Article
http://dx.doi.org/10.4217/OPR.2011.33.4.447
지구시스템모형을 이용한 황동중국해 이산화탄소분압 분포 특성 평가
박영규
1·최상화
2·김철호
11한국해양연구원 기후·연안재해연구부
2한국해양연구원 해양자료정보사업단 (426-744) 경기도 안산시 상록구 해안로 787
Assesment of pCO
2in the Yellow and East China Sea Using an Earth System Model
Young-Gyu Park
1, Sang-Hwa Choi
2, and Cheol-Ho Kim
11Climate Change & Coastal Disaster Research Department, KORDI
2Ocean Data and Information Unit, KORDI Ansan P.O. Box 29, Seoul 425-600, Korea
Abstract : Using results from an earth system model, the distribution of partial pressure of CO2 (pCO2) in surface seawater over the East China Sea is investigated. In this area pCO2 shows minimum along the edge of the continental break along the path of the Taiwan-Tsushima Current System. Apparently modelled chlorophyll is also great along the current but the maximum of the chlorophyll and the minimum of pCO2 do not coincide suggesting that the primary production is not the main cause of the pCO2 minimum. As we move toward the Yellow Sea from the Kuroshio area the temperature decreases so that the pCO2 becomes smaller. If we move further toward the Yellow Sea beyond the Taiwan-Tsushima Current System, alkalinity starts to drop substantially to intensify pCO2 while overcoming the effect of decreasing temperature and salinity. Thus pCO2 minimum occurs along the Taiwan-Tsushima Current System. Of course, the primary production lower pCO2 during spring when it is high but the effect is local. Near the Yangtze river mouth and northeastern corner of the Yellow Sea the fresh water input is large enough and dissolved inorganic carbon (DIC) becomes low enough so that pCO2 becomes lower again.
Key words : East China Sea, biogeochemical modeling, pCO2
1. 서 론
동중국해는 서쪽으로 대만해협을 통해서 남중국해와, 동쪽으로는 남해를 통하여 동해와, 남쪽으로는 쿠로시오 를 통하여 북태평양과, 북쪽으로는 황해와 연결되어있어 우리나라 해양환경과 밀접하게 연관되어 있다. 따라서 우 리나라 주변 해양환경과 그 변화를 파악함에 있어 동중국 해는 반드시 고려되어야 한다.
동중국해는 전체의 약 70%(0.9 × 106km2)가 수심 200 m 이하의 대륙붕인 세계에서 세 번째로 큰 지역해로 다양 한 생지화학 현상이 나타나는 곳이다(Wong et al. 2000;
Gong et al. 2003). 여러 연구결과 동중국해는 0.013-0.030 PgC yr−1 정도의 이산화탄소를 흡수하는 것으로 추정되며 대륙붕 펌프가 작용하고 있을 것으로 추정된다(Tsunogai et al. 1997, 1999; Peng et al. 1999; Wang et al. 2000;
Shim et al. 2007; Chou et al. 2009; Zhai and Dai 2009).
하지만 이런 결론은 제한된 지역의 연구 결과를 다른 지 역이나 다른 계절로 외삽하여 얻은 것이어서 동중국해의
*Corresponding author. E-mail : [email protected]
여 기후변화에 의한 동중국해의 표층 이산화탄소 분압 (pCO2)을 평가하였다. 이 지구시스템모형은 NOAA/GFDL 에서 IPCC 4차 보고서에 사용한 CM2.1 모형에 TOPAZ (Tracers for Ocean Phytoplankton with Allometric Zoo- plankton)라는 해양생태계 모델이 추가된 것이다(Dunne et al. 2010; Sarmiento et al. 2010). CM2.1 모형은 해양·대 기·해빙·육상모형이 접합된 전지구 기후모형인데, 이 모 형은 한반도 주변 해역을 약 1도 정도의 해상도로 표현한 다. 물론 1도 정도의 해상도로는 한반도 주변해역을 자세 하게 모사할 수는 없다. 하지만 기후변화에 의한 반응을 평가하는데 사용하는 고해상도 지역해 모델에서도 저해상 도 기후모델의 결과를 경계조건으로 사용하기 때문에, 저 해상도 기후모델 상태로도 동중국해 해역에 대한 평균적 인 해황 평가가 가능하다. 이런 저해상도 모델 결과를 분 석함으로써 고해상도 모델에 필요한 조건이 무엇인지도 파악할 수 있다. 여기에서는 2100년까지 대기 이산화탄소 농도가 현재의 두 배로 증가하는 A1B 시나리오 결과 중 2001년부터 2020년 사이 결과를 월별로 평균하여 동중국 해 표층의 이산화탄소 분압에 대하여 설명하였다.
