jasty M, et al:Production of cytokines around loosened cemented acetabular components. Analysis with immunohistochemical techniques and in situ hybridization. J Bone Joint Surg, 75(A): 863-879, 1993.
14. Johansen FE and Prywes R:Two pathways for serum regulation of the c-fos serum response element require specific sequence elements and a minimal domain of serum response factor. Mol Cell Biol, 14: 5920-5928, 1994.
15. Kim BC and Kim JH:Nuclear signaling by Rac GTPase: Essential role of phospholipase A2. Biochem J. 326: 333-337, 1997.
16. Kim BC, Lee MN, Kim JY, et al:Role of phosphatidylinositol 3-Kinase and Rac in the nuclear signaling by tumor necrosis factor- in Rat-2 fibroblasts. J Biol Chem, 274: 24372-24377, 1999.
17. Lerner UH and Ohlin A: Tumor Necrosis Factors alpha and beta can stimulate bone resorption in cultured mouse calvariae by a prostaglandin independent mechanism. J Bone Miner Res, 8: 147-155, 1993.
18. Miyauchi A, Kuroki Y, Fukase M, Fujita T, Chihara K and
Shiozawa S:Persistent expression of proto-oncogene c-fos stimulates osteoclast differentiation. Biochem Biophys Res Commun, 30: 1547-1555, 1994.
19. Peppelenbosch MP, Tertoolen LGJ, Hage WJ and De Laat SW:
Epidermal Growth Factor-induced Actin remodeling is regulated by 5- Lipoxygenase and Cyclooxygenase products. Cell, 74: 565-575, 1993.
20. Piechaczyk M and Blanchard JM: C-fos proto-oncogene regulation and function. Critical Reviews in Oncology/Hematology, 17: 93-131, 1994.
21. Reobuck KA, Jacobs JJ and Glant TT: New horizons in orthopaedic research: elucidation of cellular signal transduction pathways. J Bone Joint Surg, 81(A): 599-602, 1999.
22. Ridley AJ, Peterson HF, Johnston CL, Diekmann D and Hall A:
The small GTP-binding protein Rac regulates growth factor-induced membrane ruffling. Cell, 70: 401-410, 1992.
23. Shanbhag AS, Jacobs JJ, Black J, Galante JO and Glant TT:Cellu- lar mediators secreted by interfacial membranes obtained at revision total hip arthroplasty. J Arthroplasty, 10(4): 498-506, 1995.
24. Shanbhag A, Jacobs JJ, Black J, Galante JO and Glant TT:Effect of particles on fibroblast proliferation and bone resorption in vitro. Clin Orthop, 342: 205-217, 1997.
25. Treisman R:The SRE: A growth factor responsive transcriptional regu- lator. Seminar in Cancer Biology, 1: 47-58, 1990.
26. Vermes C, Roebuck KA, Chandrasekaran R, Dobai JG, Jacobs JJ and Glant TT: Particulate wear debris activates protein kinases and nuclear factor B, which down-regulates type I collagen synthesis in human osteoblasts. J Bone Miner Res, 15: 1756-1764, 2000.
27. Whitmarsh AJ, Shore P, Sharrocks AD and Davis RJ:Integration of MAP kinase signal transduction pathways at the Serum Responsive Element. Science, 269: 403-407, 1995.
28. Woo CH, Eom YW, Yoo MH, et al:TNF- generates Reactive Oxy- gen Species generation via cPLA2-linked cascade. J Biol Chem, 275(41):
32357-32362, 2000.
29. Woo CH, Kim BC, Kim KW, et al:Role of cytosolic phospholipase A2
as a downstream mediator of Rac in the signaling pathway to JNK stimu- lation. Biochem Biophys Res Commun, 268(1): 231-236, 2000.
30. Yao J, Glant TT, Lark MW, et al:The potential role of fibroblasts in periprosthetic osteolysis: Fibroblast response to titanium particles. J Bone Miner Res, 10: 1417-1427, 1995.
Purpose :In order to understand the intracellular signaling pathway involving the c-fos gene expression that is caused by Titanium-particles, we analyzed the involvement of Rac, cytosolic phospholipase A2, and eicosanoids (e.g. leukotriene B4and prostaglandin E2) as well as c-fos.
