고추냉이 추출물에 노출된 Bacillus cereus MK-11의 생리 및 세포반응
이현호, 석지원, 오계헌*
Physiological and Cellular Responses of Bacillus cereus MK-11 Exposed to Wasabi Extracts
Hyun-Ho Lee, Ji-Won Seok, and Kye-Heon Oh*
Received: 4 February 2020 / Revised: 23 June 2020 / Accepted: 24 June 2020
© 2020 The Korean Society for Biotechnology and Bioengineering
Abstract: The purpose of this study was to investigate the physiological and cellular responses of Bacillus cereus MK- 11 exposed to wasabi extracts. Initially, MK-11 isolated from commercial ssam-jang was examined for morphological and physiological characteristics. The BIOLOG system and 16S rRNA sequencing analysis were performed to identify the strain. It was assigned and designated as B. cereus MK-11.
The phylogenetic tree of B. cereus MK-11 was plotted based on 16S rRNA sequence comparisons. Wasabi extracts showed a dose-dependent antibacterial effect on B. cereus MK-11, while MK-11 treated using 2 mg/mL wasabi extracts completely inhibited growth within 24 h of incubation. The amount of alginate produced by the exposure of wasabi extracts decreased with increasing concentrations of the extracts, and increased with exposure time. The stress shock proteins (e.g., DnaK and GroEL), which may contribute to enhancing the cellular resis- tance to the cytotoxic effect of wasabi extracts, decreased in proportion to the concentrations of wasabi extracts used for this study. The SEM analysis showed the presence of major structural change in the envelope, as well as a preponderance of irregular rod forms with wrinkled surfaces in cells treated with sublethal concentrations of wasabi extracts. In conclu- sion, it will be a crucial clue to understanding the cytotoxic mechanisms of Bacillus cereus exposed to wasabi extracts.
Keywords: wasabi extracts, Bacillus cereus MK-11, cytotoxic- ity, cellular responses
1. INTRODUCTION
현대인들의 식생활이 서구화 되어감에 따라 성인병의 발생 이 증가하고 있으며, 이런 흐름으로부터 최근 식품산업에서 도 천연 기능성물질들에 대한 연구가 활발하게 진행되고 있 다 [1]. 그 중에서도 고추냉이는 생선 및 어패류를 생식하는 식습관이 보편화되어있는 일본과 우리나라에서 향신료로서 널리 이용되고 있으며, 특히 항균활성에도 관심이 집중되고 있다.
고추냉이 (Euterma japonicum)는 십자화과 (Brassicaceae)에 속하는 다년생 식물로서, 다양한 기능성을 가지는 isothiocyanates 계열의 화합물들이 함유되어 있다고 알려져 있다 [2]. AITC (allyl isothiocyanate)는 고추냉이, 무, 배추, 양배추, 브로콜리, 겨자, 갓 등에 많이 포함되어 있으며, glucosinoates-myro- sinase 반응에서 생성되는 분해산물 중의 한가지로서, 이들 식물을 섭취하였을 경우에 맵고 쏘는 맛을 내는 화합물이다.
AITC 는 혈소판 응집 억제, 항암 및 항산화, 그리고 항균 등의 다양한 효과가 있다고 보고되어 있다 [3,4]. 특히 AITC는 세 균, 효모, 그리고 곰팡이 등에서 강력한 항균력이 있는 것으 로 알려져 있으며, 식중독 원인균인 Aeromonas hydrophilia, Bacillus cereus, Escherichia coli, Pseudomonas fluorescens, Salmonella enterica, Staphylococcus aureus 등에 대한 연구들 이 발표된 바 있다 [5,6]. 최근에 AITC가 포함된 고추냉이 추 출물에 노출된 장내세균인 E. coli와 S. enterica의 사멸율과 세포반응이 보고되었다 [7].
세포외 다당류 (exopolysaccharide)는 다양한 세균에서 분 비되는 고분자량의 중합체 (예, dextran, alginate, xanthan, curdlan, levan 등)로서, 이들의 특성은 의료 및 산업적 측면에 서 많은 관심이 집중되고 있다 [8]. 이들 가운데, alginate는 O- 아세틸화된 β-1-4-linked D-mannuronic acid와 C-5 epimer 인 α-L-guluronic acid로 구성된 exopolysaccharide의 공중합
순천향대학교 자연과학대학 생명시스템학과Department of Life Science and Biotechnology, Soonchunhyang University, Chung-Nam 31538, Korea
Tel: +82-41-530-1353 Fax: +82-41-530-1350 e-mail: [email protected]
Research Paper
체(copolymer)로서, 열악한 주변 환경으로 부터 세균을 보호 하고, 생물막 (biofilm)을 형성하는데 중요한 역할을 하는 것으 로 알려져 있다 [9]. Alginate는 질병과 관련하여 Pseudomonas 속 (genus)에서 많은 연구가 진행되었으며 [9,10], 최근 녹차 폴리페놀인 EGCG에 노출된 Bacillus cereus에서 alginate 생 성량의 변화에 대한 연구가 보고된 바 있다 [11].
Bacillus cereus는 식품매개 병원균으로 장류(醬類), 유제품, 곡류가공품, 즉석조리식품, 어패류 생식품 등에서 오염을 일 으켜, 세균성 식중독의 문제를 초래하는 것으로 알려져 있다 [11-13]. 식중독 관리를 위해 우리나라의 식품의약품안전처 에서는 B. cereus를 식중독균 저위해성으로 구분하였으며, 일 반 식품의 경우 1 g당 1,000 CFU 이하, 이유식의 경우에 1 g 당 100 CFU 이하로 규제하고 있다. 식품제조 및 가공업체는 기준안에 부합되도록 살균처리, 또는 합성 및 천연첨가물 등 을 활용하고 있다. B. cereus의 병원성은 장내에서 조직파괴 와 독소 생산에 기인한다. 이러한 독소 중 용혈을 유도하는 hemolysin BL (HBL), nonhemolytic enterotoxin (NHE), 그리 고 cytotoxin K의 세 가지 용혈성 독소와 구토를 유발하는 cereulide 독소가 알려져 있으며, 감염으로 인해 사망원인으 로 밝혀진 바 있다 [12,14,15].