2. 모 델
NOAA/GFDL CM2.1의 기본적인 구성과 특성은 Del- worth et al. (2006)과 Gnanadesikan et al. (2006)에 자세 히 기술되어 있다. 이 기후 모형과 결합된 해양 식물플랑 크톤과 관련 동물플랑크톤을 추적자로 사용하는 생지화학 모형(Tracers for Ocean Phytoplankton with Allometric Zooplankton, TOPAZ)은 적은 수의 관계식으로 해양생태 계를 정확하게 모사하기 위해 고안되었다. 모든 주요 영양 염이 포함되었으며(N, P, Si와 Fe), 분해되기 쉬운 유기물
영양염과 철에 대한 제한은 식물플랑크톤 생리학(Sunda and Huntsman 1997)에 기초해 모수화하였다. 누락된 항 목은 동물플랑크톤의 섭식과 입자형태의 유기물 손실이다.
쇄설물의 재광물화와 용존유기물의 순환은 포함되어있다 (Dunne et al. 2005). 모형에서는 용존산소와 용존무기탄 소(DIC), 알칼리도, 식물플랑크톤 생리, 엽록소량:탄소 비 율 등이 추적자로 활용된다. 모형의 생태계는 세 종류의 식물플랑크톤으로 구성된다. “작은(Small)” 식물플랑크톤 은 우연히 침강하는 침전물에 끼이는 경우를 제외하고는 생산대에서 벗어나지 않으며, 미생물 순환계와 밀접하게 관련되어있는 남세균(cynobacteria, Prochlorococcus와 Synechococcus)과 미소진핵생물(picoeukaryotes) 등을 나 타낸다. “큰(Large)” 식물플랑크톤은 좀 더 빠르게 침강하 는 종들로 규산염이나 탄산염 골격으로 구성되어 있어 침 강하기 쉬운 종들로 구성되어 있으며 규조류와 기타 진핵 생물을 포함한다. 이러한 골격에 의한 입자성 유기탄소의 보호로 인해 유기탄소가 수주(water column) 내에서 더 깊이 침강할 수 있게 된다(Klaas and Archer 2002). “질소 고정생물(diazotrophs)”은 질소를 용존된 형태로 흡수하기 보다는 직접 고정하는 종류를 나타낸다. 모든 식물플랑크 톤은 광량과 다수의 영양염에 의해 동시에 제한을 받지만, 식물플랑크톤 종류에 따라 영양염 생리와 성장률을 결정 하는 인자가 각기 다르게 적용된다. 생지화학모형은 여러 화학종(C, N, P, Si, Ca/Alk과 Fe)과 작은 식물플랑크톤, 큰 식물플랑크톤, 질소고정생물, 유기쇄설물의 재광물화 과정과 용존 유기물의 순환 등의 과정을 포함하여 구성되 었다. 습식과 건식 침전물 유입량은 Ginoux et al.(2001)의 월별자료에 기초했으며, 다양한 용해도 인자로부터 용해 성 철을 구했다(Fan et al. 2006). TOPAZ에는 생물섭식과 재광물화, 입자침강, 소기(scavenging)와 흡착/용해를 포함
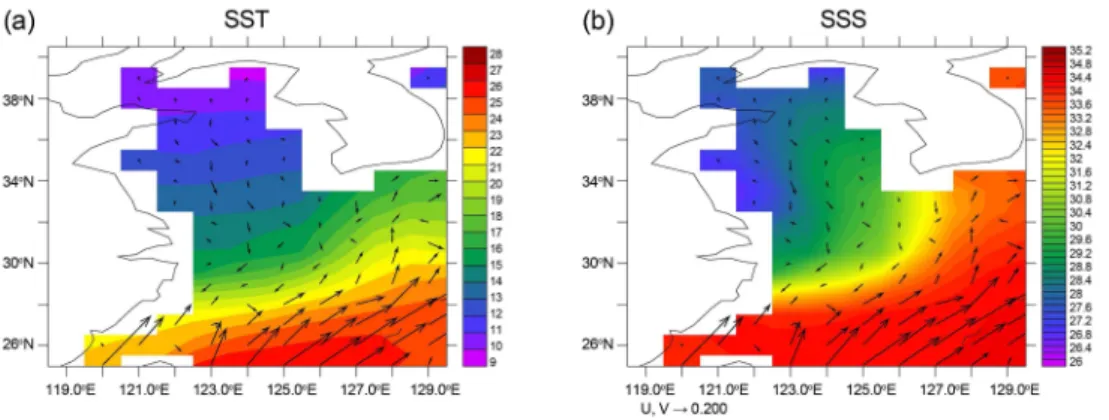
Fig. 1. (a) Surface temperature; (b) surface salinity with surface flows averaged between 2001 and 2020
한 단순화된 해양 철 순환이 포함되어 있다.
