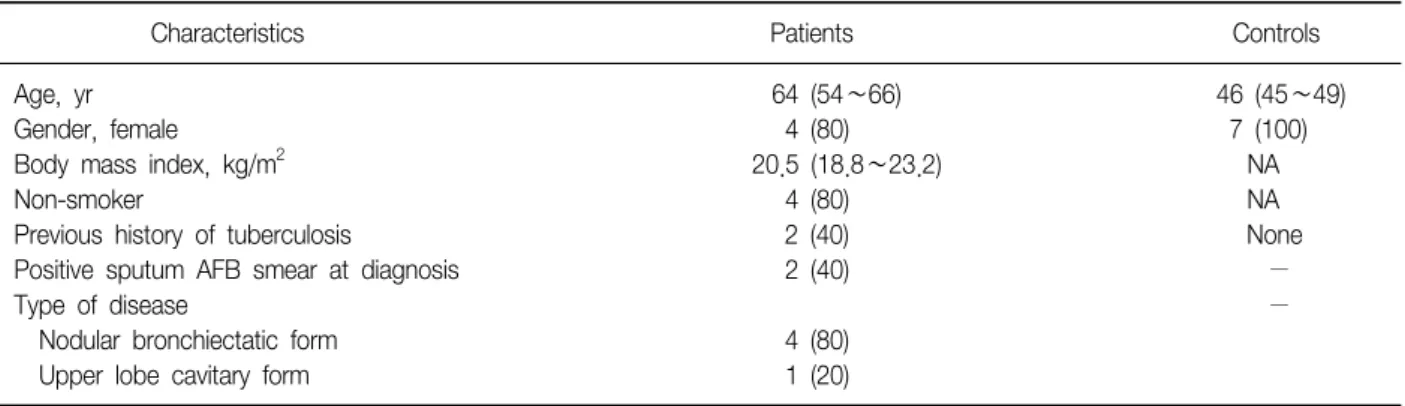
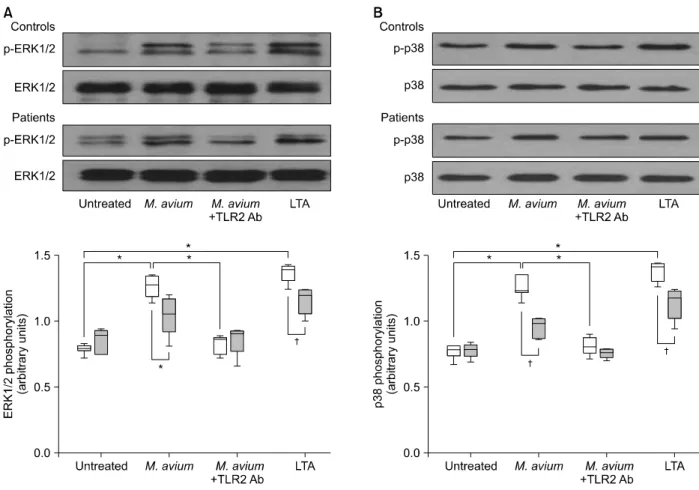
Tuberc Respir Dis 2012;72:275-283
CopyrightⒸ2012. The Korean Academy of Tuberculosis and Respiratory Diseases. All rights reserved.
Impaired Expression of MAPK Is Associated with the Downregulation of TNF-α, IL-6, and IL-10 in Mycobacterium abscessus Lung Disease
Yun Su Sim, M.D.
1*, Su-Young Kim, Ph.D.
1*, Eun Joo Kim, Ph.D.
1, Sung Jae Shin, Ph.D.
2, Won-Jung Koh, M.D.
11