내열성 바이오플라스틱 생산을 위한 미생물 촉매 개발
오 영 훈ㆍ이 승 환†ㆍ박 시 재*,†
한국화학연구원 바이오화학연구센터, *명지대학교 환경에너지공학과
Microbial Catalysts for the Production of Thermo-resistant Bioplastics
Young Hoon Oh, Seung Hwan Lee
†, and Si Jae Park*
,†*Institute of Marine Science and Technology Research, Hankyong National University, Anseong 456-749, Korea
†Department of Chemical Engineering, Hankyong National University, Anseong 456-749, Korea
Abstract: 온실가스축적으로 인한 지구온난화를 비롯한 기후변화 및 고갈되어가는 석유를 비롯한 화석원료에 대한 문제를 해결하기 위해 재생가능한 자원으로부터 바이오기반 케미칼 및 고분자 등의 화학제품을 생산하는 바이오화학 공정이 많은 관심을 받고 있다. 본 기고문에서는 바이오화학공정에 핵심적인 촉매로 사용되고 있는 재조합 미생물 및 효소의 최근 개발동향을 내열성 엔지니어링 플라스틱인 바이오나일론의 생산을 위하여 개발되고 있는 바이오촉매 를 중심으로 살펴보고자 한다.
Keywords: Biocatalyt, Biorefinery, Enzyme, Recombinant microorganism, Nylon, aminocarboxylic acid, diamine
1. 서 론
1)
이산화탄소와 같은 온실가스의 대기 중의 축적 으로 인한 지구온난화 문제와 전 세계 산업의 핵 심원료인 석유자원 고갈문제를 해결하기 위하여 다양한 연구가 진행되고 있다. White Biotechnol- ogy 또는 Industrial Biotechnology라고도 불리는 바이오화학기술은 바이오기술(효소, 전세포등의 생촉매)과 화학기술을 접목한 기술로서, 케미칼 및 플라스틱을 포함한 각종 화학소재, 바이오연료 를 포함한 에너지 등의 기존 화학산업에 대한 대 체 및 보완의 파급효과가 매우 큰 미래형 기술이 기 때문에 현재의 화학산업을 지속적으로 성장시 킬 수 있는 대안으로 여겨지고 있다.
또한, 바이오화학기술은 바이오매스와 같은 재 생가능한 자원과 화학산업용 바이오촉매를 사용 하므로 환경오염과 에너지 소모를 최소화하기 때 문에 이를 기반으로 하는 바이오화학산업 및 바이
저자 (E-mail: [email protected], [email protected])
오리파이너리의 중요성은 나날이 관심을 받고 있 는 상황이다. Figure 1에서 나타난 바와 같이, 바이 오화학산업의 공정이나 밸류체인이 석유화학산업 과 거의 유사하므로, 기존의 석유화학플랫폼을 바 탕으로 산업을 수행하며, 석유에 의존하는 기존의 석유화학산업을 보완할 수 있어 석유화학 규모 세 계 5위이면서 수송용 및 화학산업원료를 전적으로 수입에 의존하는 국내에 미치는 파급효과는 상당 하다고 할 수 있다.
최근 세계경제포럼이 다음과 같이 바이오매스 를 이용한 바이오리파이너리를 구성하고 있는 바 이오케미칼 플랫폼(biochemical platform), 열화학 플랫폼(thermochemical platform), 미생물 플랫폼 (Microorganism platform), 열과 에너지 융합 플랫 폼(Combined Heat & Power platform)의 총 4개의 플랫폼 기술을 제시하였는데[1], 본 기고문에서는 이 4가지 플랫폼 중 바이오케미칼 및 미생물 플랫 폼을 이용한 물질 중 최근에 많은 관심을 받고 있 는 바이오나일론의 생산을 다루고자 한다.
바이오케미칼 및 미생물 플랫폼 공정에는 미생
Figure 1. 바이오화학산업 공정과 밸류체인.
물 혹은 효소 등의 바이오촉매가 공정의 핵심촉매 로 투입되기 때문에, 바이오화학산업의 성공을 위 해서는 공정에 투입되는 핵심적인 촉매인 미생물 호스트의 목적산물을 생산하는 능력이 매우 중요 하고, 미생물 호스트의 목적산물을 생산하는 효율 및 성능이 전체공정의 산업적 타당성 및 효율성에 가장 큰 영향을 미친다. 성능이 우수한 미생물 호 스트 촉매의 개발을 위하여 생화학, 미생물학, 유 전체학, 단백체학, 생물정보학 등의 지식을 기반으 로 생물공학 기술이 활발하게 응용되고 있다. 바 이오화학산업은 정밀화학, 의약품 뿐 아니라 고분 자, 신소재, 그리고 범용화학제품의 생산에까지 이 용되고 있으며, 정밀화학⋅플라스틱⋅종이⋅섬유
⋅식품⋅제약 등의 매우 다양한 제품에 대해, 현 재는 DuPont, Monsanto, Cargill, Dow, Genen- cor, Maxygen, Diversa, DSM 등 여러 기업들이 1,3-propanediol, butanol, diamines, aminocabo- xylic acids, 1,4-butanediol, 3-hydroxypropionic acid, succinic acid, polylactic acid (PLA), poly- hydroxyalkanoates (PHAs), bionylon 등의 바이오 기반 화학소재를 상업화하는데 많은 관심을 보이 고 있다.