3. 결과 및 고찰
Fig. 1에 모델 결과 중 표층수온과 염분을 21세기 최초 20년 즉 2001-2020년 사이 평균값으로 나타내었다. 그림 에 화살표로 표층해류도 나타내었다. 이 모델의 해상도가 1도여서 쿠로시오나 대마난류 등을 관측에서 나타나는 것 처럼 자세히 나타낼 수는 없으나 동중국해 대륙사면을 따 르는 쿠로시오와 대만해협을 통과하는 대만난류, 동중국 해에서 대한해협을 통해 동해로 유입되는 대마난류가 재 현되었다. 수온은 태평양쪽에서 동중국해를 지나 황해쪽 으로 감소한다. 염분도 기본적으로 유사한 분포특성을 보 이는데, 양자강 입구와 황해 북동부에서는 양자강과 두만 강에 의한 저염수가 두드러지게 나타난다.
Fig. 2에는 표층의 이산화탄소 분압과 알칼리도, DIC, 해양-대기 이산화탄소 교환량(양의 값은 해양이 대기에서 이산화탄소를 흡수하는 경우, 음의 값은 반대의 경우를 의 미함)을 나타내었다. 이산화탄소 분압은 북서태평양에서 동중국해 방향으로 감소하다가 황해로 접어들면 다시 증 가하여 발해만 부근에서 최대값이 나타난다. 양자강 입구 와 황해 북동부에서도 이산화탄소 분압이 낮게 나타난다.
알칼리도는 염분과 거의 유사한 분포를 보여 쿠로시오 해 역에서 동중국해를 거쳐 황해 쪽으로 가면서 감소한다. 염 분과 마찬가지로 양자강 입구와 두만강 하구역에서 최소
값이 나타난다. DIC도 알칼리도와 마찬가지로 염분과 유 사한 분포를 보여 양자강과 두만강을 통한 담수유입이 동 중국해와 황해에서 알칼리도와 DIC의 분포를 지배하는 가장 큰 요인임을 알 수 있다.
이산화탄소 분압은 수온, 염분 등 해수의 물리적 특성 과 DIC, 알칼리도 등 화학적 특성에 의해 복합적으로 결 정된다. 북서태평양에서 동중국해로 접어들어오면서 수온 이 감소하여 이산화탄소 분압이 낮아진다. 물론 DIC의 감 소도 이에 기여하지만 알칼리도 감소에 의해 효과가 서로 상쇄되므로 수온의 변화가 더 크게 작용한다. 염분이나 수 온이 낮아지면 이산화탄소 분압이 낮아지는데 황해로 접 어들면 수온, 염분, DIC가 모두 감소하는데도 이산화탄소 분압은 증가한다. 이는 알칼리도 감소의 영향이 더 크게 작용했기 때문이다. 양자강 입구와 황해 북동부는 DIC 감 소의 영향이 알칼리도의 영향을 뛰어넘어 이산화탄소 분 압이 낮아진다. 즉 동중국해의 이산화탄소 분압은 대양의 특성을 띄는 대륙사면 부분에서는 수온에 의해, 황해에서 강 하구를 제외한다면 알칼리도에 의해, 강 하구에서는 DIC에 의해 주로 결정된다. Shim et al. (2007)은 동중국 해 북부해역 중 쿠로시오의 영향을 많이 받는 동경 125~
126도 동쪽 해역은 표층 이산화탄소 분압이 주로 수온에 의해 결정되는 것으로 보고했다. 또한, 상대적으로 수심이 얕은 동경 125~126도 서쪽 해역은 계절에 따른 담수유입 혹은 수층수직혼합 등에 의한 DIC 변화로 인해 표층 이산 화탄소 분압이 결정되는 것으로 보고했다. 연구해역 내에 Fig. 2. (a) Surface pCO2; (b) alkalinity; (c) DIC; (d) air-sea CO2 exchange averaged between 2001 and 2020
이산화탄소가 흡수되는 것으로 나타난다. 최대값은 대륙 붕 끝단을 따르는 대만-대마난류계를 따라 나타난다.