Materials and Methods :We tested whether or not Titanium-particles activate a c-fos serum response element in Rat-2 fibroblasts. To measure the activity of the c-fos serum response ele- ment, we analyzed the serum response element using a luciferase reporter system. The luciferase activity was measured using a scintillation spectrophotometer. Next, we analyzed the involvement of Rac and the eicosanoid synthesis mechanisms which are downstream mediators of Rac in the c-fos serum response element activation cascade.
Results : Titanium-particles cause an activation of the c-fos serum response element and this activation was selectively repressed by RacN17 and by pretreatment of the inhibitors of cytosolic phospholipase A2, cyclooxygenase or 5-lipoxygenase. Eicosanoid synthesis was increased in a Rac-dependent manner in response to the presence of Titanium- particles.
Conclusion :‘Rac, a member of G-protein, which is involved in the eicosanoid synthesis’may play a critical role in the Titanium-induced signaling cascade. Thus, we speculated that the ‘Rac- cytosolic phospholipase A2-eicosanoids-c-fos cascade’may be a possible mechanism that pro- duces eicosanoid synthesis caused by Titanium-particles in the periprosthetic osteolytic process.
Key Words : Fibroblast, Titanium, Rac, Eicosanoid, c-fos
The Signaling Pathway of G-protein Rac and Eicosanoid Synthesis by Titanium Particles
Sang-Soo Lee, M.D., Jae-Young Her, M.D., Chang-Hoon Woo, M.S.*, Jae-Hong Kim, Ph.D.*, and Jun-Dong Chang, M.D.**
Department of Orthopaedic Surgery, Pohang St. Mary’s Hospital, Pohang; Graduate School of Biotechnology, Korea University, Seoul*; Department of Orthopaedic Surgery, College of Medicine, Hallym University, Seoul, Korea**
Abstract
Address reprint requests to Jun-Dong Chang, M.D.
Department of Orthopaedic Surgery, Hangang Sacred-Heart Hospital 94-200 Youngdeungpo-dong, Youngdeungpo-gu, Seoul 150-020, Korea Tel: +82-2-2639-5301 Fax: +82-2-2631-9337
E-mail: [email protected]
1. 제 3차 아시아 태평양 인공관절 학술대회
제 3차 아시아 태평양 인공관절 학술대회(The 3rd Asia-Pacific Arthroplasty Society 2001, Seoul, Korea 일명 APAS 2001, Seoul)가 2001년 9월 13(목)-16일(일)서울 조선호텔에서 개최됩니다. 대회 내용은 인공 고관절, 슬관 절 분야의 심포지움, 세계적인 관절염 분야 권위자의 초청특강, 논문발표, 포스터 전시, 의학기기 전시로 구성되며,이 번 학술대회에 국내 정형외과 의사들의 많은 참여를 바랍니다. 홈페이지로 등록 및 초록접수가 가능합니다.
�일 시: 2001년 9월 13(목)-16일(일) 4일간
�장 소: 서울 웨스턴 조선호텔(1층 회의장, 2층 전시장)
�주 제: “Toward the perfect artificial joints”
�회의구성: -인공 고관절,인공 슬관절 분야의 심포지움 -초청연사의 특강
-논문발표 -포스터 전시 -의학기기 전시
�예상 참가국: 아시아 태평양 16개국외(한국, 일본, 호주, 뉴질랜드, 중국, 대만, 홍콩, 인도, 태국, 베트남, 말레이시 아, 파키스탄, 필리핀, 싱가포르, 인도네시아 외 …)
�공식언어: 영어
�주 최: 아시아 태평양 인공관절 학회 서울 사무국(조직위원장: 경희대학교 유명철 교수)
�후 원: 대한고관절학회, 대한슬관절학회
�회의 사무국: (주)한진관광 APAS 2001 Seoul 사무국, 담당: 김성민 서울특별시 중구 소공동 51 해운센터 신 관 5층 100-770
Tel : 02-726-5557, Fax : 02-778-2514,
E-mail : [email protected]