본 기고문에서는 현재 큰 관심을 받고 있는 바 이오플라스틱의 핵심적인 촉매로 적용되고 있는 미생물 호스트의 개발에 관한 최근 동향을 논의하 고자 한다.
2. 바이오플라스틱
바이오플라스틱은 식물 등의 바이오매스로부터 얻어지거나, 미생물을 이용한 발효공정을 통하여 생산되는 플라스틱을 말한다. 바이오플라스틱은 원료물질과 생산공정에 따라 크게 세 가지 그룹으 로 나눌 수 있다.
첫째 그룹에 해당하는 바이오플라스틱은 바이오
매스를 원료로 활용하여, 바이오 촉매인 미생물을
통해서만 생산되는 고분자로서 polyhydroxyalkanoate
(PHA)과 poly(γ-glutamic acid) (PGA)가 첫째 그
룹의 대표적인 예이며, 이와 같은 고분자는 당이
나 oil 등의 탄소원을 미생물이 대사하여 생성되는
대사산물로서 미생물 내에 축적되거나, 밖으로 배
출되어 배양액에 존재한다. 고분자를 구성하고 있
는 단량체와 합성에 요구되는 효소 등은 모두 미
생물에 원래 존재하는 대사회로나 개량된 대사회
로를 통해 직접 생산되고 제공된다.
둘째 그룹에 해당하는 고분자는 현재 가장 널리 사용되는 PLA와 같이 산업적으로 생산되고 있는 대부분의 바이오플라스틱이 해당되는 그룹으로 바이오기술과 화학기술이 모두 적용된 하이브리 드 공정을 통해 합성되는 고분자이다. 고분자의 전체 혹은 일부분의 단량체, 혹은 고분자 단량체 의 전구체를 미생물의 발효나 생물전환을 통해 생 산한 후 이들을 후속 화학공정을 통해 고분자를 합성한다. poly(butylene succinate) (PBS), poly (trimethylene terephthalate) (PTT), PLA, nylon 4 등이 이 그룹에 속하며 PTT는 1, 3-propanediol을 미생물 발효 공정으로 생산한 후 또 다른 단량체 인 석유화학공정 유래인 terephthalate과 중합하여 합성된다. PLA와 nylon 4는 미생물 발효에 의해 생산된 lactic acid와 glutamic acid를 전구체로 사 용하여 lactide, 2-pyrrolidone 등의 고분자의 단량 체를 합성후 중합을 통하여 고분자를 생산한다.
셋째 그룹은 온전히 화학공정을 통해서만 합성 이 되는데, 바이오매스를 원료로 한 화학공정을 통해 생산된 고분자 단량체의 화학합성을 통해 고 분자를 합성하게 된다. Nylon 5, 10과 nylon 6, 10 이 대표적인 예이며, castor oil의 화학처리를 통해 얻어진 sebacic acid가 대표적인 단량체로 이용된 다. 최근에 nylon 5, 10의 단량체인 cadaverine 또 한 바이오공정을 통해 생산되는 공정이 개발되고 있는 추세를 감안하면, 바이오플라스틱의 생산공 정에서 바이오촉매가 차지하고 있는 비율은 매우 커지고 있는 것을 알 수 있다.
3. 바이오나일론
다양한 바이오플라스틱 중 현재 상업화가 활발 하게 이루어지고 있는 고분자는 PLA, PBS, PTT 를 포함한 폴리에스터 제품들이 있으나, 아직까지 생산되는 바이오플라스틱은 종류와 양에 있어서 현저히 부족한 상태이며 앞으로도 많은 연구와 개 발이 필요한 실정이다. 바이오나일론은 내열성 및 기계적 강도가 뛰어난 엔니지어링 플라스틱으로
서의 응용이 가능한 플라스틱이기 때문에 최근에 열적, 기계적 강도가 약한 바이오고분자의 한계를 극복할 수 있는 고분자로서 관심을 받고 있다. 나 일론 4, 나일론 5, 나일론 6와 같이 피롤리돈 (pyrrolidone), 발레로락탐(valerolactame), 카프로 락탐(caprolactame) 등의 aminocarboxylic acid 유 래 락탐을 단량체로 사용하거나 퓨트레신(putre- scine), 카다베린(cadaverine) 등의 다이아민(dia- mine)을 단량체로 사용하는 바이오나일론 합성공 정은 크게 나일론 단량체로 사용될 수 있는 ami- nocarboxylic acid, 다이아민을 생산하는 미생물공 정과 미생물공정으로부터 생산된 aminocarboxylic acid를 이용하여 락탐을 생산한 후 생산된 락탐을 단량체로 사용하여 나일론을 합성하는 화학공정 으로 이루어져 있다. 다이아민을 단량체로 사용할 때에는 미생물공정 혹은 화학공정으로부터 생산 된 succinic acid, adipic acid, sebacic acid 등의 다 이에시드(diacid)를 이용하여 나일론을 합성하게 된다.