동중국해 이산화탄소 분압에 영향을 주는 또 하나의 요
나타내었다. 모델에서 모사된 엽록소량은 관측된 값과 유 사한 범위에 있다(Son et al. 2005; Kim et al. 2009). 대륙 붕 경계를 따라 황해 쪽으로 대만해협부터 대한해협까지 높은 값을 나타낸다. Fig. 1에서 볼 수 있듯이 이 해역에서 황해 내부에서 대만해협 쪽으로 남서진하는 흐름과 북동 진하는 대만-대마난류계가 만난다. 이 두 흐름은 서로 방 향이 반대이므로 두 물이 만나는 곳에서 강한 혼합이 생 겨 저층의 영양염이 표층으로 공급되면서 생물활동이 높 아진다.
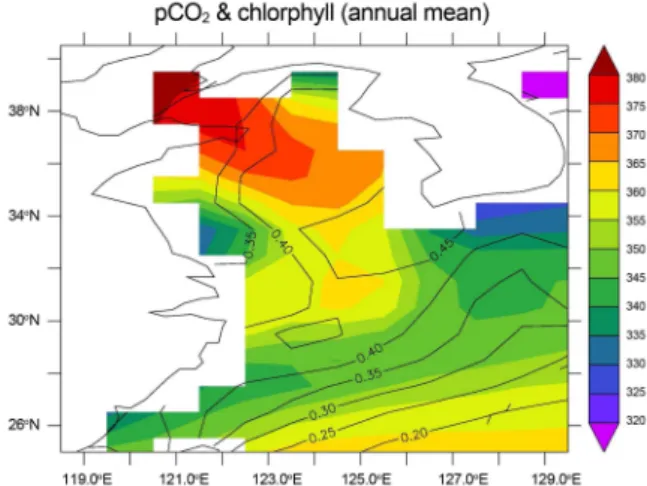
Fig. 3에 이산화탄소 분압도 함께 나타내었는데, 이산화 탄소 분압이 최소인 해역과 엽록소가 최대인 해역이 대륙 붕 경계를 따라 나타나나, 이산화탄소 분압이 최소인 해역 이 엽록소량이 최대인 해역보다 남쪽에 나타나 두 분포가 일치하지 않음을 알 수 있다. 양자강 하구역과 황해 북동 부에서는 생물활동이 낮으나 이산화탄소 분압이 낮게 나 타난다. 즉 연평균 분포에서는 이산화탄소 분압이 생물활 동이 아니라 해수의 물리화학적 특성에 의해 결정됨을 알 수 있다. 이런 경향은 Shim et al. (2007)의 경우처럼 표층 30 m까지만, 혹은 Zhai and Dai (2009)에서처럼 표층 40 m층까지만 비교하더라도 같은 결과가 나타난다. 남해 Fig. 3. Comparison of surface pCO2 (shading), and chloro-
phyll averaged between surface and 70 m (con- tours). Both values are obtained from average between 2001 and 2020
Fig. 4. Comparison of surface pCO2 (shading), and (a) chlorophyll averaged between surface and 70 m; (b) SST; (c) SSS (contour) during spring (March, April and May) between 2001 and 2020
안에서는 엽록소는 높게, 이산화탄소 분압은 낮게 나타나 생물활동이 이 해역의 이산화탄소 분압을 낮추는데 기여 했음을 알 수 있다. 하지만 이 모델이 해상도가 낮아 연안 에서 나타나는 현상을 잘 재현할 수 없기 때문에 이 현상 의 신뢰도는 높지 않다.
Zhai and Dai (2009)는 양자강 외해역에서 4월에서 10 월 사이 생물활동으로 이산화탄소 분압이 감소한다고 하 였다. 모델의 해상도가 낮아 양자강 하구역에서 나타나는 해양현상을 재현할 수 없으나 생물활동이 높아지는 춘계 에 생물활동과 이산화탄소 분압간 관계를 파악하기 위하 여 Fig. 4에서 봄철에 국한하여 이산화탄소 분압과 엽록소 량을 비교하였다. 대륙사면 남쪽과 쿠로시오 해역에서는 이산화탄소 분압과 SST가 비슷한 모양으로 분포하기 때 문에 연평균에서와 마찬가지로 이산화탄소 분압 분포를 결정함에 있어 SST가 가장 중요한 요인임을 알 수 있다.