식품에서 식중독 원인균의 생장을 억제하기 위해서 오랜 기간에 걸쳐 식품보존제 또는 합성 화학첨가제 등이 사용되 어왔다. 그러나 소비자들의 건강에 대한 관심이 커짐에 따라 식중독 원인균의 생장 억제에 효과적이며 다양한 기능성을 가진 천연첨가물의 사용이 주목받고 있다. 이러한 물질로서 고추냉이 또는 그 주요성분인 AITC는 상당히 효과적임에도 불구하고, 그 메커니즘에 대한 연구는 미비한 실정이다.
본 연구에서는 시판 중인 쌈장으로부터 B. cereus를 분리 하여 천연 고추냉이 추출물(wasabi extracts)에 노출된 세균 의 생리 및 세포반응을 조사하였다. 고추냉이 추출물의 다양 한 농도와 시간에 노출되었을 때, B. cereus의 사멸률과 alginate 의 생성량의 변화를 확인하였으며, 고추냉이 추출물 의 노출에 의해 유도되는 스트레스 충격단백질 (stress shock proteins)인 DanK와 GroEL의 변화를 SDS-PAGE 및 Western blot을 통해 조사하였다. 또한 이 세균이 고추냉이 추출물에 노출되었을 때, 세포의 외부형태 변화를 주사전자현미경을 통해 관찰하였다.
2. MATERIALS AND METHODS
2.1. 균주의 확보 및 배양조건
시판 중인 쌈장에서 농화배양 기법을 통하여 B. cereus를 분 리하였다. 시료 5 g을 100 mL의 멸균된 생리식염수가 담긴 플라스크에 넣고 교반하였으며, 20여 분간 정치시킨 후, B.
cereus의 분리를 위해 5% sheep blood가 포함된 blood agar (Difco, Detroit, MI. USA) 고체평판배지에 시료 100 μL를 도 말하여 37
oC에서 48시간동안 배양하였다. 배양된 세균들 가 운데 생장상태가 가장 양호한 집락인 MK-11을 선별하였다.
선별된 세균은 3회에 걸친 도말을 통하여 순수배양으로 최 종 분리하였다. 분리세균은 Luria-Bertani (1 L당 tryptone 10 g, yeast extract 5 g, NaCl 10 g)의 고체 또는 액체배지에 접 종하여 진탕배양기(160 rpm, 37
oC) 에서 배양 및 유지시키며 본 실험에 사용하였다.
2.2. 분리세균의 형태적 관찰 및 생리화학적 특성조사 분리세균인 B. cereus를 LB 고체평판배지에 도말하여 단일 집락을 관찰하고, 그람염색을 한 후 위상차 현미경을 사용하 여 형태학적 특성을 조사하였다. 분리세균에 대한 다양한 생 리 및 생화학적 특성조사로 glucose, lactose, gelatin, starch 등 의 이용여부, indole 생성시험, citrate 이용여부, MR-VP 시험, Klingler iron agar (KIA) 배지에서 H
2S 생성여부, litmus milk 이용여부, catalase와 oxidase 활성여부 등은 기존의 알려진 방법 [16]에 근거하여 실시하였다.
2.3. 분리세균의 동정
분리세균은 GP2 MicroPlate
TM(Biolog, Hayward, CA, USA) 를 통해 다양한 탄소원 이용능력에 근거하여 동정되었다. 분 리세균은 5% sheep blood agar에 도말한 후, 18시간동안 37
oC에서 배양하였다. 배양된 균주를 멸균된 생리식염수에 현탁하여 GP2 MicroPlate에 접종하였다. 이 후 BIOLOG Automated Micro-station Instrument (Biolog, Hayward, CA, USA)를 이용하여 균주를 동정하였다.
2.4. 16S rRNA 염기서열을 통한 계통수 분석
분리세균 MK-11은 16S rRNA 염기서열 분석과 이를 바탕으 로 한 유전학적 계통수(phylogenetic tree)를 작성하기 위하여, 중합효소연쇄반응(PCR)을 통해 16S rRNA 유전자를 증폭하 였다. Genomic DNA는 분리균주로부터 컬럼 방식의 G-spin
TMGenomic DNA Extraction Kit (Intron Biotechnology Inc., Korea) 을 사용하여 추출하였으며, universal primer인 27F primer (5'-AGAGTTTGATCMTGGCTCAG-3‘)와 1492R primer (5'-TACGGYTACCTTGTTACGACTT-3')를 사용하였 다. PCR은 genomic DNA를 주형으로 하여, PCR premix (GenDEPOT, Korea)를 이용하여 수행되었으며, 수행조건은 변성(denaturation) (94
oC, 1 분), 냉각(annealing) (60
oC, 1 분), 신장(elongation) (70
oC, 1분) 등의 단계를 33회 반복한 뒤, 70
oC에서 15분간 유지하였다. PCR을 통하여 증폭된 DNA 단편은 전기영동을 통해 확인한 후 agarose extraction kit (Intron, Korea)을 사용하여 gel로부터 회수 및 정제하였다.
염기서열은 ABI373 Automated sequencer (Foster City, CA,
USA)를 이용하여 결정하였으며, NCBI의 BLAST database
를 활용하여 상동성을 확인하였다. 분석된 16S rRNA 염기서
열은 Clustal X software (http://www.clustal.org)를 이용하여
정리하였으며 Molecular Evolutionary Genetic Analysis 7
(MEGA7; The Biodesign Institute, Tempe, AZ, USA) 소프트
웨어를 이용하여 계통수를 작성하였다.