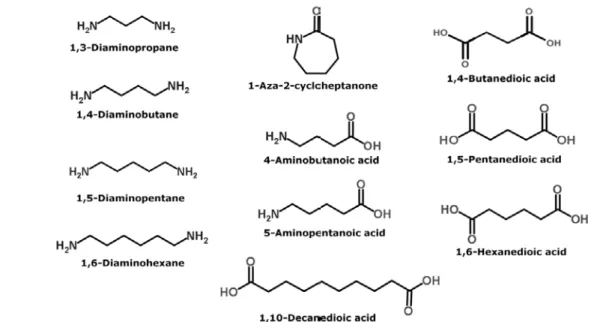
Figure 2에는 나일론합성에 사용 가능한 아미노 산, 락탐, 다이아민, 다이에시드 단량체를 나타내 었다. 효율적인 바이오나일론 생산공정을 개발하 기 위해서는 미생물공정으로 생산된 나일론 단량 체를 이용하여 화학적으로 고분자를 합성할 수 있 도록 바이오-화학 융합공정을 디자인 하는 것이 매우 중요한데, 이를 위하여 단량체의 전구체로 사용되는 aminocarboxylic acid, 다이아민을 생산 하는 미생물 프로세스의 개발이 필요하며, 공정 전반에서 가장 중요한 영향을 끼치는 미생물 균주 의 개량은 필수적이다.
3.1. Whole cell biocatalyst를 이용한 나일론 단량체의 생산
현재 활발하게 연구되고 있는 나일론 단량체인
aminocarboxylic acid는 탄소수 4개인 gamma
aminobutyric acid (GABA), 탄소수 5개인 5-ami-
novaleric acid, 탄소수 6개인 6-aminohexanoic
acid로서, GABA의 경우 전구체로 아미노산인 글
루탐산(glutamic acid)을 사용하여 생산되며 5-
Figure 2. 나일론의 단량체로 사용되는 케미칼.
Aminovaleric acid, 6-aminohexanoic acid는 전구 체로 라이신(lysine)을 사용하여 생산된다. amino- carboxylic acid는 전구체로 아미노산을 사용하기 때문에 글루탐산, 라이신과 같은 아미노산 전구체 를 원료로 사용하여 직접 aminocarboxylic acid로 전환하는 미생물 혹은 효소공정과 글루코스 등과 같은 탄소원을 이용하여 글루탐산 혹은 라이신과 같은 아미노산 전구체를 생산하고 이를 이용하여 aminocarboxylic acid로 전환하는 미생물 직접생 산공정과 같이 생산방법이 크게 두 가지로 나뉠 수 있다.
3.1.1 Gamma aminobutyric acid (GABA) GABA는 탄소수 4개인 아미노산으로서 단백질 합성에 사용되지는 않지만 사람의 뇌, 눈, 신경계, 혈액에 많이 분포하고 있다고 밝혀져 있다[2].
GABA는 뇌의 활성을 향상시키는 효과가 있어, 뇌의 활성 및 긴장완화 등을 유도하는 식품이나 의약품에 첨가되어 사용되고 있다. GABA의 상업 적 생산은 주로 글루탐산의 미생물 혹은 효소전환 을 통하여 생산되고 있는데, 이는 미생물을 이용 하여 글루코스로부터 직접 GABA를 생산하는 방
법은 비교적 비효율적이기 때문이다. GABA의 생
산을 위해서 글루탐산 혹은 글루탐산의 염형태인
글루탐산나트륨(Glutamate sodium salt, MSG)을
GABA 생산의 직접적인 전구체로 사용하는 기술
이 활발하게 개발되고 있으며, 핵심적인 효소로
글루탐산에서 이산화탄소를 제거하여 GABA를
만드는 글루탐산탈산화효소(Glutamate decarbo-
xylase, GAD)가 사용되고 있다. GAD의 최적의
반응 pH는 4.5 근처이며, 특이하게도 pH가 상승
함에 따라서 효소활성이 점진적으로 낮아지는 것
이 아니라 활성을 거의 잃어버리기 때문에, 반응
pH를 최적인 4.5 근처로 유지하는 것이 매우 중요
하다. 따라서, 최적의 반응 pH인 4.5 근처로 유지
하기 위해서는 일반적으로 염산, 황산, 초산 등과
같은 산을 반응액에 투입하게 된다. 하지만, 글루
탐산이 GABA로 전환되면서 반응액의 pH가 지속
적으로 상승하기 때문에 높은 농도의 글루탐산을
GABA로 전환하기 위해서는 많은 양의 산을 반응
액에 투입해야 한다. 이러한 반응에서 생성된 염을
제거하기 위하여 이용되는 이온교환수지 공정, 재
결정과 같은 정제공정은 GABA 생산단가를 높이
는 주원인이다.