엽록소 농도가 최대인 남해연안에서는 이산화탄소 분압이 최소가 되고 엽록소와 이산화탄소 분압이 유사하게 분포 하므로 생물활동이 이산화탄소 분압을 낮춤을 알 수 있다.
이런 경향은 겨울철 이외 모든 계절에 나타난다. 양자강 하구 남쪽(북위 28도, 동경 123도 부근)에서도 발견되는 이산화탄소 분압이 낮은 해역에서 엽록소 농도가 높게 나 타난다. 따라서 이 해역에서도 생물활동이 이산화탄소 분 압 분포에 영향을 준다는 것을 알 수 있다. 연평균분포에 서와 마찬가지로 황해 내부로 들어가면 알칼리도가 감소
하여 이산화탄소 분압이 높아진다. 즉 동중국해 대륙사면 부근에서는 온도과 염분 감소에 의한 효과, 생물활동에 의 한 효과, 알칼리도에 의한 효과가 섞여서 나타나는데, 염 분과 알칼리도가 가장 우세한 요인이고 생물활동은 국지 적으로 작용한다. 생물활동의 영향은 생물활동이 가장 크 게 나타나는 봄철 이외에는 큰 영향을 보이지 않는다.
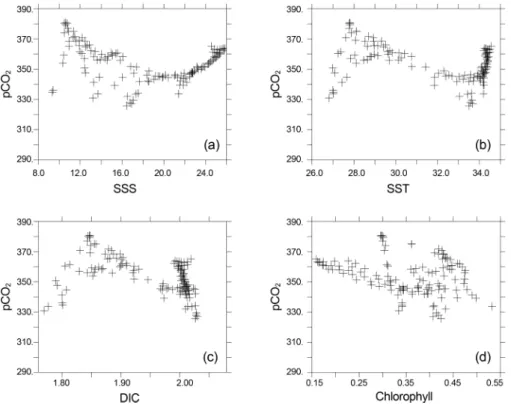
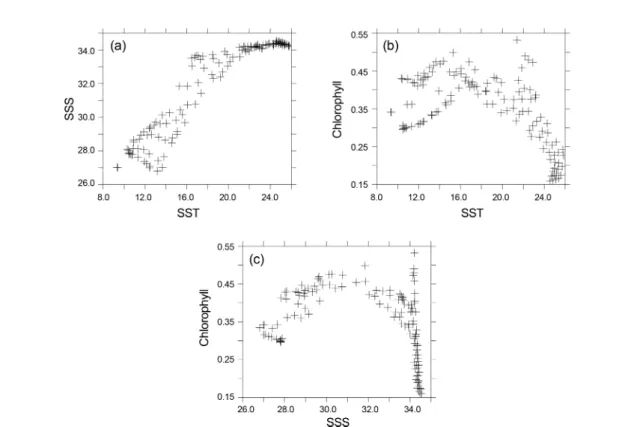
표층 이산화탄소 분압을 조절하는 요인을 좀 더 정량적 으로 파악하기 위하여 Fig. 5에 표층 이산화탄소 분압과 표층 수온, 표층 염분, 표층 DIC, 표층부터 70 m까지 평균 된 엽록소량을 비교하였다. Fig. 6에서는 표층 수온과 표 층 염분, 표층부터 70 m까지 평균된 엽록소량을 비교하였 다. 비교에 사용된 영역은 Fig. 1에 표시된 영역 중 동해 부분(원산만)을 제외한 부분이다. 표층수온이 18oC 이상 이면 이산화탄소 분압과 표층 수온은 매우 뚜렷한 정의 상관관계를 갖는다. 이런 경향이 나타나는 해역은 대마난 류와 대만난류, 대마난류의 영향을 받는 동중국해 외부해 역이다. 수온이 18oC 이하로 낮아지면 이산화탄소 분압이 전반적으로 올라가지만 열역학적으로 수온은 이산화탄소 분압과 정의 상관성을 가지므로 수온이 이산화탄소 분압 을 지배하는 요인이 아님을 알 수 있다. 둘 사이 상관관계 도 밀접하지 않다.
염분이 29에서 33사이일 때 염분이 감소함에 따라 이산 화탄소 분압이 증가한다. 이런 관계가 나타나는 해역은 황 해와 동중국해 내부로 담수의 영향을 많이 받는 곳이다.