2.5. 고추냉이 추출물의 준비
고추냉이 추출물은 강원도 철원군에서 구매한 고추냉이 뿌 리를 이용하여 준비하였다. 곱게 간 고추냉이 뿌리 50 g은 100 mL의 dichloromethane과 함께 30분간 완전히 풀어주었 으며, 이후 25
oC에서 1시간동안 교반하고, 15분간 초음파 처 리를 실시하였다. 그 후, 원심분리 (4,500 rpm, 30분, 4
oC)하 여 잔여물을 제외한 상등액을 취하였으며, 0.45 µm PVDF로 여과 및 추출하여 본 실험에 사용하였다 (Fig. 1). 이 고추냉 이 추출물은 무색투명하였으며, 고추냉이 특유의 톡 쏘는 향 을 가지고 있다. 준비된 고추냉이 추출물에 포함된 AITC의 함량을 측정하기 위하여 AITC 표준곡선을 준비하였으며, 표준용액은 AITC (Sigma Co., Steinheim, Germany)를 적정 농도로 희석하여 사용하였다 [17].
2.6. 고추냉이 추출물에 노출된 분리세균의 사멸률 조사 Bacillus cereus MK-11에 대한 고추냉이 추출물의 살균 효과 를 알아보기 위하여 분리균주를 LB 액체 배지에 배양하였다.
세균이 대수생장기에 도달하였을 때, 원심분리 (13,000 rpm, 4
oC, 10분)하여 팰릿을 얻었으며, PBS buffer로 3회 세척하였 다. 이 후, 원심분리를 동일조건 하에 재실시하여 얻어진 균 체를 다양한 농도 (0-2 mg/mL)의 고추냉이 추출물에 각각 노 출시켜 4시간 간격으로 LB 고체 평판배지에 100 µL씩 도말 하여 37
oC에서 18시간동안 배양하였다. 배양이 끝난 후, 형 성된 집락을 계수하여, 고추냉이 추출물의 노출시간과 농도 에 따른 분리세균의 사멸률을 각각 조사하였다.
2.7. 고추냉이 추출물에 노출된 분리세균의 alginate 생성 량 측정
고추냉이 추출물의 농도와 노출시간에 따른 분리세균의 alginate 의 생성량 변화를 측정하였다. Chang 등 [18]의 방법 으로 상등액에 포함된 alginate를 정제하여 측정하였다. 다양 한 농도의 고추냉이 추출물 (0-2 mg/mL)이 첨가된 LB 액체배
지에 분리세균을 접종하여 고추냉이 추출물에 노출시킨 후, 1시간 마다 상등액을 취하여 ice ethanol을 첨가한 후, −20
oC 에서 18시간 동안 처리하였다. 처리된 시료를 원심분리 (13,000 rpm, 4
oC, 60 분)하여 얻은 상등액을 동결건조 시킨 후, 증류수에 녹여 alginate를 얻었다. 얻은 alginate 시료 100 μL와 5% phenol solution 100 μL를 혼합한 후, 500 μL의 황산을 첨가한 뒤, 30초간 혼합하여 80
oC에서 30분간 반응시 켰다. 이 후, 반응시킨 튜브를 상온에서 식힌 뒤, 흡광도 (490 nm) 를 측정하여 alginate를 정량하였다. 정량은 sodium alginate (Sigma Co, St. Louis, MO)를 통해 표준곡선을 작성 하여 적용하였다. 표준곡선은 다양한 농도의 alginate 용액 (30, 60, 90, 120, 150, 180, 210, 240 µg/mL)을 제조하여 반응 시킨 뒤, 490 nm에서 흡광도를 측정하여 작성하였다.
2.8. 고추냉이 추출물 노출에 의한 SSPs 분석
고추냉이 추출물의 농도와 노출 시간에 따른 분리세균의 스 트레스 충격단백질(SSP) 변화를 관찰하기 위하여 분리세균 을 고추냉이 추출물에 노출시킨 후, 원심분리 (4,000 rpm, 4
oC, 10분)하여 균체를 얻었다. 얻어진 균체는 PBS buffer로 3회 세척한 후 PRO-PREP
™(Intron, Korea)에 균체를 현탁하 여 −20
oC에서 20분간 반응시켰다. 반응시킨 후, sonicator (Fisher Scientific, Pittsburgh, PA, USA) 로 30초씩 15회 파쇄 하여, 원심분리 (13,000 rpm, 4
oC, 40분)하였다. 그 후 상등액 만을 취하여 새로운 튜브에 옮겼으며 −70
oC에 보관하면서 본 실험에 사용하였다. 고추냉이 추출물에 노출된 분리세균 에서의 스트레스 충격단백질의 변화를 조사하기 위하여 SDS-PAGE 와 Western blot 분석을 실시하였다 [19]. Separating gel은 12% acrylamide gel을 사용하였으며, stacking gel은 8%
acrylamide gel을 사용하였다. 전기영동을 실시한 후, gel은 2.5% Coomassie brilliant blue staining solution (Bio-Rad Co., Hercules, CA, USA)으로 1시간 동안 염색하였다. Destaining solution I (50% methanol, 10% acetic acid) 로 30분씩 3회 탈 색하였고, destaining solution II (5% methanol, 7% acetic acid)로 1시간 동안 탈색하였다. 고추냉이 추출물에 의한 스 트레스 충격단백질(stress shock proteins, SSPs)의 분석은 Western blot을 통해 실시하였다. 1차 항체는 anti-DnaK (Abcam, Cambridge, UK) 와 anti-GroEL (Abcam, Cambridge, UK)을 사용하였으며, 2차 항체는 goat anti-mouse IgG H&L (HRP) (Abcam, Cambridge, UK)를 각각 사용하였다. 이 후 ECL kit (Intron, Korea)을 사용하여 X-선 필름 (AGFA, Belgim)에 감광한 후, 현상하여 SSPs의 발현을 분석하였다.
2.9. 주사전자현미경에 의한 세포 외부형태 관찰
고추냉이 추출물에 노출된 분리세균 MK-11의 외부 형태 변
화를 노출되지 않은 세포와 비교하기 위하여 주사전자현미
경으로 관찰하였다. 분리세균은 LB 액체배지에 배양하여 대
수생장기 때의 세균을 원심분리 (4,000 rpm, 10분, 4
oC) 를 통
해 확보하였다. 얻어진 균체는 PBS buffer로 3회 세척하여 준
비하였다. 이들 균체는 고추냉이 추출물 2 mg/mL의 농도에
Fig. 1. Flowchart for the preparation of wasabi extracts.