이러한 문제를 해결하기 위해 한국의 바이오벤 처기업인 엠에이치투 바이오케미칼(MH2)은 글루 탐산을 GAD를 이용하여 GABA로 전환하는 반응 에 pH를 조절하기 위하여 일반적으로 사용되는 산 대신에 글루탐산을 도입하는 공정을 개발하였다 [3]. 글루탐산을 GABA의 전구체로 사용함으로써, 반응액의 pH 또한 글루탐산으로 조절하는 방법을 적용하여 염의 발생을 최소화할 수 있었다. MSG 를 사용하여 GABA를 생산하였을 때에는 GABA 단위 중량당 투입된 염산의 양이 0.35로 매우 높 았으나, 글루탐산을 사용하였을 때에는 그 값이 0.018로 매우 낮아졌다. 글루탐산을 기질로 사용 하였을 때에 생산된 GABA의 최종농도는 240 g/L 에 달했다. 최근에 양이온교환수지를 이용한 MSG를 기질로 사용하면서 염의 생성을 최소화하 는 GABA 생산공정이 보고되었다. 이 반응에서 양이온교환수지는 반응액의 pH를 떨어뜨리는 고 체산으로 사용되어 염의 생성을 억제하는 효과를 보였다.
Dinh 등은 0.2 M sodium acetate buffer (pH 4.6)에서 1 M의 MSG를 GAD를 이용하여 GABA 로 전환하는 반응 중에 양이온교환수지를 투입하 였으며, 이 반응을 MSG의 GABA로의 전환율을 13%에서 67%로 향상시킨 결과를 발표하였다[4].
또한 sodium acetate buffer 대신 물을 반응액으로 사용하였을 때에도 MSG의 전환율이 크게 향상됨 을 보여 양이온교환수지가 반응도중의 pH향상을 억제하여 반응수율을 향상시키는 것을 확인하였 다[4]. GAD를 이용한 효소반응공정은 GAD효소 의 분리정제가 어려워 촉매로 사용되는 효소의 생 산가격이 비교적 높으며, 효소반응에 피리독살 5'- 포스페이트(pyridoxal 5'-phosphate, PLP)와 같은 조효소가 필요하기 때문에 GABA의 대량생산을 통한 상업화에 어려움이 있을 수 있다. 따라서 이 러한 점을 보완하기 위해서 바이오촉매로서 GAD 를 효율적으로 발현하는 자연 미생물이나 재조합 미생물을 이용하여 MSG 혹은 글루탐산을 전환하 여 GABA를 생산하는 기술이 개발되고 있다.
GAD를 발현하는 자연미생물 중 lactic acid
bacteria (LAB)가 효율적으로 MSG를 GABA로 전환하는 것으로 알려져 있으며, Lactobacillus brevis NCL912 균주의 경우에는 유가식 배양을 통해 1 M의 GABA를 생산하는 것으로 보고되었 다[5]. LAB 이외에 GABA 생산을 위해서 GAD를 발현하는 재조합 대장균이 활발하게 연구되고 있 다. Le Vo 등은 대장균 유래 GAD, gluta- mate/GABA antiporter (GadC), GABA amino- transferase (GabT) 등이 GABA 생산시 어떤 영향 을 미치는지에 대해 재조합 대장균을 이용하여 규 명하였으며, GabT는 생성된 GABA를 이용하여 succinic semialdehyde를 만드는 효소이기 때문에 셀에서 제거하는 것이 GABA의 생산에 유리한 것을 보고하였다. 대장균 gabT 뮤턴트 균주에서 GAD 와 GadC를 동시에 발현하였을 때 10 g/L의 MSG 로부터 5.46 g/L의 GABA가 생성되었으며 MSG 의 GABA로의 전환수율은 89.5%에 달했다[6].
대장균 유래의 GAD 이외에도 Neurospora cras- sa OR74A 유래 GAD를 발현하는 재조합 대장균 또한 5.25 g/L의 GABA를 10 g/L의 MSG로부터 86.2%의 전환수율로 생성되었다는 보고는 다양한 유래의 GAD가 GABA전환 재조합 균주 개발에 이용될 수 있음을 의미한다[7].