Fig. 5. Comparison of surface pCO2 versus (a) SST; (b) SSS; (c) surface DIC; (d) chlorophyll averaged between surface and 70 m
이 범위에서는 염분과 DIC가 감소하지만 알칼리도 감소 가 더 커 염분이 감소할수록 이산화탄소 분압이 증가한다.
염분이 34보다 높은 동중국해 외해역과 쿠로시오 해역에 서는 염분이 증가함에 따라 이산화탄소 분압이 연직방향 Fig. 6. Comparison of SST versus (a) SSS; (b) chlorophyll
averaged between surface and 70 m; (c) SSS versus chlorophyll averaged between surface and 70 m
Fig. 7. pCO2 (shading) and SST (contour) for (a) spring (March, April and May); (b) summer (June, July and August);
(c) fall (September, October and November); (d) winter (December, January and February)
으로 매우 급격하게 증가한다(Fig. 6b). 앞서 설명한 것과 같이 이 해역에서는 이산화탄소 분압이 수온변화에 의해 결정되는데, 대양의 특성을 띄는 이 해역에서는 수온변화 에 비해 염분변화가 매우 작기 때문에 염분과 이산화탄소 분압을 비교하면 염분에 대해 이산화탄소 분압이 매우 급 격하게 변한다.
염분이 28 이하로 떨어지면 DIC 감소 효과가 알칼리도 감소 효과를 상쇄시켜 염분 감소에 따라 이산화탄소 분압 이 감소한다. Figs. 1과 2의 비교에서 알 수 있듯이 DIC와 알칼리도는 염분과 매우 높은 상관관계를 보인다. 특히 이 산화탄소 분압과 DIC와의 관계는 이산화탄소 분압과 염 분의 관계와 매우 유사하게 나타난다. 그러나, 알칼리도와 DIC는 해수 내 이산화탄소에 서로 상반된 영향을 미치므 로, 알칼리도와 DIC의 상대비 차이에 의해 이산화탄소 분 압은 변화를 나타내게 된다.
엽록소량과 이산화탄소 분압은 엽록소량이 약 0.3 mg m−3보다 작을 때는 엽록소량이 증가함에 따라 이산화탄소 분압이 감소한다. 이런 밀접한 상관관계가 나타나는 해역 은 생물활동이 큰 동중국해나 황해 내부가 아니라, Fig.
6b에 나타낸 것과 같이 수온이 20oC 이상인 외해역이다.
즉 수온에 의해 엽록소량과 이산화탄소 분압이 동시에 지 배된다고 보는 것이 타당하다. 엽록소량이 0.3 mg m−3보 다 큰 경우 엽록소량과 이산화탄소 분압간에는 특별한 상 관관계가 보이지 않는다. 국지적으로 높은 생산량이 이산
화탄소 분압을 낮출 수는 있지만 전반적인 동중국해 이산 화탄소 분압 분포를 지배하는 주요 원인이 아니다.
이산화탄소 분압의 계절변화와 SST는 Fig. 7에 나타내 었다. Fig. 8에는 이산화탄소 분압과 SSS를 계절별로 나 타내었다. 전반적인 경향, 즉 이산화탄소 분압이 쿠로시오 해역에서 동중국해 쪽으로 감소하다가 황해 내부로 가면 서 증가하는 분포는 모든 계절에 유사하게 나타난다. 겨울 철에 수온이 낮아 이산화탄소 분압이 가장 낮게 나타난다.
이는 겨울철에 이산화탄소 분압이 다른 계절에 비해 매우 낮게 나타남을 보고한 Kim et al. (submitted) 등과도 유사 한 결과이다. 여름철과 가을철에 양자강 북쪽과 황해 북동 부에서 이산화탄소 분압이 낮게 나타나는데 이는 연평균 분포에서 설명했듯이 강을 통한 담수유입에 의한 것이다.