서 12시간 노출시켰다. 이후 균체는 원심분리를 통해 회수한 뒤, PBS로 3회 세척하였고 0.45 µm인 membrane filter로 분 리세균을 여과하여 membrane filter에 분리세균 MK-11을 부 착시켰다. 이 후 2.5% glutaraldehyde (Sigma Co., St. Louis, MO, USA)로 4시간동안 1차 고정하였으며, PBS buffer로 2 회 15분 동안 세척하였다. 2차 고정으로 1% osmium tetroxide (Sigma, USA)로 2시간 처리하였으며, PBS buffer로 2회 15분간 세척하였다. 탈수를 위해 다양한 농도의 ethanol (30, 50, 60, 70, 80, 95, 100%) 을 준비하여, 저 농도로부터 각 단계별로 10분씩 1회 탈수시켰다. 마지막 농도인 100%
ethanol의 경우에 3회 반복 탈수시켰다. 건조과정이 끝난 후, isoamyl acetate 와 ethanol이 1:1로 혼합한 시약으로 15분간 처리하고, 100% isoamyl acetate에서 4시간 동안 건조시켰다.
이후 코팅을 위하여 metal stub에 carbon tape를 부착하고 건 조된 membrane filter를 0.4 mm × 0.4 mm 크기로 잘라 붙여 두었다. Sputter coater (108 auto, Cressington Scientific Instru- ments, Watford, UK) 를 사용하여, 22 mA로 1분간 백금 코팅 하여 주사전자현미경 (JSM-7800F Prime, JEOL Ltd, Japan) 으로 세포의 외부형태를 관찰하였다 [20].
3. RESULTS AND DISCUSSION
3.1. 세균의 분리 및 배양조건
시판 중인 쌈장을 농화시킨 배양액으로부터 5% sheep blood 가 첨가된 blood agar에서 투명대가 형성되는 집락을 선별하 였으며, blood agar에서 3회에 걸쳐 도말을 통해 생장상태가 가장 양호한 세균인 MK-11을 최종 분리하였다. 분리세균은 LB 고체배지와 액체배지에 접종하였으며, 액체배지의 경우 진탕배양기 (160 rpm, 37
oC)에서 배양 및 유지하며 본 실험 에 사용하였다.
3.2. 분리세균의 생리 화학적 특성조사
분리세균 MK-11을 LB 고체 평판배지에 배양하여 형태학적 특성을 조사하였다. 집락은 밝은 흰색을 나타내었고, 표면은 거칠고 중심부가 솟았으며, 집락의 형태는 부정형으로 퍼져 있는 모습으로 나타났다. 그람염색을 하여 현미경으로 관찰 한 결과, 그람 양성의 간균으로 확인되었다. 분리세균 MK- 11에 대한 다양한 생리화학적 특성을 조사하였다. Table 1에 는 특성조사의 결과가 요약되어있다. Glucose 배지에서는 시 간이 경과함에 따라 붉은색이 노란색으로 변하고, 기체포집 관에 기체가 형성되어 양성반응을 보였다. Lactose는 이용되 지 않아서 음성임을 확인하였고, 젤라틴과 녹말 배지에서 모 두 투명대가 형성되어 양성을 나타내었다. 인돌 (indole) 생 성시험 결과는 음성으로 확인되었으며, Simmon’s citrate 시 험과 methyl red 시험에서는 양성, 그리고 Voges-Proskauer 시험에서는 음성 반응을 나타내었다. KIA 배지를 통해 H
2S 의 생성을 확인한 결과는 음성으로 나타났으며, litmus milk 시험에서 배지의 변화가 없어 음성으로 확인되었다. Catalase
시험은 양성으로, 그리고 oxidase 시험은 음성으로 나타났다.
3.3. 분리세균의 동정
분리세균 MK-11의 동정은 다양한 탄소원의 이용여부를 통 해 그람양성 세균의 동정에 사용되는 BIOLOG 분석시스템 을 사용하였다. 분석된 결과는 MicroLog
TMdatabase software 를 통해 확인한 결과, Bacillus cereus로 동정되었으며, Bacillus cereus MK-11로 명명하였다. BIOLOG 분석시스템 을 사용한 결과는 Table 2에 나타내었다.
3.4. 16S rRNA 염기서열의 계통수 분석
16S rRNA 염기서열 분석을 통하여 분리세균 MK-11의 동정 및 계통수 분석을 조사하였다. 결정된 염기서열은 NCBI의 Nucleotide BLAST 분석프로그램을 사용하여 상동성을 비교 하였으며, 그 결과로 Bacillus cereus와 99%의 유전적 상동성 을 나타냈다. 이를 바탕으로 B. cereus MK-11과 B. cereus 종 에 속하는 다른 균주들과의 유사성을 분자유전학적 계통수 로 나타내었다 (Fig. 2).