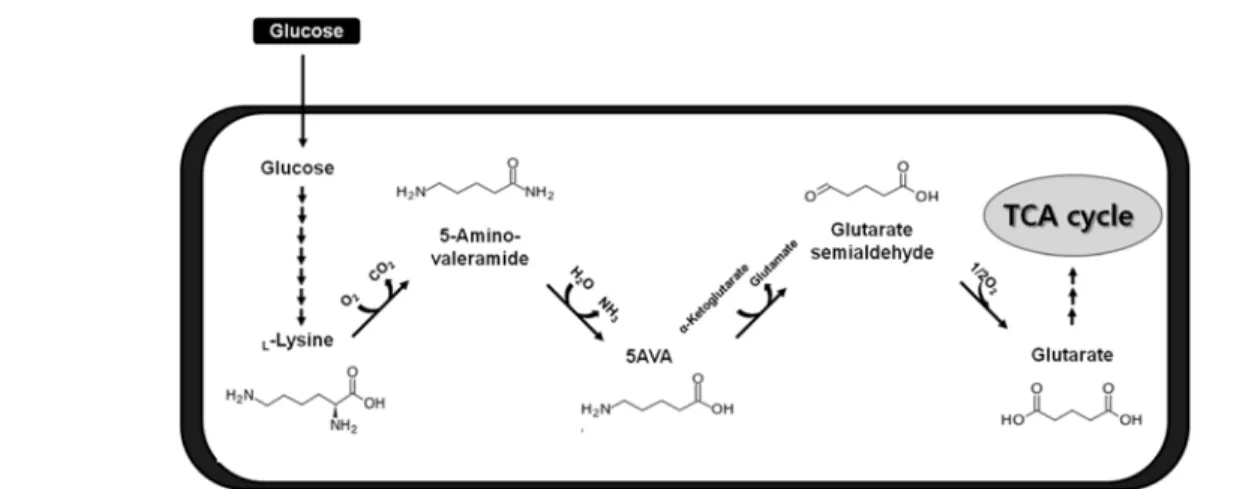
3.1.2. 5-aminovaleric acid (5AVA)
5-aminovaleric acid (5AVA)는 Pseudomonas
putida의 라이신(L-lysine) 분해 대사회로의 중간대사산물로서, glutarate, 5-hydroxyvalerate, 1,5-
pentanediol, valerolactam 등의 다양한 C5 케미칼
의 플랫폼 케미칼로서 사용될 가능성이 매우 높은
물질이다. 이외에도 nylon 5와 같은 바이오나일론
의 단량체로도 사용이 가능하기 때문에 최근 관심
을 받고 있다. P. putida에서 5AVA는 lysine
2-monooxygenase (DavB) 와 delta-aminovaleramidase
(DavA)의 두 효소에 의해서 L-lysine으로부터 만
들어지지만, P. putida의 aminovalerate 대사회로
에 의해 5AVA는 glutarate로 전환된 후 TCA cy-
cle을 이용하여 미생물 대사에 사용되기 때문에
자연적인 P. putida 균주에서의 5AVA 생성은 적은
Figure 3. 글루코스로부터 5-aminovalerate 및 glutarate의 생산.
편이다. 따라서, aminovalerate 대사회로를 보유하지 않은 미생물을 호스트균주로 사용하여 5AVA생산 을 하는 것이 효율적으로 생각되어 재조합 대장균 을 이용한 5AVA 생산이 시도되고 있다(Figure 3).
P. putida 유래 lysine 2-monooxygenase (DavB)
와 delta-aminovaleramidase (DavA)를 발현하는 재조합 대장균 WL3110은 10 g/L의 L-lysine으로 부터 3.6 g/L의 5AVA를 생산하였으며, 5AVA aminotransferase (GabT), glutarate semialdehyde dehydrogenase (GabD)를 DavA, DavB와 동시에 발현한 결과 10 g/L의 L-lysine으로부터 1.7 g/L의 glutarate을 생산할 수 있었다[8].
3.1.3. Cadaverine
Cadaverine (카다베린)은 탄소수가 5개인 다이 아민으로서 나일론 510, 512 등 다양한 나일론의 단량체로 사용될 수 있어 산업계의 많은 관심을 끌어 왔다. 특히, cadaverine은 Lysine decarboxy- lase(LDC)에 의한 L-lysine의 decarboxylation 반 응에 의하여 생성될 수 있기 때문에 이미 산업적 으로 대량생산되고 있는 L-lysine을 활용할 수 있 는 새로운 화학제품으로 각광받고 있다. 현재까지 대장균 유래의 LDC인 CadA와 LdcC를 이용한 다 양한 연구가 보고되어 있는데, 미생물 기반의 카 다베린의 생산은 대부분 대장균의 CadA와 LdcC 를 이용하여 진행되어 왔다[9]. 이 중 CadA는
LdcC보다 먼저 발견이 되어 사용되었으나, 활성 과 pH의 스펙트럼 등의 약점 등을 보유하고 있다 [9,10]. 반면 LdcC는 보다 넓은 pH에서 활성을 갖 는다는 장점을 갖고 있지만 promoter가 약해 자연 적인 발현이 제한된다는 단점을 갖는다[11]. 일본 의 아지노모토(Ajinomoto), 토레이(Toray)와 독일 의 BASF 등을 중심으로 Whole cell biocatalyst를 이용한 카다베린의 생산이 다양하게 연구되어 왔 다. 대장균 유래 CadA를 과발현하는 재조합 대장 균을 연속적인 계대배양을 통해 고농도로 배양하 고 최종적으로 50 L 발효기에서 22 L의 배양액에 서 수확한 균주를 이용해 3.1 kg의 L-lysine으로부 터 2.2 kg의 카다베린을 100% (mol/mol)의 수율 로 생산할 수 있었다[12].