4. 요약 및 결론
동중국해는 쿠로시오의 영향을 받아 대양의 특성을 띠 는 해수와 연안수, 담수 등이 만나 복잡한 현상이 나타나 는 곳이다. 이 해역은 대륙붕 펌프가 작용하는 곳으로도 추정되어 지구의 탄소순환에 중요한 역할을 하는 곳으로 추정되어 이 해역의 해양-대기 이산화탄소 교환에 관한 여러 연구가 진행되어 왔다. 하지만 연구가 일부 해역에만 국한되어 있어, 성격이 복잡한 이 해역의 특성을 모두 포 함하고 있지는 못하고 있다. 여기에서는 지구시스템모델 Fig. 8. pCO2 (shading) and SSS (contour) for (a) spring (March, April and May); (b) summer (June, July and August);
(c) fall (September, October and November); (d) winter (December, January and February)
al. 2007). 수직혼합에 의해 저층의 DIC가 높은 해수가 표 층으로 올라오면서 이산화탄소 분압이 높아진다. 모델에 도 수직혼합이 포함되어 있어 봄철과 가을철에 성층이 약 하게 나타난다. 하지만 실제 해양에서 나타나는 해저퇴적 층에서의 유기물 분해가 모델에서 제대로 모사되지 않는 다. 즉, 모델이 동중국해에서 관측되는 수직혼합에 이산화 탄소 분압의 변화를 제대로 나타내지는 못하였다. 이 결과 에서 퇴적층과 해수의 상호작용도 천해의 생지화학과정 모델에서 중요한 과정이라 추론할 수 있다.
쿠로시오 해역에서 동중국해로 들어오면 수온이 감소하 여 이산화탄소 분압이 낮아지는데 대만-대마난류계를 지 나 황해 쪽으로 접어들면 알칼리도가 감소하여 이산화탄 소 분압이 다시 높아진다. 따라서 대만-대마난류계를 따라 이산화탄소 분압이 최소가 된다. 양자강 하구역과 황해 북 동부의 두만강 근처에서는 담수유입에 의한 저염화와 낮 은 DIC로 인해 이산화탄소 분압이 다시 낮아진다. 대만- 대마난류계의 북쪽 경계에서 황해에서 남서진하는 해류와 혼합에 의해 저층의 영양염이 공급되면서 생물활동이 높 아진다. 하지만 엽록소가 최대인 해역과 이산화탄소 분압 이 최소인 해역이 일치하지 않기 때문에 생물활동이 국지 적으로 이산화탄소 분압을 낮출 수는 있지만 전체적인 이 산화탄소 분압 분포를 결정하는 주요 원인은 아니다.
황해를 포함한 동중국해 내부에서 알칼리도와 DIC는 염분과 매우 유사한 분포특성을 지니고 담수유입이 이 해 역의 이산화탄소 분압 분포를 결정함에 있어 중요 요인임 을 알 수 있다. 여기서 분석한 모델은 해상도가 낮아 양자 강이나 두만강을 통해 유입되는 담수와 해양이 일으키는 상호작용을 자세하게 재현해내지 못한다. Shim et al.
(2007)과 Zhai and Dai (2009) 등은 양자강 담수가 생물활 동과도 밀접하게 관련되어 있음을 시사한다. 따라서 동중 국해의 생지화학과정을 구체적으로 재현하려면 강을 통한 담수유입이 고려된 고해상도 모형이 필요하다.
사 사
본 논문을 세심하게 검토해주신 심사위원님들께 깊은 감사를 드립니다. 본 연구는 “지구시스템모듈 원천기술 개발을 위한 해빙 및 해양 생지화학과정 개발(PN65060)”
과 “기후변화가 남해권 해양생태계에 미치는 영향 및 기 능평가기술개발(PM56600)”, “유출 CO2의 거동 모사 및 모니터링 표준화 구축(PMS227D)”의 지원을 받아 수행되 었습니다.
Res 114:C07026. doi:10.1029/2008JC005128
Delworth TL, Broccoli AJ, Rosati A, Stouffer RJ, Balaji V, Beesley JA, Cooke WF, Dixon KW, Dunne J, Dunne KA, Durachta JW, Findell KL, Ginoux P, Gnanadesikan A, Gordon CT, Griffies SM, Gudgel R, Harrison MJ, Held IM, Hemler RS, Horowitz RW, Klein SA, Knutson TR, Kushner PJ, Langenhorst AR, Lee HC, Lin SJ, Lu J, Malyshev SL, Milly PCD, Ramaswamy V, Russell J, Schwarzkopf MD, Shevliakova E, Sirutis JJ, Spelman MJ, Stern WF, Winton M, Wittenberg AT, Wyman B, Zeng F, Zhang R (2006) GFDL’s CM2 global coupled climate models. part I: formulation and simulation characteristics. J Climate 19:643-674. doi:10.
1175/JCLI3629.1
Dunne JP, Armstrong RA, Gnanadesikan A, Sarmiento JL (2005) Empirical and mechanistic models for the particle export ratio. Global Biogeochem Cy 19:GB4026. doi:10.
