Introduction
Ischemic stroke is caused by a reduction of blood flow and re- sults in the deficiency of glucose and oxygen supply to the brain.
Increased levels of reactive oxygen species (ROS) are the major reason of tissue injury after cerebral ischemia. ROS are consid- ered regulatory molecules and known to act as second messen- gers during ischemia-reperfusion (I/R). ROS are generated dur- ing ischemia-reperfusion injury and promote the apoptosis pathway.
1Antioxidant attenuates oxidative damage induced by I/R through decreasing mechanism of ROS production.
2Specifi- cally, dehydroascorbic acid (DHA) and glutathione (GSH) play an important physiological role in cells as a antioxidant, free rad- ical scavenger.
3,4A high intracellular GSH concentration protects cells against ROS by reacting directly with free radicals
5and GSH is involved in a variety of mechanism related with human diseases.
6-10DHA inhibited cell death by increasing the GSH lev-
els.
11Melatonin is synthesized in the pineal gland and has been known to exert direct and indirect antioxidant actions.
12,13Mela- tonin reduces the harmful cellular actions of ROS in I/R barin injury.
14,15In in vivo cerebral ischemia model, previous research- es demonstrates that melatonin treatment attenuates brain dam- age against ischemia-hypoxia induced injury.
16,17Previous stud- ies confirmed that melatonin treatment led to the maintenance of higher contents of GSH through its regulation of the AA-GSH cycle. Melatonin strongly is involved in AA-GSH cycle.
18In the present study, we examined the synergetic beneficial effect of melatonin on the combinational treatment of DHA and GSH.
The blood brain barrier (BBB) includes biochemical mecha- nisms that control the exchange of materials between blood and the brain. BBB is crucial in central nervous system considering regulation of the brain microenvironment for neuron’s optimal functioning.
19Tight junctions are important BBB structural components that seal the gaps between adjacent endothelial cells
Vascular Neurology 2013;5:35-41 ISSN 2092-6855
The Combinational Treatment of Antioxidants Protects
Brain Endothelial Cells Against Oxygen-Glucose Deprivation Followed by Reperfusion Induced Injury
So Mang Kang,
1,2Juhyun Song,
1Won Taek Lee,
1Kyung Ah Park,
1Kyoung Min Lee,
3Jong Eun Lee
1,21Department of Anatomy, 2BK21 Plus Project for Medical Sciences, Yonsei University College of Medicine, Seoul, Korea
3Seoul National University College of Medicine, Seoul, Korea
Received November 15, 2013 Revised November 21, 2013 Accepted November 27, 2013 Correspondence Jong Eun Lee, PhD Department of Anatomy, Brain Korea 21 Project for Medical Science, Yonsei University College of Medicine, 50-1 Yonsei-ro, Seodaemun-gu, Seoul 120-752, Korea
Tel +82-2-2228-1646, 1659 Fax +82-2-365-0700 E-mail [email protected]
Antioxidants including dehydroascorbic acid (DHA), glutathione (GSH) and melatonin have cel- lular protective effects on murine brain endothelial (bEnd.3) cells after oxidative stress and isch- emic injury. Up to date, there are no studies that the combinational treatment of these antioxi- dants boosts the cellular protective effect on bEnd.3 cells. In this study, we investigated the protective effect of antioxidant combinational treatment on bEnd.3 cells after oxygen-glucose de- privation followed by reperfusion (OGD/R) induced injury. The effect of antioxidants combina- tional treatment on the bEnd.3 cells was examined by cell viability test, western blot analysis, im- munocytochemistry. The combinational treatment of DHA (1 mM), GSH (1 mM) protects bEnd.3 cells after OGD/R induced injury and also melatonin (10 μM) treatment boosts the pro- tective effect in combinational treatment of DHA and GSH on bEnd.3 cells after OGD/R induced injury. Western blot analysis suggests that the combinational treatment of DHA, GSH and melato- nin attenuated the expression of vascular endothelial growth factor (VEGF), phosphorylation of ERK compared with each single treatment after OGD/R induced injury. Claudin 5, hypoxia in- ducible factor-1α (HIF-1α) and VEGF immunostaining data suggests the preventive effect of anti- oxidants combinational treatment. In conclusion, the combinational treatment of antioxidants el- evates the cellular protective effect on bEnd.3cells against OGD/R induced injury.
Vascular Neurology 2013;5:35-41 Key Wordsaa Dehydroascorbic acid, Glutathione, Melatonin,
Oxygen-glucose deprivation followed by reperfusion, Murine brain endothelial cells.
and regulate cellular permeability.
20,21We investigated whether the antioxidants combinational treatment preserves the tight junction protein. Hypoxia inducible factor-1α (HIF-1α) is a key regulator of hypoxia-induced gene expression and induces a va- riety of mechanisms including angiogenesis, cell survival, cell death after hypoxia.