3.5. 고추냉이 추출물에 노출된 분리세균의 사멸률 조사 분리세균 MK-11에 다양한 농도와 시간으로 고추냉이 추출 물을 노출시켜 분리세균의 사멸률을 조사하였다. 고추냉이 추출물에 포함된 AITC의 양은 고추냉이 뿌리 50 g당 14.8 mg으로 측정되었으며, 본 실험에서는 이 추출물을 희석하여 사용하였다. 고추냉이 추출물의 농도와 시간이 증가할수록 MK-11의 사멸률은 증가하였다. MK-11 세균의 수는 0.5 mg/
mL 농도의 고추냉이 추출물에 24시간 노출 시, 초기 균수 (1
× 10
7CFU/mL) 와 비교하여 1.4 × 10
5CFU/mL, 1 mg/mL 농 도에서 1.3 × 10
3CFU/mL, 1.5 mg/mL의 농도에 24시간 노출 시 약 1.6×10 CFU/mL로 감소하였다. 고추냉이 추출물 2 mg/
Table 1. Morphological and physiological characteristics of isolate, MK-11
Morphological characteristics
Cell shape Rod
Gram Staining Positive
Physiological characteristics
Glucose fermentation +
Lactose fermentation
−
Gelatin hydrolysis +
Starch hydrolysis +
Indole production
−
Simmon's citrate +
Methyl red +
Voges-Proskauer
−
H2S (KIA)
−
Litmus milk (peptonization)
−
Oxidase
−
Catalase +
+ : Positive reaction, − : Negative reaction
Table 2. Biochemical characteristics of MK-11 using the BIOLOG Analysis System
Biochemical testsWater
−
D-Tagatose +α-Cyclodextrin + D-Trehalose +
β-Cyclodextrin + Turanose +
Dextrin + Xylitol
−
Glycogen + D-Xylose +
Inulin
−
Acetic aicd−
Mannan + α-Hydroxybutyric acid +
Tween 40 + β-Hydroxybutyric acid +
Tween 80 + γ-Hydroxybutyric acid
−
N-Acetyl-D-glucosamine + p-Hydroxyphenylacetic acid
−
N-Acetyl-β-D-mannosamine + α-Ketoglutaric acid
−
Amygdalin
−
α-Ketovaleric acid +L-Arabinose + Lactamide +
D-Arabitol
−
D-Lactic acid-methyl ester−
Arbutin
−
L-Lactic acid +D-Cellobiose + D-Malic acid
−
D-Fructose + L-Malic acid
−
L-Fucose
−
Pyruvatic acid metyl ester +D-Galactose
−
Succinic acid mono-metyl ester−
D-Galacturonic acid
−
Propionic acid−
Gentiobiose
−
Pyruvic acid +D-Gluconic acid
−
Succinamic acid−
α-D-Glucose + Succinic acid
−
m-Inositol + N-Acetyl-L-Glutamic acid
−
α-D-Lactose + L-Alaninamide
−
Lactulose
−
D-Alanine−
Maltose + L-Alanine +
Maltotriose + L-Alanyl-glycine +
D-Mannitol
−
L-Asparagine +D-Mannose
−
L-Glutamic acid−
D-Melezitose
−
Glycyl-L-Glutamic acid +D-Melibiose
−
L-Pyroglutamic acid−
α-Methyl-D-galactoside
−
L-Serine +β-Methyl-D-galactoside
−
Putrescine +3-Methyl-glucose + 2,3-Butanediol +
α-Methyl-D-glucoside + Glycerol +
β-Methyl-D-glucoside + Adenosine +
α-Methyl-D-mannoside
−
2'-Deoxy adenosine +Palatinose + Inosine +
D-Psicose + Thymidine +
D-Raffinose
−
Uridine +L-Rhamnose
−
Adenosine-5'-monophosphate +D-Ribose + Thymidine-5'-monophosphate +
Salicin
−
Uridine-5'-monophosphate +Sedoheptulosan
−
D-Fructose-6-phosphate +D-Sorbitol
−
α-D-Glucose-1-phosphate−
Stachyose
−
D-Glucose-6-phosphate−
Sucrose + D-L-α-Glycerol phosphate +
A single MK-11 colony grown on LB plate was streaked on BUG agar media containing 5% sheep blood and incubated 24h at 37
oC. Cells were
suspended in Sterilized Saline and inoculated into the GP2 MicroPlate
TM. After incubation for 24 h, the result was read with Biolog's automated
Micro-Station
TM.
mL 의 농도에서 분리세균이 노출될 시에 집락은 모두 사멸되 었다 (Fig. 3). B. cereus에 대한 AITC의 살균효과는 직접 AITC의 증기(vapor)를 노출시키는 방법 [21]과 AITC를 운반 체(carrier materials)로서 갈조류 또는 silica MCM-41에 흡착 시켜 이용하는 방법 [6]을 통해 각각 보고된 바 있다. Lu 등 [2] 은 Staphylococcus aureus B0031에 고추냉이 분말을 노출 시켰을 때, 초기 균수 10
5CFU/mL에서 12시간 후, 10
3CFU/
mL로 감소하였다. Masuda 등 [22]은 쥐의 위장에 감염시킨 Helicobacter pylori ATCC 43504에 대하여 고추냉이 잎 추출 물을 처리한 실험군과 처리하지 않은 대조군을 비교하여 실 험군에서 H. pylori의 집락수가 급격히 감소됨을 보고하였다.
본 연구결과에 따르면, MK-11은 2 mg/mL 농도의 고추냉이 추출물에 노출되었을 경우에 급격히 사멸하여 24시간 이후 에는 집락을 관찰할 수 없었다. 이를 통해 고추냉이 추출물 은 일정 농도 및 시간에서 B. cereus MK-11에 대한 뛰어난 살 균력을 지닌 것으로 나타났다.
3.6. 고추냉이 추출물에 노출된 분리세균의 alginate 생성 량 측정
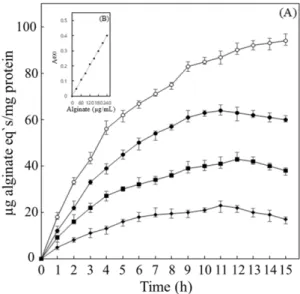
고추냉이 추출물에 노출된 분리세균 MK-11로부터 생성된 alginate의 양을 측정하였다. MK-11의 alginate 생성량은 고 추냉이 추출물의 노출 농도가 증가함에 따라 감소하였으며, 또한 일정 농도에서 노출 시간이 증가함에 따라 alginate 생 산량은 배양 11시간까지 지속적으로 증가하였으며, 그 후 서 서히 감소하는 경향을 보여주었다 (Fig. 4). 다양한 농도의 고 추냉이 추출물에 노출된 MK-11에서 algiante는 약 11시간 경 과 시에 최대생성량을 나타내었는데, 1.0 mg/mL에서 약 60 ug/mL, 1.5 mg/mL에서 약 38 ug/mL, 그리고 2.0 mg/mL에서 약 20 ug/mL로 각각 측정되었다. 그러나 고추냉이 추출물에 노출되지 않은 MK-11의 alginate 생성량은 배양 15시간까지 지속적으로 증가하였다. Alginate는 세포벽의 구성 물질로 음이온성의 다당류이며, 생물막 (biofilm) 형성, 건조방지, 세 포형태 유지, 외부환경으로부터 보호와 같은 생존에 중요한 요인으로 작용한다 [23]. B. cereus에서 AITC와 alginate 생성 과의 상관관계는 보고된 바가 없다. 다만 Kim 등 [11]이 B.
