Korean J Physiol Pharmacol 2018;22(4):427-436 https://doi.org/10.4196/kjpp.2018.22.4.427
Author contributions: Conception and design of study: S.H.K. and J.C.K.
Acquisition of data, analysis and/or interpretation of data: Y.S.K., E.B.N., B.W.S., S.H.K., and G.D.P. Drafting the manuscript: S.H.K. Revising the manu- script, revising the manuscript critically for important intellectual content:
D.Y.S., G.D.P., and S.H.K.
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License, which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Copyright © Korean J Physiol Pharmacol, pISSN 1226-4512, eISSN 2093-3827
INTRODUCTION
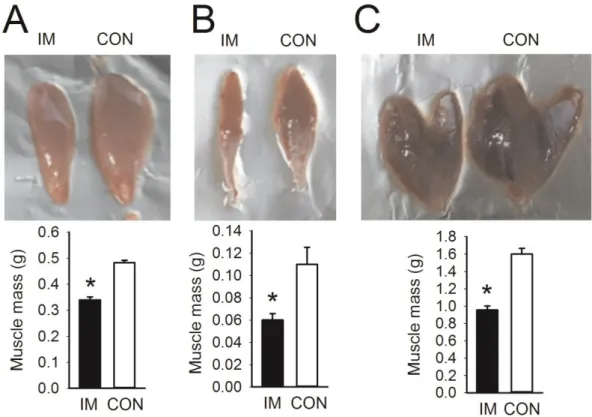
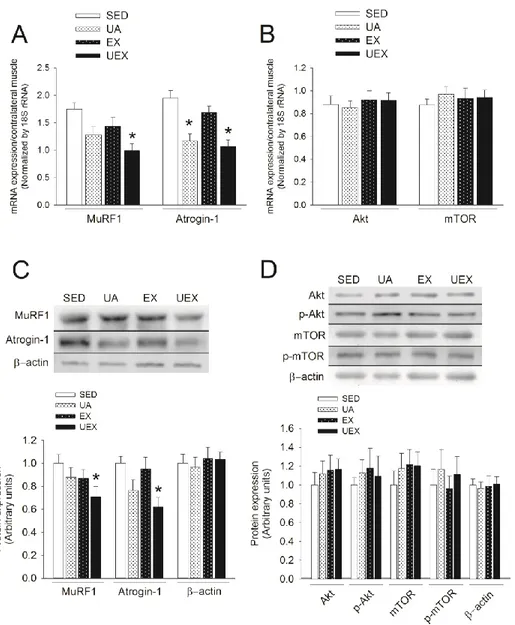
Muscle atrophy, caused by a decrease in physical activity or aging, leads to problems related to an increase in body fat and de- creased muscle strength and cardiovascular endurance, resulting in fatigue [1-4] and deterioration of bone health [5,6], ultimately affecting the activities of daily living and quality of life [7-9].
Thus, efforts to improve muscle atrophy in the increasing elderly population should be explored.
Hypertrophy of the skeletal muscles is required to prevent muscle atrophy [10], and exercising with weights, including resis- tance exercise training, is effective [11]. According to recent stud- ies, even though muscle size and strength can be increased with low-intensity resistance training [12], higher-intensity resistance training (i.e., 80% of one repetition maximum) is more effective [13-15]. However, many adults, including elderly people (more than 70%), are inactive or sedentary, mainly participating only in light physical activities such as stretching or walking [16-18],
Original Article
Concurrent treatment with ursolic acid and low-intensity treadmill exercise improves muscle atrophy and related outcomes in rats
Jae Cheol Kim 1 , Yun Seok Kang 1 , Eun Bi Noh 1 , Baek Woon Seo 1 , Dae Yun Seo 2 , Gi Duck Park 3, *, and Sang Hyun Kim 1, *
1