3.2. 당을 탄소원으로 이용하여 나일론 단량체
를 직접 생산하는 재조합 미생물의 개발
재조합 미생물을 이용한 글루코스로부터
GABA를 직접 생산하는 방법은 글루탐산을 효율
적으로 생산하는 Corynebacterium 균주를 호스트
균주로 사용하여 개발이 되고 있지만, 글루탐산의
직접전환에 비해 효율이 낮기 때문에 개발이 더
필요하다. 5AVA를 글루코스로부터 직접 생산하
기 위해서 라이신의 생산회로가 강화되어 글루코
스로부터 라이신 생산능이 강화된 재조합 대장균
균주에 DavA와 DavB를 발현한 결과 0.27 g/L와
Figure 4. 바이오기술을 활용한 cadaverine의 생산.
0.5 g/L의 5AVA가 회분식 배양과 유가식 배양을 통해 글루코스로부터 생산될 수 있었다[8].
또한, 최근에 나일론의 단량체인 다이아민을 글 루코스로부터 직접 생산하는 재조합 대장균이 보 고되었다[13,14]. 대장균의 대사회로의 개량을 통 해 탄소수가 4개인 퓨트레신과 탄소수가 5개인 카 다베린을 생산할 수 있는 재조합 대장균을 만들 수 있었고, 최종적으로 유가식 배양방법의 개발을 통해 퓨트레신과 카다베린을 24.2 g/L와 9.61 g/L 의 농도로 생산할 수 있었다[13,14].
한편, 카다베린의 전구체인 L-lysine을 과생산 하는 Corynebacterium 균주를 이용한 카다베린의 생산도 시도되고 있다. Lysine 산업에서 사용되는 Corynebacterium 균주의 경우 글루코스로부터 높 은 수율과 농도로 L-lysine을 생산할 수 있기 때문 에 글루코스로부터 카다베린을 생산하는 경우에 재조합 대장균보다 가능성이 높은 것으로 판단되고 있다. 하지만 현재까지 보고된 Corynebacterium 균주를 이용한 카다베린 생산의 경우, 생산된 카 다베린의 농도 및 수율이 10 g/L 이하로 재조합 대장균에 비해서도 높다고 볼 수 없는데 이는 실 제 산업계에서 사용되는 L-lysien 과생산용 돌연
변이 균주를 획득하기가 어렵기 때문인 것으로 보 인다. 즉, L-lysine 생산능이 없는 야생형 균주로부 터 시작하여 이를 대사공학적으로 개량하여 L-ly- sine과 카다베린을 생산하도록 하였기 때문에 높 은 농도로 카다베린을 생산할 수 없었던 것이다.
실제로 독일의 BASF사에서 등록한 특허에 따르 면, 산업용 Corynebacterium 균주를 개량하여 최 종적으로 72.0 g/L의 카다베린을 생산한 것으로 보고되었다[15]. 하지만 최근에는 산업용 Corynebacterium 균주와 야생형 균주의 whole ge- nome sequence를 비교하여 최소한의 대사공학적 개량을 통해 L-lysine을 과생산하는 재조합 Corynebacterium 균주도 보고되고 있기 때문에 가 까운 시일 내에 야생형 Corynebacterium 균주로부 터 카다베린 과생산 균주를 제작할 수 있을 것으 로 기대되고 있다[16,17].
한편, 목질계 바이오매스를 구성하는 주된 탄소
원 중 하나인 자일로스로부터 고농도의 카다베린
을 생산한 연구가 보고되었다. 자일로스 대사능이
강화된 변이 Corynebacterium 균주의 유가식 배양
을 통해 최종적으로 103 g/L의 카다베린을 생산할
수 있었다[18].
Figure 5. 바이오나일론4 생산을 위한 바이오-화학 융합공정.
Figure 4에 현재 보고된 글루코스로 및 자일로 스로부터 생산된 카다베린의 농도를 비교하였다.
당으로부터 GABA, 5AVA, 다이아민 등 나일론의 전구체를 생산하는 것은 바이오플라스틱 및 엔지 니어링 플라스틱 산업에 커다란 반향을 일으킬 수 있을 것으로 기대되나 현재는 수율 및 생산농도가 낮은 상태이기 때문에 보다 합리적이고 효과적인 균주의 개량을 통해 생산효율의 향상을 이루어야 하는 상태이다.
3.3. 바이오공정으로부터 생산된 나일론 단량체 를 이용한 바이오나일론의 합성
재조합 미생물을 이용하여 생산된 나일론모노 머가 바이오화학산업에 적용되기 위해서는 고분 자화학합성공정에 성공적으로 이용되어야 한다.
최근에 바이오공정유래 GABA를 이용한 나일론 4 합성이 보고되어 바이오매스 기반 바이오나일론 에 대한 기대가 높아졌다. GAD를 발현하는 재조 합 대장균을 유가식 배양을 통해서 고농도로 배양 한 다음 GABA 생산을 위한 생촉매로 사용하여 200 g/L의 고농도 MSG를 76.2 g/L의 GABA로 전환하는 기술이 개발되었다. 이렇게 생산된
GABA는 Al
2O
3를 촉매로 사용하여 톨루엔에서 화학반응을 이용하여 96%의 전환율로 2-피롤리돈 (2-pyrrolidone)으로 전환될 수 있었고, 2-피롤리돈 을 이용하여 분자량이 200,000~330,000인 나일론 4를 합성할 수 있었다(Figure 5)[25].