cereus에 녹차 카테킨 (catechin)인 EGCG를 노출시킨 결과, EGCG 의 농도가 증가됨에 따라 alginate 생성량이 감소됨을 보고하였다. 또한 alginate를 생성하는 세균으로 Pseudomonas 종(species)이 널리 알려져 있는데, Borges 등 [24]은 Ps.
aeruginosa,에 AITC를 노출시켰을 때 alginate가 주성분인 생 물막의 생성을 저해하고, 운동성을 감소시키는 것을 보고하 였다. Lee 등 [25]은 Ps. aeruginosa에 폭약 RDX를 처리한 결 과 노출 농도가 증가함에 따라 대사능력이 감소하고 외부 스 트레스요인에 취약해짐을 보고하였는데, 이들 결과는 대상 Fig. 2. Phylogenetic tree of the isolate, B. cereus MK-11 (●) based on 16S rRNA gene sequences. The tree was constructed by the neighbor-joining method. Number at nodes are bootstrap percentage based on 1000 resampled data sets. Bar, 0.005 substitutions per nucleotide position.
Fig. 3. Mortality rate of B. cereus MK-11 cells following
exposure to wasabi extracts. MK-11 cells were maintained at the
concentrations of 0 mg/mL ( ○), 0.5 mg/mL (●), 1 mg/mL (■),
1.5 mg/mL (▲), and 2 mg/mL (◆) wasabi extracts. At intervals,
the numbers of colonies (CFU/mL) were measured. Data shown
represent the mean±SD based on triplicate studies.
균주들이 천연 및 인공화합물에 노출됨으로서 alginate 생성 량의 감소에 기인하는 것으로 사료된다.
3.7. 고추냉이 추출물에 노출된 분리세균의 SSPs 발현조사 고추냉이 추출물에 노출된 MK-11의 스트레스 충격단백질 (예, DnaK와 GroEL)의 발현을 조사하였다. 먼저 일정시간 (24 시간) 동안 다양한 농도의 고추냉이 추출물에 노출시킨 MK-11에서 스트레스 충격단백질의 발현 패턴 (Fig. 5), 그리 고 일정한 농도 (2 mg/mL)에서 4시간 간격으로 최대 24시간 까지 노출시켜 이들 단백질의 발현 패턴을 각각 조사하였다 (Fig. 6). MK-11의 스트레스 충격단백질은 고추냉이 추출물
의 처리 시간 및 농도가 증가함에 따라 발현량은 감소하였다.
고추냉이 추출물을 다양한 농도로 처리하였을 때, DnaK와 GroEL의 발현은 0.5 mg/mL 농도에서 대조군과 비교하여 유 사한 발현량을 보였으나 2 mg/mL의 농도에서는 대조군에 비하여 확실한 감소를 보여주었다. 다양한 시간으로 처리된 결과, 대조군과 비교하여 4시간 경과 시에 DnaK와 GroEL의 발현량은 유사하였으며, 4시간 이상 노출된 경우에서 발현 량은 점차 감소하여 24시간 경과 시에 가장 낮았다. 스트레 스 충격단백질인 DnaK와 GroEL은 Escherichia coli에서 가 장 먼저 발견되어 널리 연구되었으며, 온도, pH, NaCl 등의 스트레스 환경에서 변성된 단백질의 3차 구조를 회복시키는 것으로 보고되었다 [26]. 고추냉이 추출물에 의한 노출이 세 균의 스트레스 충격단백질 반응과 관련된 연구는 알려진 바 가 거의 없다. Periago 등 [27]은 고온 (42 - 50
oC)에 노출된 B.
cereus에서 DnaK와 GroEL의 존재 및 발현을 확인하였으며, 고온 환경에서 노출 시간에 따라 발현량이 변화함을 보고하 였다. Cho 등 [28]은 스트레스 요인으로 녹차폴리페놀 (tea polyphenols)을 처리한 MRSA에서 DnaK와 GroEL의 발현이 감소됨을 발표하였다. 본 연구는 고추냉이 추출물이 대상 세 균 (예, Bacillus cereus)에 대하여 스트레스로 작용하여 충격 단백질을 생성한다는 최초의 보고로서, 스트레스 요인으로 고추냉이 추출물의 노출농도 증가 또는 노출시간 지속에 따 른 MK-11의 DnaK 및 GroEL의 발현이 점차적으로 감소하여 궁극적으로 사멸되는 것으로 생각된다.
3.8. 주사전자현미경에 의한 세포 외부형태 관찰
고추냉이 추출물의 노출에 따른 분리세균 MK-11의 외부형 태를 주사전자현미경을 통해 관찰하였다. 고추냉이 추출물 을 처리하지 않은 MK-11 세포는 표면이 매끄럽고 정상적이 고 전형적인 간균의 형태를 보여주었다. 그러나 2 mg/mL 농 도의 고추냉이 추출물을 12시간 처리한 MK-11의 세포의 표 면은 매끄럽지 못하고 움푹 패인 형태로 관찰되었다 (Fig. 7).
고추냉이 추출물에 노출에 의한 B. cereus의 세포 외부형태
Fig. 5. Western blot analysis of stress shock proteins (SSPs) production in in B. cereus MK-11 treated with different concentrations of wasabi extracts for 24 h; 0 mg/mL (lane 1), 0.5 mg/mL (lane 2), 1 mg/mL (lane 3), 1.5 mg/mL (lane 4), 2 mg/mL (lane 5), The SSPs were analyzed by SDS-PAGE (A), and western blot with anti-DnaK (B), and anti-GroEL (C) monoclonal antibodies, respectively.