22In addition, vascular endothelial growth factor (VEGF) has a detrimental effect in endothelium permea- bility leading to vasogenic edema.
23Therefore, activation of ERK contributes to cell death
24,25and regulates the expression of VEGF.
26We examined whether antioxidant combinational treat- ment contributes to the expression of VEGF and phosphoryla- tion of ERK1/2. In the present study, we investigated the syner- getic protective effect of antioxidants combinational treatment on bEND3 cells against OGD injury.
Materials and Methods
Cell culture
Murine brain endothelial cells (bEnd.3 cells, Manassas, VA, USA) were purchased from ATCC and cultured in Dulbecco’s modified Eagle’s medium (DMEM; Hyclone Laboratories, Lo- gan, UT, USA), supplemented with 10% (v/v) fetal bovine serum (FBS; Hyclone Laboratories, Logan, UT, USA) and 100 units/mL of penicillin/streptomycin (Hyclone Laboratories, Logan, UT, USA), at 37°C in a humidified atmosphere in the presence of 5%
CO
2. Culture medium was changed every 2 days.
27Drug treatment
DHA was dissolved in 0.1 M bicarbonate buffer and GSH and Melatonin were melted in phosphate-buffered saline (PBS). Cul- tured bEnd.3 cells were divided into four groups as follows: 1) Control group, incubated with normal media; 2) Experimental group, cultured in non-treated medium for 6 hours after 6 hours of OGD injury; 3) combinational treatment of DHA and GSH group, corresponding to cells incubated in the presence of DHA and GSH for 6 hours (1 mM DHA+1 mM GSH) after OGD in- jury; and 4) combinational treatment of DHA, GSH and MEL group, treated DHA, GSH and MEL for reperfusion 6 hours (1 mM DHA+1 mM GSH + 10 μM Melatonin).
28Oxygen-glucose deprivation and reperfusion Confluent cells were transferred to an anaerobic chamber (Forma Scientific, Marietta, OH, USA) (O
2tension, 0.1%), washed three times with PBS. Then culture medium was re- placed with deoxygenated glucose-free balanced salt solution and cells were incubated for 6 hours. Following OGD injury, cells were incubated for 6 hours under normal growth condi- tions with or without drug treatment.
29Cell viability assays
Cells (5×10
4cells/well) were seeded into 48-well plates, and went through all experiment procedures, including pretreat-
ment, OGD injury and reperfusion. After that, the cells were rinsed twice with PBS, then 3-[4,5-dimethylthiazol-2-yl]-2,5-di- phenyl tetrasodium bromide (MTT, 2 mg/mL, Sigma, St. Louis, MO, USA) was added to each well. After 6 hours of incubation, medium were removed and dimethyl sulfoxide (DMSO) was added for solubilizing purple formazan product formed by MTT treatment. Then, supernatant from each well (200 μL/well) was replaced into 96-well plates for reading the absorbance with an ELISA reader at 570 nm. All experiments were repeated at least three times with similar results. The mean absorbance for the triplicate cultures of each drug was calculated, and the mean blank value was subtracted from these. Cell viability in the con- trol medium without any treatment was represented as 100%.
Cell viability displayed the relative value compared with the con- trol group.
27Hoechst 33258 and propidium iodide staining Cell viability was evaluated by staining bEnd.3 cells with Hoechst 33258 dye (Sigma, St. Louis, MO, USA) and propidium iodide (PI; Sigma, St. Louis, MO, USA). Hoechst dye was added to the culture medium (2-3 μg/mL) and then the samples were kept at 37.8°C for 30 min. PI solution was then added (2-5 μg/mL) just before observing with an fluorescent microscope (BX51;
Olympus) equipped with epifluorescence and a UV filter block.
Hoechst-positive, PI-positive and Hoechst-positive, PI-negative cells were counted as dead and live cells, respectively.
30Western blot analysis
After pretreatment, OGD injury and restoration, the cells were rapidly washed with ice-cold PBS, scraped and collected.
Cell pellets were lysed with ice-cold RIPA buffer (Sigma, St.
Louis, MO, USA). The lysates were centrifuged at 13200 rpm for 1 h at 4°C to yield whole-cell extracts. The protein content of the cell lysates was quantified using BCA method (Pierce, Rockford, IL, USA). Protein (20 μg) was separated on a 10% SDS-poly- acrylamide (PAGE) gel and transferred onto a polyvinylidene difluoride (PVDF) membrane. After blocking with 5% bovine serum albumin prepared in Tris-buffered saline/Tween [TBS-T;
20 nM Tris (pH 7.2); 150 mM NaCl; 0.1% Tween 20] for 1 h at room temperature (RT), immunoblottings (overnight at 4°C) were reacted with primary antibodies that specifically detect VEGF (1 : 1000), ERK (1 : 1000), p-ERK (1 : 2000), Beta actin (1 : 2000) (all purchased from Millipore, Billerica, MA, USA). Then, the immunoblots were reacted with HRP-linked anti-mouse and -rabbit IgG antibodies purchased from Abcam (Cambridge, MA, USA) for 1 h at RT, and enhanced chemiluminescence was per- formed by ECL (Pierce, Rockford, IL, USA).