최근에 필자의 그룹에서는 GABA의 대량생산 방법과 비슷한 기술을 이용하여 5AVA를 라이신 으로부터 효율적으로 생산하는 기술을 개발하여, 5AVA 또한 나일론모노머로서 성공적으로 사용될 수 있다는 보고를 준비하고 있다. 또한, 바이오매 스 유래의 원료를 이용하여, 바이오기술 기반의 cadaverine과 sebacic acid를 생산하고 이를 활용 하여 100% 바이오유래의 단량체로 이루어진 Nylon 5, 10의 합성기술을 개발하고 있다. 이러한 연구를 통해 바이오공정과 화학공정이 융합된 하 이브리드 공정을 이용하여 석유화학유래 고분자 를 대체할 수 있는 바이오매스 유래 고분자를 생 산할 수 있는 가능성이 매우 높음을 알 수 있다.
4. 맺음말
바이오케미칼, 바이오연료, 바이오고분자 등을
생산하기 위한 바이오화학기술은 바이오매스를 원료로 하고, 바이오기술을 활용한다는 측면에서 기존의 석유화학과 차별되나, 화학산업에서 보유 하고 있는 인프라를 얼마나 활용하는가가 바이오 화학산업의 성공적인 산업화의 척도가 될 수 있으 며, 이는 생산물질의 생산성, 물성 등이 현재의 공 정에 적용되는 석유화학산업 유래의 물질들과 동 등하거나 우수해야 함을 의미한다. 이를 위해서는 바이오공정에 핵심촉매로 사용되는 미생물 호스 트 균주 및 효소의 우수성이 확보되어야만 하며, 최근에 개발되고 있는 효소공학, 시스템생물공학 등에 기반한 바이오기술들이 그 역할을 수행할 것 으로 기대된다[26].
본 기고문에서 바이오나일론을 예로 들어 살펴 보았듯이 대사공학적으로 개량된 우수한 미생물 호스트 균주와 활성이 우수한 바이오효소촉매들 이 기존의 화학산업과 성공적으로 융합된 바이오 화학산업의 발전에 핵심적인 역할을 할 것은 자명 해보이나 초기투자비용이 매우 높은 만큼 정부의 적극적인 투자가 필요하며, 기존의 바이오산업의 한계를 극복하면서 화학산업과 융합될 수 있는 산 업바이오화학기술의 개발을 위해 산⋅학⋅연의 여러 연구자들이 힘을 모아야 할 것이다.
5. 감사의 글
This work was supported by Industrial Strategic Technology Development Program (10047910, Production of biobased cadaverine and polymer- ization of Bio-polyamide 510) funded by the Ministry of Trade, Industry & Energy (MOTEI, Korea) and the Technology Development Program to Solve Climate Changes (Systems Metabolic Engineering for Biorefineries) from the Ministry of Science, ICT and Future Planning (MSIP) through the National Research Foundation (NRF) of Korea (NRF-2012-C1AAA001-2012M1A2A20 26556).
참 고 문 헌
1. The future of industrial biorefineries, World Economic Forum report (2010).
2. E. Roberts and S. Frankel, Glutamic acid decarboxylase in brain, J. Biol. Chem., 188, 789-795 (1951).
3. 김민홍, 효소를 이용한 고순도 감마 아미노 부 틸산의 제조방법 국내등록특허 10-0857215 (2008).
4. T. H. Dinh, N. A. T. Ho, T. J. Kang, K. A.
McDonald, and K. Won, Salt-free production of γ-aminobutyric acid from glutamate using glutamate decarboxylase separated from Escherichia coli, J. Chem. Tech. Biotechnol., DOI: 10.1002/jctb.4251 (2013).
5. H. Li, T. Qiu, G. Huang, and Y. Cao, Production of gamma-aminobutyric acid by Lactobacillus brevis NCL912 using fed-batch fermentation, Microb. Cell. Fact., 9, 85-92 (2010).
6. T. D. Le Vo, T. W. Kim, and S. H. Hong, Effects of glutamate decarboxylase and gamma-aminobutyric acid (GABA) trans- porter on the bioconversion of GABA in engineered Escherichia coli, Bioprocess
Biosyst. Eng., 35, 645-650 (2012).7. T. D. Le Vo, J. S. Ko, S. H. Lee, S. J. Park, and S. H. Hong, Overexpression of Neurospora crassa OR74A glutamate decar- boxylase in Escherichia coli for efficient GABA production, Biotechnol. Bioprocess
Eng., 18, 1062-1066 (2013).8. S. J. Park, E. Y. Kim, W. Noh, H. M. Park,
Y. H. Oh, and S. H. Lee, Metabolic
engineering of Escherichia coli for the
production of 5-aminovalerate and glutarate as
C5 platform chemicals, Metab. Eng., 16,
42-47 (2013).
coli ldcC gene encodes another lysine decarboxylase, probably a constitutive enzy- me, Genes Genet. Syst., 72, 167-172 (1997).