Fig. 4. Exopolymer production as alginate equivalents by B.
cereus MK-11. (A) The cells were grown in LB broth only (○), LB broth in the presence of wasabi extract 1 mg/mL ( ●), 1.5 mg/
mL ( ■), 2 mg/mL (◆). (B) The standard curve was prepared to determine alginate quantitatively. The results are the means±
standard errors of three independent experiments.
를 관찰한 연구 결과는 거의 보고된 바 없다. 다만 Kim 등 [14]은 시판 쌈장에서 분리한 용혈성 B. cereus MH-2를 NaCl 에 노출시킨 후, 주사전자현미경을 통하여 균주의 세포 표면 에 구멍이 뚫리고 심하게 쭈그러진 불규칙한 형태로 관찰된 다는 것을 보고하였다. 본 연구를 통해서 동일한 종(種)의 B.
cereus라 할지라도, 노출 화합물의 종류에 따라 대상 세포의 외부 형태도 달라짐이 분명히 입증되었다. Maruthupandy와 Seo [29]는 polyacrylate로 캡슐화된 AITC를 E. coli에 처리하 여 세포 형태 및 외막이 변형되었음을 보고하였다. Lin 등 [30]은 AITC를 처리한 Salmonella enterica serovar Montevideo, Escherichia coli, Listeria monocytogenes 등에서 세포막의 손 상과 세포 내 대사물질이 유출되어 세포가 사멸된다는데 기 반을 둔 항균메커니즘을 제시하였다. 그러나 고추냉이 추출 물이 대상 균주들에 대하여 세포벽 (cell wall) 또는 세포막 (cell membrane)에서 작용하는지에 대해서는 분명하지 않으 며, 이를 규명하기 위해서 더욱 상세한 연구가 진행되어야 할 것으로 사료된다.
우리나라의 전통 발효 시판 장류 제품에서 B. cereus는 식 품 안전성 차원에서 지속적으로 문제가 제기되어왔으며, 이 에 대하여 2007년부터 한국식품의약품안전처에서는 이 균 주에 대한 허용 기준을 설정하여 관리해 오고 있다. 그러나
이 문제가 여전히 해결되지 않는 이유는 장류에 존재하는 B.
cereus가 생성하는 내생포자를 효율적으로 제거하는 방안이 맛이나 향 등의 성상 (性狀)을 유지하는데 적합하지 못하기 때문일 것으로 생각된다. 지금까지 고추냉이 추출물에 의한 B. cereus에 의한 증식억제는 거의 보고된 바가 없다. 이러한 점들을 종합해 볼 때, 고추냉이 추출물은 맛과 풍미 등의 성 상 변화를 최소화 하면서, B. cereus 증식억제를 위한 식품보 존제로서 경쟁력이 있는 방법으로 제시될 수 있을 것으로 사 료된다.
4. CONCLUSION
본 연구에서는 시판 중인 쌈장에서 분리한 B. cereus MK-11 을 고추냉이 추출물에 노출시켜 여러 가지 생리 및 세포반응 을 조사하였다. 분리세균에 대하여 다양한 생리화학적 특성 조사를 실시하였으며, BIOLOG 분석시스템으로 B. cereus로 동정하고, B. cereus MK-11로 명명하였다. MK-11 균주의 16S rRNA 염기서열 분석을 수행하여, 분자유전학적 계통수 를 작성하였다. 다양한 농도의 고추냉이 추출물에 노출된 MK-11의 사멸률은 추출물의 농도 및 노출시간이 증가함에 따라 증가하였으며, 2 mg/mL의 농도에서 24시간 처리 시에 더 이상의 집락은 관찰되지 않았다. 고추냉이 추출물에 의한 MK-11 의 alginate 생성량은 추출물의 농도가 증가함에 따라 감소하였으며, 노출시간에 따라 증가되었다. Western blot을 통해 고추냉이 추출물에 의한 스트레스 충격단백질의 발현 은 추출물의 농도 및 시간이 증가함에 따라 스트레스 충격단 백질(DnaK와 GroEL)은 감소하는 것으로 나타났다. 고추냉 이 추출물에 노출된 MK-11의 세포 외부를 주사전자현미경 을 통해 관찰한 결과, 세포 표면이 형태학적으로 크게 변화 된 모습을 보여주었다. 결론적으로, 본 연구는 고추냉이 추 출물의 노출에 의한 B. cereus의 세포독성 메커니즘을 이해 하는데 중요한 단서가 될 것이다.
Fig. 6. Western blot analysis of stress shock proteins (SSPs) production in B. cereus MK-11 treated with 2 mg/mL wasabi extracts for different exposure times (h). The SSPs were analyzed by SDS-PAGE (A), and western blot with anti-DnaK (B), and anti-GroEL (C) monoclonal antibodies, respectively.
Fig. 7. Scanning electron micrographs of (A) B. cereus MK-11
untreated cells, (B) cells treated with
wasabi extractsfor 12
hours.
Acknowledgements
본 연구는 순천향대학교 학술연구비지원사업의 일부지원으 로 수행되었음.
REFERENCES
1. Kim S. W., S. J Moon, and B. M. Popkin (2000) The nutrition transition in South Korea. Am. J. Clin. Nutr. 71: 44-53.
2. Lu, Z., C. R. Dockery, M. Crosby, K. Chavarria, B. Patterson, and M. Giedd (2016) Antibacterial activities of wasabi against Esche- richia coli O157:H7 and Staphylococcus aureus. Front. Micro- biol. 7: 1403-1411.
3. Kumagai, H., N. Kashima, T. Seki, H. Sakurai, K. Ishii, and T.
Ariga (1994) Analysis of volatile components in essential oil of upland wasabi and their inhibitory effects on platelet aggregation.
Biosci. Biotech. Biochem. 58: 2131-2135.
4. Zhang, Y. (2010) Allyl isothiocyanate as a cancer chemopreven- tive phytochemical. Mol. Nutr. Food Res. 54: 127-135.
5. Shofran, B. G., S. T. Purrington, F. Breidt, and H. P. Fleming (1998) Antimicrobial properties of sinigrin and its hydrolysis prod- ucts. J. Food Sci. 63: 621-624.