27Immunocytochemistry (ICC)
The expression of VEGF, claudin-5 and HIF-1α in bEnd.3
cells was confirmed by immunocytochemistry (ICC). The ex-
pression of VEGF, claudin-5 and HIF-1α in bEnd.3 cells was
confirmed by immunocytochemistry. All the experimental groups was washed 3 times with phosphate-buffered saline, fixed with 4% paraformaldehyde for 3 hours, and then washed with PBS. bEnd.3 cells were permeabilized with 0.025% Triton X-100 and were blocked for 1 hour at room temperature with dilution buffer (Invitrogen life science, Carlsbad, CA, USA). Primary an- tibody rabbit anti VEGF (1 : 500, Millipore, Billerica, MA, USA), rabbit anti HIF-1 α (1 : 500, Santa Cruz Biotechnology, Santa Cruz, CA, USA) prepared in the dilution buffer was added to the samples and incubated for 3 hours at room temperature. Pri- mary antibody was removed and cells were washed 3 times for 3 minutes each with PBS. Later samples were incubated with FITC-conjugated goat anti rabbit second antibodies (1 : 200, Jackson Immunoresearch, West Grove, PA, USA) or Rhoda- mine-conjugated donkey anti rabbit second antibodies (1 : 500, Millipore, Billerica, MA, USA) for 2 hours at room temperature.
Cells were washed again 3 times for 3 minutes each with PBS and stained with 4’,6-diamino-2-phenylindole (1 : 100, Invitro- gen life science, Carlsbad, CA, USA) for 10 minutes at room temperature. The fixed samples were imaged using Zeiss LSM 700 confocal microscope (Carl Zeiss Microimaging, Thorn- wood, NY, USA).
Statistical analysis
Statistical comparisons were performed by using an indepen- dent t-test for 2 groups or analysis of variance with the SPSS software. Values are expressed as mean±S.E.M of 3-6 experi- ments. Differences were considered significant at p<0.1, p<0.05,
p<0.01.Results
Combination treatment of antioxidants enhances cell viability of bEND3 cells against OGD/R Induced Injury
We examined MTT [3-(4,5-dimethylthiazol-2-yl)-2,5-diphe- nyl tetrasodium bromide] assay commonly used as cell viability test for confirming the difference of cell viability between groups after OGD injury. Fig. 1A shows that DHA group and GSH group and MEL group elevate the cell viability than it of EC group. All single treatment groups have a cellular protective ef- fect in bEND3 cells against OGD/R injury. All combinational treatment groups (D+G, G+M, M+D, D+G+M groups) increase the cell viability of bEND3 cells compared to single treatment groups (DHA, GSH, MEL group). Especially, D+G+M group significantly elevates the cell viability of bEND3 cells compared with D+G group. Melatonin boosts the cell viability of bEND3 cells in the combination treatment group of DHA and GSH after OGD injury (Fig. 1A). Fig. 1B indicates that EC group shows that PI positive cells (red color) as a marker of death cells in- creases markedly in comparison with NC group. In comparison EC group, D+G group and D+G+M group decreases consider-
ably PI positive cells (Fig. 1B). Fig. 1B suggests that the combina- tional treatment contributes to reduction of death cells on bEND3 cells after OGD injury (p<0.1, p<0.05, p<0.001).
60 50 40 30 20 10
0 EC DHA GSH MEL D+G G+M M+D D+G+M
Cell viability (%)
†
* *
†
A
Figure 1. The cell viability measurement of bEND3 cells after OGD/R induced injury. A: The cell viability is lower in EC group due to OGD/R injury compared to NC group (100%). Also, the cell viability is higher combination treatment groups than single treatment groups. Especially, D+G group and D+G+M group sig- nificantly increases the cell viability of bEND3 cells after OGD/R injury. B: Hoechst and PI immunostaining shows that PI positive cells (red color) as a maker of death cells decreases by treating combinational antioxidants compared to EC group. PI positive cells increases in EC group compared with NC group due to OGD/R injury. PI positive cells decreases in D+G group and D+G+M group compared with EC group. Hoeschst: blue, PI: red.
*p<0.1, †p<0.05, ‡p<0.001. bEND3: brain endothelial, OGD/R: ox- ygen-glucose deprivation followed by reperfusion, NC: normal control group, EC: experimental group, DHA: DHA treatment group, GSH: GSH treatment group, MEL: melatonin treatment group, D+G: DHA and GSH treatment group, G+M: GSH and Melatonin treatment group, M+D: Melatonin and DHA treatment group, D+G+M: DHA and GSH and Melatonin treatment group.
