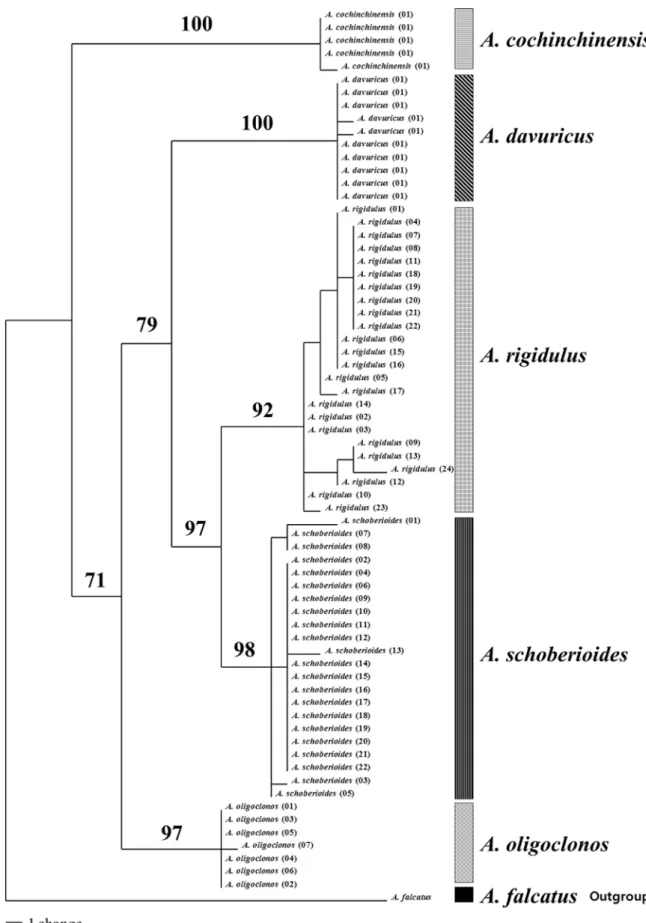
Systematic study of Korean Asparagus L. based on morphology and nuclear ITS sequences
Seong-Hyun Cho and Young-Dong Kim*
Department of Life Science, Hallym University, Chuncheon 200-702, Korea (Received 24 August 2012; Revised 6 September 2012; Accepted 14 September 2012)
외부형태와 ITS 염기서열에 기초한 한국산 비짜루속 식물의 분류학적 고찰
조성현·김영동*