12. N. Kiyohiko, E. Shuichi, M. Yukiko, T.
Kazuhiko, and H. Yoshinori, Method for producing cadaverine dicarboxylase, US patent, US 0003497 A1 (2005).
13. Z. G. Qian, X. X. Xia, and S. Y. Lee, Metabolic engineering of Escherichia coli for the production of putrescine: a four carbon diamine, Biotechnol. Bioeng., 104, 651-662 (2009).
14. Z. G. Qian, X. X. Xia, and S. Y. Lee, Metabolic engineering of Escherichia coli for the production of cadaverine: a five carbon diamine, Biotechnol. Bioeng., 108, 93-103 (2011).
15. M. Volkert, O. Zelder, B. Ernst, and W. K.
Jeong, Method forfermentatively producing 1,5-diaminopentane, US patent, US 0292429 A1 (2010).
16. M. Ikeda, J. Ohnishi, M. Hayashi, and S.
Mitsuhashi, A genome-based approach to create a minimally mutated Corynebacterium glutamicum strain for efficient L-lysine production, J. Ind. Microbiol. Biotechnol., 33, 610-615 (2006).
17. J. Becker, O. Zelder, S. Hafner, H. Schroder, and C. Wittmann, From zero to hero—Design-
Yamada, Metabolic engineering of Coryne- bacterium glutamicum for cadaverine fermen- tation. Biosci. Biotechnol. Biochem., 71, 2130- 2135 (2007).
20. T. Tateno, Y. Okada, T. Tsuchidate, T.
Tanaka, H. Fukuda, and A. Kondo, Direct production of cadaverine from soluble starch using Corynebacterium glutamicum coexpre- ssing alpha-amylase andlysine decarboxylase,
Appl. Microbiol. Biotechnol., 82, 115-121(2008).
21. S. Verseck, H. Hager, A. Karau, L. Eggeling, and H. Sahm, Verfahrenzur fermentativen Herstellung von Cadaverin. DE 1020070050- 72A1 (2008).
22. S. Kind, W. K. Jeong, H. Schoroder, C.
Zelder, and C. Wittmann, Identification and elimination of the competing N-acetyl- diaminopentane pathway for improved production of diaminopentane by Coryne- bacterium glutamicum, Appl. Environ. Micro-
biol., 76, 5175-5180 (2010).23. S. Kind, W. K. Jeong, H. Schoroder, and C.
Wittmann, Systems-wide metabolic pathway engineering in Corynebacterium glutamicum for bio-based production of diaminopentane,
Metab. Eng., 12, 341-351 (2010).24. N. Buschke, H. Schroder, and C. Wittmann,
Metabolic engineering of Corynebacterium
오 영 훈
2005∼2009 서강대학교 화공생명공학 학사 2009∼2011 서강대학교 화공생명공학 석사 2011∼현재 한국화학연구원 바이오화학연
구센터 연구원
이 승 환
1993∼1998 KAIST 생물공학과 학사 1998∼2000 KAIST 화학공학과 석사 2000∼2004 KAIST 생명화학공학과 박사 2004∼2006 KAIST 응용과학연구소 Post
Doc
2006∼2013 한국화학연구원 바이오화학연 구센터 선임연구원
2013∼현재 한국화학연구원 바이오화학연 구센터 책임연구원
박 시 재
1993∼1997 서울대학교 화학공학과 학사 1997∼1999 KAIST 화학공학과 석사 1999∼2003 KAIST 생명화학공학과 박사 2003∼2009 LG화학기술연구원 CRD연구소
차장
2009∼2012 한국화학연구원 바이오화학연 구센터 선임연구원 2012∼현재 명지대학교 환경에너지공학과
조교수
glutamicum for production of 1,5-diamino- pentane from hemicellulose, Biotechnol. J. 6, 306-317 (2011).
25. S. J. Park, E. Y. Kim, W. Noh, Y. H. Oh, H. Y. Kim, and B. K. Song, Synthesis of nylon4 from gamma-aminobutyrate (GABA) produced by recombinant Escherichia coli,
Bioprocess Biosyst. Eng., 36, 885-892 (2013).

![Figure 4. 바이오기술을 활용한 cadaverine의 생산. 0.5 g/L의 5AVA가 회분식 배양과 유가식 배양을 통해 글루코스로부터 생산될 수 있었다[8]](https://thumb-ap.123doks.com/thumbv2/123dokinfo/4963458.300458/7.825.200.623.117.449/바이오기술을-활용한-cadaverine의-회분식-배양과-유가식-배양을-글루코스로부터.webp)

![[기획특집: 바이오 촉매] 바이오에탄올로부터 부타디엔 제조를 위한 촉매전환기술 개발](data:image/gif;base64,R0lGODlhAQABAIAAAP///wAAACH5BAEAAAAALAAAAAABAAEAAAICRAEAOw==)