B
Hoeschst
NC
EC
D+G
D+G+M
PI Merge
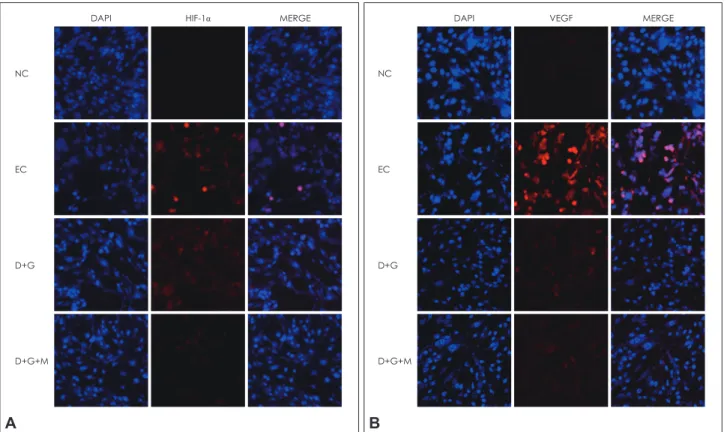
Combination treatment of antioxidants reduces the expression of HIF-1
αand VEGF on bEND3 cells against OGD/R induced injury
To measure the expression of HIF-1α and VEGF on bEND3 cells against OGD injury, we conducted ICC using HIF-1α and VEGF antibodies. The expression of HIF-1α as a inducer of hy- poxia related genes increases conspicuously in EC group and D+G group reduces HIF-1α positive cells (red color) compared to EC group (Fig. 2A). Specifically, D+G+M group reduces HIF- 1α positive cells among all groups (Fig. 2A). The expression of HIF-1α is attenuated by antioxidant combination treatment un- der hypoxia condition. Fig. 2B shows that the expression of VEGF (red color) increases evidently in EC group and decreases in combination treatment groups (D+G group and D+G+M group) (Fig. 2B). The expression of HIF-1α and VEGF increases markedly in EC group and decreases in combinational treat- ment groups including D+G group and D+G+M group. The ex- pression pattern of HIF-1α and VEGF is similar in all groups.
Combination treatment of antioxidants preserves tight junction protein on bEND3 cells against OGD/R induced injury
To measure the expression of claudin 5 known as tight junc- tion protein on bEND3 cells after OGD injury, we conducted using ICC claudin 5 antibody. Fig. 3 indicates that the expression of claudin 5 (green color) decreases substantially in EC group compared to NC group. Claudin 5 as tight junction protein re- duces against OGD injury. However, in combinational treatment groups, claudin 5 protein is preserved in comparison with EC group. The combinational treatment of DHA, GSH and Melato- nin has a preventive effect of tight junction protein on bEND3 cells against OGD injury.
Combination treatment of antioxidants reduces vascular permeability of bEND3 cells through VEGF and ERK1/2 pathway against OGD/R induced injury
To determine the protein expression of VEGF and phosphor- ERK1/2 on bEND3 cells after OGD injury, we conducted west- ern blot analysis using VEGF and phosphor-ERK1/2 antibodies.
Fig. 4A shows that the expression of VEGF considerably increas-
Figure 2. Immunocytochemistry (ICC) to investigate the expression of HIF-1α and VEGF on bEND3 cells after OGD/R induced injury. A:
HIF-1α positive cells increases in EC group due to OGD/R injury compared to NC group. In addition, HIF-1α positive cells decreases in D+G group and D+G+M group compared to EC group. HIF-1α decreases more in D+G+M group than D+G group. HIF-1α: red, DAPI: blue.
B: VEGF positive cells increases in EC group due to OGD/R injury compared to NC group. VEGF positive cells decreases in combination group such as D+G group and D+G+M group compare with EC group. Especially, VEGF known as vascular permeability maker decreases in D+G+M group compared to D+G group. VEGF: red, DAPI: blue.HIF-1α: Hypoxia inducible factor-1α (HIF-1α), VEGF: vascular endothe- lial growth factor, bEND3: brain endothelial, OGD/R: oxygen-glucose deprivation followed by reperfusion, EC: experimental group, NC: nor- mal control group, DAPI: 4’,6-diamidino-2-phenylindole.
A
DAPI
NC
EC
D+G
D+G+M
HIF-1α MERGE
B
DAPI
NC
EC
D+G
D+G+M
VEGF MERGE
es in EC group compared to NC group. Also D+G+M group shows significantly decreases compared with MEL group and D+G group (Fig. 4A). Fig. 4A suggests that melatonin treatment boosts the decrease of VEGF expression in combinational treat- ment of DHA and GSH. Fig. 4B shows that the phosphorylation of ERK1/2 increases in EC group compared with NC group and decreases considerably in D+G group. Especially, the phosphor- ylation of ERK1/2 decreases conspicuously in D+G+M group in comparison with all groups. Fig. 4B indicates that melatonin is associated with the decreased phosphorylation of ERK in com- binational treatment of DHA and GSH group.