1029/2004GB002390
Dunne JP, Gnanadesikan A, Sarmiento JL, Slater RD (2010) Technical description of the prototype version (v0) of Tracers Of Phytoplankton with Allometric Zooplank- ton (TOPAZ) ocean biogeochemical model as used in the Princeton IFMIP model. Biogeosci Disc 6:10381- 10446
Fan SM, Moxim W, Levy H (2006) Aeolian input of bioavailable iron to the ocean. Geophys Res Lett 33:
L07602. doi:10.1029/2005GL024852
Geider RJ, MacIntyre HL, Kana TM (1997) A dynamic model of phytoplankton growth and acclimation:
responses of the balanced growth rate and chlorophyll a:
carbon ratio to light, nutrient limitation and temperature.
Mar Ecol Prog Ser 148:187-200
Ginoux P, Chin M, Tegen I, Prospero JM, Holben B, Dubovik O, Lin SJ (2001) Sources and distributions of dust aerosols simulated with the GOCART model. J Geophys Res 106(17):20255-20273
Gnanadesikan A, Dixon KW, Griffies SM, Balaji V, Barreiro M, Beesley JA, Cooke WF, Delworth TL, Gerdes R, Harrison MJ, Held IM, Hurlin WJ, Lee HC, Liang Z, Nong G, Pacanowsky RC, Rosati A, Russell J, Samuels BL, Song Q, Spelman MJ, Stouffer RJ, Sweeney CO, Vecchi G, Winton M, Wittenberg AT, Zeng F, Zhang R, Dunne JP (2006) GFDL’s CM2 Global Coupled Climate Models. Part II: the baseline ocean simulation. J Climate
19:675-697. doi: 10.1175/JC LI3630.1
Gong GC, Wen YH, Wang BW, Liu GJ (2003) Seasonal variation of chlorophyll a concentration, primary production and environmental conditions in the subtrop- ical East China Sea. Deep-Sea Res II 50:1219-1236 Kim D, Choi SH, Kim KH, Shim JH, Yoo S, Kim CH
(2009) Spatial and temporal variations in nutrient and chlorophyll-a concentrations in the northern East China Sea surrounding Cheju Island. Cont Shelf Res 29:1426- 1436
Kim D, Choi SH, Shim JH, Kim KH, Kim CH (2011) Seasonal variability of sea-air CO2 fluxes in the northern East China Sea. Cont Shelf Res (submitted)
Klaas C, Archer D (2002) Association of sinking organic matter with various types of mineral ballast in the deep sea: implications for the rain ratio. Global Biogeochem Cy 16(4):1116. doi:10.1029/2001GB001765
Peng TH, Hung JJ, Wanninkhof R, Millero F (1999) Carbon budget in the East China Sea in spring. Tellus B 51(2):531-540
Sarmiento JL, Slater RD, Junne J, Gnanadesikan A, Hiscock MR (2010) Efficiency of small scale carbon mitigation by patch iron fertilization. Biogeosciences 7:3593-3624 Shim JH, Kim D, Kang YC, Lee JH, Jang ST, Kim CH
(2007) Seasonal variations in pCO2 and its control- ling factors in surface seawater of the northern East China Sea. Cont Shelf Res 27:2623-2636
Son S, Campbell J, Dowell M, Yoo S, Noh J (2005) Primaty production in the Yellow Sea determined by ocean color remote sensing. Mar Ecol Prog Ser 303:91- 103
Sunda WG, Huntsman SA (1997) Interrelated influence of iron, light, and cell size on marine phytoplankton growth. Nature 390:389-392
Tsunogai S, Watanabe S, Nakamura J, Ono T, Sato T (1997) A preliminary study of carbon system in the East China Sea. J Oceanogr 53:9-17
Tsunogai S, Watanabe S, Sato T (1999) Is there a
“continental shelf pump” for the absorption of atmos- pheric CO2? Tellus B 51(3):701-712
Wang SL, Chen CTA, Hong GH, Chung CS (2000) Carbon oxide and related parameters in the East China Sea. Cont Shelf Res 20:525-544
Wong GTF, Chao SY, Li YH, Shiah FK (2000) The Kuroshio edge exchange processes (KEEP) study: an introduction to hypotheses and highlight. Cont Shelf Res 20:335-347
Zhai W, Dai M (2009) On the seasonal variation of sir-sea CO2 fluxes in the outer Changjiang (Yangtze River) Estuary, East China Sea. Mar Chem 117:2-10
Received Oct. 28, 2011 Revised Nov. 14, 2011 Accepted Dec. 2, 2011