6. Siahaan E. A., A. Meillisa, H. C. Woo, C. W. Lee, J.H. Han, and B. S. Chun (2013) Controlled release of allyl isothiocyanate from brown algae Laminaria japonica and mesoporous silica MCM-41 for inhibiting food-borne bacteria Food Sci. Biotechnol. 22(S): 19- 24.
7. Kim, H. S. (2020) Cytotoxicity and cellular responses of enteric bacteria, Escherichia coli MK-7 and Salmonella enterica MK-9 exposed to wasabi extracts. MS Thesis. Soonchunhyang Univer- sity, Asan, Chungnam, Korea.
8. Sabra, W., A. P. Zeng, and W. D. Deckwer (2001) Bacterial algi- nate: physiology, product quality and process aspects. Appl. Micro- biol. Biotechnol. 56: 315-325.
9. Nwodo, U. U., E. Green, and A. I. Okoh (2012) Bacterial exopoly- saccharides: functionality and prospects. Int. J. Mol. Sci. 13:
14002-14015.
10. Pier, G. B. (1998) Pseudomonas aeruginosa: a key problem in cys- tic fibrosis. ASM News 64: 339-347.
11. Kim, D. M., S. K. Park, and K. H. Oh (2016) Cellular responses and proteomic analysis of hemolytic Bacillus cereus MH-2 exposed to epigallocatechin gallate (EGCG). Korean J. Microbiol.
52: 260-268.
12. Granum, P. E. and T. Lund (1997) Bacillus cereus and its food poi- soning toxins. FEMS Microbiol. Lett. 157: 223-228.
13. Kim, S. K., K. P. Kim, S. S. Jang, E. M. Shin, M. J. Kim, S. Oh, and S. Ryu (2009) Prevalence and toxigenic profiles of Bacillus cereus isolated from dried red peppers, rice, and sunsik in Korea. J.
Food Prot. 72: 578-582.
14. Kim, D. M., S. K. Park, and K. H. Oh (2017) Identification and characterization of hemolytic Bacillus cereus isolated from com- mercial ssam-jang. KSBB. J. 32: 179-186.
15. Dierick, K., E. Van Coillie, I. Swiecicka, G. Meyfroidt, H.
Devlieger, A. Meulemans, G. Hoedemaekers, L. Fourie, M. Heyn- drickx, and J. Mahillon (2005) Fatal family outbreak of Bacillus cereus-associated food poisoning. J. Clin. Microbiol. 43: 4277- 4279.
16. Krueger, W. B. and B. J. Kolodziej (1986) Laboratory Procedures for General Microbiology. Kendall / Hunt Publishing Company, IA, USA.
17. Hyun, J. H. and J. M. Choi (2012) Spectrophotometric determina- tion of allyl isothiocyanate after ultrasonic solvent extraction. J.
Kor. Soc. Environ. Anal. 15: 234-238.
18. Chang, W. S., M. Van De Mortel, L. Nielsen, G. Nino De Guz- man, X. Li, and L. J. Halverson (2007) Alginate production by Pseudomonas putida creates a hydrated microenvironment and contributes to biofilm architecture and stress tolerance under water-limiting conditions. J. Bacteriol. 189: 8290-8299.
19. Bollag, D. M., M. D. Rozycki, and S. J. Edelstein (1996) Protein Methods. 2nd ed. Wiley-Liss, NY, USA.
20. Ho, E. M., H. W. Chang, S. I. Kim, H. Y. Kahng, and K. H. Oh (2004) Analysis of TNT (2,4,6-trinitrotoluene)-inducible cellular responses and stress shock proteome in Stenotrophomonas sp. OK- 5. Curr. Microbiol. 49: 346-352.
21. Isshiki, K., K. Tokuoka, R. Mori, and S. Chiba (1992) Preliminary examination of allyl isothiocyanate vapor for food preservation.
Biosci. Biotechnol. Biochem. 56: 1476-1477.
22. Masuda, S., H. Masuda, Y. Shimamura, C. Sugiyama, and F. Taka- bayashi (2017) Improvement effects of wasabi (wasabia japon- ica) leaves and allyl isothiocyanate on stomach lesions of mongolian gerbils infected with Helicobacter pylori. Nat. Prod.
Commun. 12: 595-598.
23. Boyd, A. and A. M. Chakrabarty (1995) Pseudomonas aerugi- nosa biofilms: role of the alginate exopolysaccharide. J. Ind.
Microbiol. 15: 162-168.
24. Borges, A., L. C. Simões, M. J. Saavedra, and M. Simões (2014) The action of selected isothiocyanates on bacterial biofilm preven- tion and control. Int. Biodeterior. Biodegrad. 86: 25-33.
25. Lee, B. U., S. C. Park, Y. S. Cho, and K. H. Oh (2008) Exopoly- mer biosynthesis and proteomic changes of Pseudomonas sp. HK- 6 under stress of TNT (2,4,6-trinitrotoluene). Curr. Microbiol. 57:
477-483.
26. Chuang, S. E. and F. R. Blatiber (1993) Characterization of twenty-six new heat shock genes of Escherichia coli. J. Bacteriol.
175: 5242-5252.
27. Periago, P. M., W. Van Schaik, T. Abee, and J. A. Wouters (2002) Identification of proteins involved in the heat stress response of Bacillus cereus ATCC 14579. Appl. Environ. Microbiol. 68: 3486- 3495.
28. Cho, Y. S., N. L. Schiller, and K. H. Oh (2008) Antibacterial effects of green tea polyphenols on clinical isolates of methicillin- resistant Staphylococcus aureus. Curr. Microbiol. 57: 542-546.
29. Maruthupandy, M., and J. Seo (2019) Allyl isothiocyanate encap- sulated halloysite covered with polyacrylate as a potential antibac- terial agent against food spoilage bacteria. Mater. Sci. & Eng. C.
105: 1-9.
30. Lin, C. M., J. F. Preston 3rd, and C. I. Wei (2000) Antibacterial mechanism of allyl isothiocyanate. J. Food Prot. 63: 727-734.