Discussion
Reactive oxygen species (ROS) are important neurotoxic fac- tors in cerebral ischemia-reperfusion injury. ROS increase the susceptibility of brain tissues to ischemic damage and induce a variety of molecular cascades and finally results in BBB hyper- permeability, brain edema, hemorrhage, inflammation and cell death.
31BBB disruption are commonly observed in the patients
with various neurological disorders including stroke and de- mentia.
32,33Previous researches suggested that antioxidants pre- serve BBB disruption and decreases ROS after cerebral ischemia reperfusion after stroke in vivo
34-36and in vitro.
37,38DHA as an ox- idized form of ascorbic acid (AA) prevents cellular apoptosis
39and also ameliorates endothelial dysfunction related to nitric ox- ide (NO) activity.
40In in vivo stroke model, DHA significantly reduce the infarct volume and neurologic deficits.
11DHA is taken up through the glucose transport mechanism
41and is regenerat- ed to AA at the expense of the GSH within the cells.
42DHA in-
Figure 3. Immunocytochemistry (ICC) for confirming expression of claudin 5 on bEND3 cells in OGD/R induced injury. Claudin 5 positive cells (green) decreases in EC group compared to NC group due to OGD/R injury. Also, claudin 5 increases in combina- tional groups such as D+G group and D+G+M group compare with EC group. Specifically, claudin 5 known as tight junction pro- tein preserves in D+G+M group compared to D+G group. Claudin 5: green, DAPI: blue. bEND3: brain endothelial, OGD/R: oxygen- glucose deprivation followed by reperfusion, EC: experimental group, NC: normal control group, D+G: DHA and GSH treatment group, D+G+M: DHA and GSH and Melatonin treatment group, DAPI: 4’,6-diamidino-2-phenylindole.
DAPI
NC
EC
D+G
D+G+M
Claudin 5 MERGE
Figure 4. Western blot analysis for confirming vascular permeabili- ty of bEND3 cells in OGD/R induced injury. A: The protein expres- sion of VEGF, vascular permeability maker, increases in EC group owing to OGD/R injury compared to NC group. In antioxidant com- binational treatment group, the protein expression of VEGF de- creases compared with EC group. In detail, the expression of VEGF decreases more in D+G+M group compared to D+G group.
(B) The protein expression of ERK and phosphor-ERK increases in EC group. The protein expression of phosphor ERK decreases in D+G group and D+G+M group. *p<0.001, †p<0.05, ‡p<0.1, bEND3:
brain endothelial, OGD/R: oxygen-glucose deprivation followed by reperfusion, VEGF: vascular endothelial growth factor, p-ERK:
phospho-extracelluar single-regulated kinases, NC: normal control group, EC: experimental group, MEL: only melatonin treated group, ERK: extracelluar single-regulated kinases, D+G: DHA and GSH treatment group, D+G+M: DHA and GSH and Melatonin treatment group.
0.7 0.6 0.5 0.4 0.3 0.2 0.1
0 NC EC MEL D+G D+G+M
Relative optical density
†
A
*
VEGF B actin
1 0.9 0.8 0.7 0.6 0.5 0.4 0.3 0.2 0.1
0 NC EC MEL D+G D+G+M
Relative optical density
†
†
‡
B
p-ERK ERK B actin
hibited H
2O
2-induced cell death by increasing the GSH levels.
11GSH as a antioxidant plays also an important free radical scav- enger in cells.
3,4GSH is involved in a variety of mechanism in- cluding cell survival pathway and protects cells against ROS.
5-10Melatonin is synthesized in the pineal gland and has been known to involved in antioxidant actions.
12,13Therefore, melato- nin reduces the cellular damage by ROS in I/R barin injury.
14,15In in vivo cerebral ischemia model, previous researches demon- strates that melatonin treatment attenuates brain damage against ischemia-hypoxia induced injury.
16,17Wang et al.
18demonstrated that melatonin treatment induces the maintenance of higher contents of GSH through its regulation of the AA-GSH cycle.
Melatonin strongly contributes to AA-GSH cycle.
18In the pres- ent study, we demonstrated that the cell viability was enhanced considerably by the combinational treatment of DHA, GSH and melatonin compared to antioxidant single treatment. In addi- tion, the combinational treatment of DHA, GSH and melatonin reduces markedly the death cells and prevents the cells after OGD/R injury. BBB damage aggravates by reperfusion after ischemia
43,44and the tight junctions as BBB structural compo- nents
45,46disrupts against ischemic injury. Argaw et al. demon- strated that disruption of claudin-5 contribute to BBB damage in various CNS pathological conditions and specifically VEGF me- diates these mechanism.
47Tao et al. demonstrates that melatonin prevents tight junction integrity against ischemic injury by con- firming ZO-1 expression in endothelial cells.
48Our results also indicates that claudin 5 as a tight junction protein was preserved by DHA and GSH and melatonin combination treatment. In this study, we could not determine that melatonin can boost the preventive effect of tight junction protein in combinational treat- ment of DHA and GSH group. However, the preventive effect of tight junction protein is almost the same between the combina- tional treatment of DHA and GSH and the combinational treat- ment of DHA and GSH and melatonin. Moreover, the expres- sion of HIF-1α and VEGF attenuated conspicuously in the combinational treatment of DHA and GSH and melatonin. The antioxidant combinational treatment prevents bEND3. cells against hypoxia injury suggesting that HIF-1α and VEGF induc- es hypoxia gene’s transcription
49and triggers apoptosis pathway and exacerbate damaged region under hypoxia injury.
50There- fore, VEGF induces activation of ERK1/2.
26Slevin et al. demon- strated that increased ERK1/2 phosphorylation was noted in the vulnerable penumbra after acute ischemic stroke in humans and in animal models.
51Also, previous study demonstrated that VEGF activates signaling pathways via the phosphorylation of ERK and inhibition of it also resulted in reduced cell death.
26Al- though VEGF is a potential inducer in angiogenesis, it can have detrimental effects such as an increase in endothelium permea- bility leading to vasogenic edema.
52Zhang and Chopp found that early postis chemic administration of VEGF significantly increased BBB permeability.
53In the present study, findings of western blot shows that the combinational treatment of antioxi-
dants promotes reduction of VEGF expression in spite of the same exposure of hypoxia injury. These findings suggests that the antioxidant combinational treatment reduces the BBB per- meability and the possibility of making vasogenic edema. In ad- dition, by confirming the phosphorylation of ERK1/2, we con- cludes that the combinational treatment of DHA and GSH and melatonin inhibits cell death mechanism through increase of ERK1/2 expression like previous studies.
24,25Like our findings, Narasimhan et al. also demonstrated that pERK1/2 expression decreased after OGD in endothelial cells that had decreased VEGF levels and cell death decreased by blocking VEGF.
26In the present study, we demonstrated that the combinational treat- ment of DHA and GSH and melatonin increases the cell viability and preserves BBB permeability and also inhibits the apoptosis pathway associated with VEGF and ERK1/2. Hence, the combi- national treatment of DHA and GSH and melatonin boosts the preventive effect of bEND3 cells against OGD/R induced injury.
Acknowledgments
This research was supported by the Brain Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Science, ICT & Future Planning (2009-0080364). This work was sup- ported by the Brain Korea 21 Plus Project for Medical Science, Yonsei Uni- versity.
REFERENCES
1. Lei C, Deng J, Wang B, Cheng D, Yang Q, Dong H, et al. Reactive ox- ygen species scavenger inhibits STAT3 activation after transient focal cerebral ischemia-reperfusion injury in rats. Anesth Analg 2011;113:
153-159.
2. Cervantes MI, de Oca Balderas PM, de Jesús Gutiérrez-Baños J, Oro- zco-Ibarra M, Fernández-Rojas B, Medina-Campos ON, et al. Com- parison of antioxidant activity of hydroethanolic fresh and aged garlic extracts and their effects on cerebral ischemia. Food Chem 2013;140:
343-352.
3. Carr AC, Frei B. Toward a new recommended dietary allowance for vitamin C based on antioxidant and health effects in humans. Am J Clin Nutr 1999;69:1086-1107.
4. Padayatty SJ, Katz A, Wang Y, Eck P, Kwon O, Lee JH, et al. Vitamin C as an antioxidant: evaluation of its role in disease prevention. J Am Coll Nutr 2003;22:18-35.
5. Winterbourn CC, Metodiewa D. The reaction of superoxide with re- duced glutathione. Arch Biochem Biophys 1994;314:284-290.
6. Meister A. Glutathione-ascorbic acid antioxidant system in animals. J Biol Chem 1994;269:9397-9400.
7. Wu G, Fang YZ, Yang S, Lupton JR, Turner ND. Glutathione metabo- lism and its implications for health. J Nutr 2004;134:489-492.
8. Beutler E. Abnormalities of the hexose monophosphate shunt. Semin Hematol 1971;8:311-347.
9. Roos D, Weening RS, Voetman AA, van Schaik ML, Bot AA, Meer- hof LJ, et al. Protection of phagocytic leukocytes by endogenous glu- tathione: studies in a family with glutathione reductase deficiency.
Blood 1979;53:851-866.
10. Meister A, Anderson ME. Glutathione. Annu Rev Biochem 1983;52:
711-760.
11. Puskas F, Gergely P Jr, Banki K, Perl A. Stimulation of the pentose phosphate pathway and glutathione levels by dehydroascorbate, the oxidized form of vitamin C. FASEB J 2000;14:1352-1361.
12. Poeggeler B, Saarela S, Reiter RJ, Tan DX, Chen LD, Manchester LC,
et al. Melatonin--a highly potent endogenous radical scavenger and electron donor: new aspects of the oxidation chemistry of this indole accessed in vitro. Ann N Y Acad Sci 1994;738:419-420.
13. Barlow-Walden LR, Reiter RJ, Abe M, Pablos M, Menendez-Pelaez A, Chen LD, et al. Melatonin stimulates brain glutathione peroxidase ac- tivity. Neurochem Int 1995;26:497-502.
14. Lim C, Alexander MP, LaFleche G, Schnyer DM, Verfaellie M. The neurological and cognitive sequelae of cardiac arrest. Neurology 2004;
63:1774-1778.
15. Sachdev PS, Brodaty H, Valenzuela MJ, Lorentz LM, Koschera A.
Progression of cognitive impairment in stroke patients. Neurology 2004;63:1618-1623.
16. Ginsberg MD, Busto R. Rodent models of cerebral ischemia. Stroke 1989;20:1627-1642.
17. Hartman RE, Lee JM, Zipfel GJ, Wozniak DF. Characterizing learning deficits and hippocampal neuron loss following transient global cere- bral ischemia in rats. Brain Res 2005;1043:48-56.
18. Wang P, Yin L, Liang D, Li C, Ma F, Yue Z. Delayed senescence of ap- ple leaves by exogenous melatonin treatment: toward regulating the ascorbate-glutathione cycle. J Pineal Res 2012;53:11-20.
19. Abbott NJ. Astrocyte-endothelial interactions and blood-brain barri- er permeability. J Anat 2002;200:629-638.
20. Wolburg H, Lippoldt A. Tight junctions of the blood-brain barrier:
development, composition and regulation. Vascul Pharmacol 2002;
38:323-337.
21. Kaur C, Ling EA. Blood brain barrier in hypoxic-ischemic conditions.
Curr Neurovasc Res 2008;5:71-81.
22. Semenza G. Signal transduction to hypoxia-inducible factor 1. Bio- chem Pharmacol 2002;64:993-998.
23. Zhang ZG, Zhang L, Jiang Q, Zhang R, Davies K, Powers C, et al.
VEGF enhances angiogenesis and promotes blood-brain barrier leak- age in the ischemic brain. J Clin Invest 2000;106:829-838.
24. Kauppinen TM, Chan WY, Suh SW, Wiggins AK, Huang EJ, Swanson RA. Direct phosphorylation and regulation of poly(ADP-ribose) polymerase-1 by extracellular signal-regulated kinases 1/2. Proc Natl Acad Sci U S A 2006;103:7136-7141.
25. Stanciu M, Wang Y, Kentor R, Burke N, Watkins S, Kress G, et al. Per- sistent activation of ERK contributes to glutamate-induced oxidative toxicity in a neuronal cell line and primary cortical neuron cultures. J Biol Chem 2000;275:12200-12206.
26. Narasimhan P, Liu J, Song YS, Massengale JL, Chan PH. VEGF Stim- ulates the ERK 1/2 signaling pathway and apoptosis in cerebral endo- thelial cells after ischemic conditions. Stroke 2009;40:1467-1473.
27. Jung HJ, Jeon YH, Bokara KK, Koo BN, Lee WT, Park KA, et al. Ag- matine promotes the migration of murine brain endothelial cells via multiple signaling pathways. Life Sci 2013;92:42-50.
28. Dayoub JC, Ortiz F, LÓpez LC, Venegas C, Del Pino-Zumaquero A, Roda O, et al. Synergism between melatonin and atorvastatin against endothelial cell damage induced by lipopolysaccharide. J Pineal Res 2011;51:324-330.
29. Yang MZ, Mun CH, Choi YJ, Baik JH, Park KA, Lee WT, et al. Agma- tine inhibits matrix metalloproteinase-9 via endothelial nitric oxide synthase in cerebral endothelial cells. Neurol Res 2007;29:749-754.
30. Bokara KK, Kwon KH, Nho Y, Lee WT, Park KA, Lee JE. Retroviral expression of arginine decarboxylase attenuates oxidative burden in mouse cortical neural stem cells. Stem Cells Dev 2011;20:527-537.
31. Li HL, Kang YM, Yu L, Xu HY, Zhao H. Melatonin reduces blood pressure in rats with stress-induced hypertension via GABAA recep- tors. Clin Exp Pharmacol Physiol 2009;36:436-440.
32. Kong X, Li X, Cai Z, Yang N, Liu Y, Shu J, et al. Melatonin regulates the viability and differentiation of rat midbrain neural stem cells. Cell Mol Neurobiol 2008;28:569-579.
33. Rubin LL, Staddon JM. The cell biology of the blood-brain barrier.
Annu Rev Neurosci 1999;22:11-28.
34. Wang Y, Peng KM, Li JL, Song H, Li SH, Wei L, et al. Ultrastructure and melatonin 1a receptor distribution in the ovaries of African os- trich chicks. Cytotechnology 2008;56:187-195.
35. Xia CM, Shao CH, Xin L, Wang YR, Ding CN, Wang J, et al. Effects of melatonin on blood pressure in stress-induced hypertension in rats. Clin Exp Pharmacol Physiol 2008;35:1258-1264.
36. Li ZR, Shen MH, Niu WM. [Involvement of melatonin in the adjust- ing effect of electroacupuncture in resisting oxygen stress in cerebral ischemia-reperfusion injury rats]. Zhen Ci Yan Jiu 2008;33:164-168.
37. Li L, Zhang B, Tao Y, Wang Y, Wei H, Zhao J, et al. DL-3-n-bu- tylphthalide protects endothelial cells against oxidative/nitrosative stress, mitochondrial damage and subsequent cell death after oxygen glucose deprivation in vitro. Brain Res 2009;1290:91-101.
38. Cavdar Z, Egrilmez MY, Altun ZS, Arslan N, Yener N, Sayin O, et al.
Resveratrol reduces matrix metalloproteinase-2 activity induced by oxygen-glucose deprivation and reoxygenation in human cerebral microvascular endothelial cells. Int J Vitam Nutr Res 2012;82:267-274.
39. May JM, Qu ZC, Neel DR, Li X. Recycling of vitamin C from its oxi- dized forms by human endothelial cells. Biochim Biophys Acta 2003;1640:153-161.
40. May JM. How does ascorbic acid prevent endothelial dysfunction?
Free Radic Biol Med 2000;28:1421-1429.
41. Vera JC, Rivas CI, Fischbarg J, Golde DW. Mammalian facilitative hexose transporters mediate the transport of dehydroascorbic acid.
Nature 1993;364:79-82.
42. Daskalopoulos R, Korcok J, Tao L, Wilson JX. Accumulation of intra- cellular ascorbate from dehydroascorbic acid by astrocytes is de- creased after oxidative stress and restored by propofol. Glia 2002;39:
124-132.
43. del Zoppo GJ, von Kummer R, Hamann GF. Ischaemic damage of brain microvessels: inherent risks for thrombolytic treatment in stroke. J Neurol Neurosurg Psychiatry 1998;65:1-9.
44. Simard JM, Kent TA, Chen M, Tarasov KV, Gerzanich V. Brain oede- ma in focal ischaemia: molecular pathophysiology and theoretical implications. Lancet Neurol 2007;6:258-268.
45. Mark KS, Davis TP. Cerebral microvascular changes in permeability and tight junctions induced by hypoxia-reoxygenation. Am J Physiol Heart Circ Physiol 2002;282:H1485-H1494.
46. Traweger A, Fang D, Liu YC, Stelzhammer W, Krizbai IA, Fresser F, et al.
The tight junction-specific protein occludin is a functional target of the E3 ubiquitin-protein ligase itch. J Biol Chem 2002;277:10201-10208.
47. Argaw AT, Gurfein BT, Zhang Y, Zameer A, John GR. VEGF-mediat- ed disruption of endothelial CLN-5 promotes blood-brain barrier breakdown. Proc Natl Acad Sci U S A 2009;106:1977-1982.
48. Tao RR, Huang JY, Shao XJ, Ye WF, Tian Y, Liao MH, et al. Ischemic injury promotes Keap1 nitration and disturbance of antioxidative re- sponses in endothelial cells: a potential vasoprotective effect of mela- tonin. J Pineal Res 2013;54:271-281.
49. Agrawal M, Kumar V, Singh AK, Kashyap MP, Khanna VK, Siddiqui MA, et al. trans-Resveratrol protects ischemic PC12 Cells by inhibit- ing the hypoxia associated transcription factors and increasing the levels of antioxidant defense enzymes. ACS Chem Neurosci 2013;4:
285-294.
50. Li YN, Pan R, Qin XJ, Yang WL, Qi Z, Liu W, et al. Ischemic neurons activate astrocytes to disrupt endothelial barrier via increasing VEGF expression. J Neurochem 2013. [Epub ahead of print]
51. Slevin M, Krupinski J, Slowik A, Rubio F, Szczudlik A, Gaffney J. Ac- tivation of MAP kinase (ERK-1/ERK-2), tyrosine kinase and VEGF in the human brain following acute ischaemic stroke. Neuroreport 2000;11:2759-2764.
52. Bates DO, Harper SJ. Regulation of vascular permeability by vascular endothelial growth factors. Vascul Pharmacol 2002;39:225-237.
53. Zhang Z, Chopp M. Vascular endothelial growth factor and angio- poietins in focal cerebral ischemia. Trends Cardiovasc Med 2002;12:
62-66.