약학회 지 제 4 4 권 제 2 호 197~203(2000)
Yakhak Hoeji
Vol. 44, No. 2A s i a l o f e t u i n 에 대 한
A s p e r g i l l u s o r y z a e ,
b o v i n e l i v e r ,S a c c h a r o m y c e s f r a g i l i s
유 래 P - g a l a c t o s i d a s e 의 반 응 조 건윤재경
•
이영재*
구본응•
윤상영•
유창수 중앙대학교 약학대학 (Received March 21, 2000)김하형
"
T h e R e a c tio n C o n d itio n s o f (3 - G a la c to s id a s e s fr o m A sp erg illu s o ry za e, B o v in e L iv e r , a n d S a cch a ro m y ces fra g ilis to A s ia lo fe tu in
Jae-Kyoung Youn, Young-Jae Lee, Bon-Woong Koo, Sang-Young Youn, Chang-Soo Ryu and HaHyung Kim^
College of Pharmacy, Chung-Ang University
A b s tra ct —— The enzymatic properties of p-galactosidases from
Aspergillus oryzae,
bovine liver andSac
charomyces fragilis
have been studied using enzyme-linked lectin assay based on the RCA1 20 and BS-II lectins which specifically bind to terminal galactose and GlcNAc residue, respectively. Asialofetuin, a monomeric glycoprotein w ith approximately 48 kDa in molecular weight, was used as a substrate. T his gly
coprotein contains three A^-linked triantennary complex type carbohydrate chains w ith each of which te r
minating in G alpl 4GlcNAc (74%). Their optimal pHs were 3.5 and 6.5 (A
oryzae),
and 3.5~5.5 (bovine liver andS. fragilis)
at 37°C during 24 hrs, and the effective concentrations were 0.9, 2.9, and 1.7 mg/m/, respectively. The enzyme fromA. oryzae
requires 100 mM or K"^, while the enzyme from bovine liver requires for activity. However all of the three p-galactosidases were inactivated by SDS and These results indicate that the hydrolysis of glycoprotein such as asialofetuin depends on the reaction conditions of p-galactosidases and some metal ions.
K eyw ord s □ |3-galactosidase, activity, asialofetuin, lectin.
당말단 가수분해효소
(exoglycosidase)
는 복합당질(glycoconjugate)
의 다당의 비환원 말단(nonreducing end)
으로부터 단당 단위로 가수분해하는효소의 총청 으?-
순차적 분해(sequential degradation)
에의해 다 당의 일차구조 해석과 각 결합 단당의 아노머결합을 결정하는데 널러 이용되고 있다.^^ P-galactosidase((3- D-galactoside galactohydrolase, EC 3.2.1.23)
는glu- cose-P-D-galactoside
로 된 올리고당 배열로 부터galactose
률유리시키는 당말단가수분해효소의 일중으 로,
골지체에서 올러고당^ 의 합성시 발현하며세균본^논문에 관한 문의는 이 저자세게로 02-820-5612 02-820-5606
에서 고등동식물에이르기까지 다양하게 분포하고
,
주 용도로는다당의 구조연구,
유전공학에서 클론 선택의 마커,
융합유전자 형성에 이용될 뿐만 아니라 유아의 유당불내에 의한 소화불량의 개선및 경 구 우 ^등 의 섭취시 유당불내에 의한설사에 유용하여 소화제로사*
용되기도 한다.^^
최근
Aspergillus niger, Streptococcal 6646K, Dip- lococcus pneumoniae, Jack-bean 유래의 p-galactosi- dase
등이 분리되어 이용되고 있으며,
이들은 각각 기
질특이성,
빈 최 적 pH,
유효농도,
반응온도 및이온
둥에 의한저해 흑은 활 성 작 ^에 어가 나는 것으
로보고되고 있다.^오
기질 특이성에 있어서는 주로
Galp ^ 4GlcNAc
의197
윤재경 • 이영재 • 구본웅 • 윤상^ • • 김하형
( a )
Gal
Pl->«4GlcNAc
01—2Man
a1
G a ly? l-^4 G lcN A c^lv
Gal; Sl
나4GlcNAc;91^
Man
$l~»4GlcNAc/31—»4GlcNAc/31—>Asn
(b) (c)
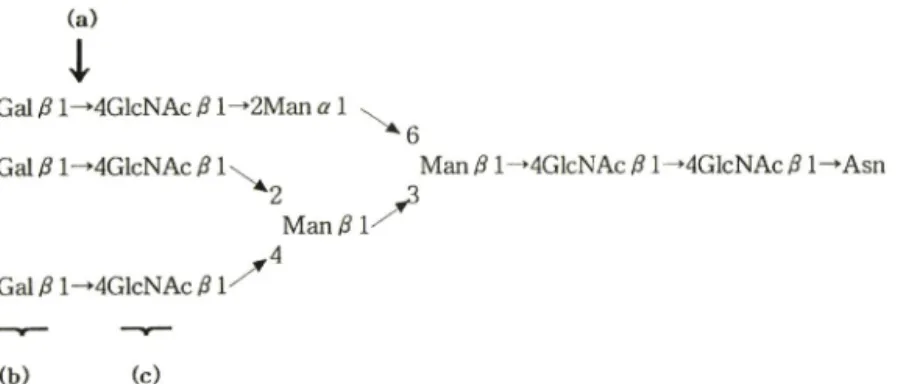
Fig. 1 - Structure of the major TV-linked triantennary complex-type oligosaccharide of asialofetuin. (a) indicates the site of hydrolysis of p-galactosidase. (b) and (c) indicate
respectively.
the site of binding specificity of RCA, 2 0 and BS-II lectin,
결합을 가수분해하는 것으로 알려져 있으나, strep
tococcal
6646K, Jack bean유래의 경우에는
Gaip3G lcN Ac
의 결합도 가수분해하는 것으로 알려져 있으
며, 반응 최적
pH는
Aniger 유래
p H 4 0 —4.5,Strep
tococcal
6646K유래
p H 5.5,D. pneumoniae 유래
p H 6 .0 -6 .5 , Jack bean유래
p H 3 .5 ~ 4 .5인 것으 로 보고되고 있고, 반응•온도는
37°C,빈움시간은
18시 간이 최적 빈움조건으로 알려져 있다.®> 또한, Strepto
coccal
6646K유래는
E D T A에 의해서는 반용이 저해 되나, Mn^■"이온등에 의해서는 활성화되는 결과가 보 고되고 었다7* 그러나, Aspergillus oryzae,
bovine liver,Saccharomyces fragilis 유태의 경우 대량으로 정제하는 방법이 보고되고 었으나,*> 반응조건은 기질 을
/M iitro p h e n y l (3-D-galactoside와 같은 저분자
glycon
을 이용하여 이루어지고 있으며, 당단백질이나
당지질과 같은 복합당질을 대상으로 한 보고는 미미한 실정이다.
당말단 가수분해효소의 반응조건을 확인하기 위한 방법으로는 colorim etric assay법
,®*
synthetic chromo- genic substrate로 부터 유리되어 나오는 chrom ogen 의 즉정,"*지 synthetic fluorescent substrate로 부터유리되어 나오는 형광도의 측정
,
tritium -labe le doligosaccharide substrate를이용한 radioisotopic assay
법,*각le c tin울이용한 방 법 등 의 이용되고 있다
.
이 중
,
lectin을 이용한 경우,
형광검출기의 이용,
radioisotope을사§■하여야히는불편함등을극복하고
,
특정당과결합하는
^ 1
적인성질로 당구조분석이나 세포표면의 당단백질혹은 당지질의 구조,
기능연구 에많이 이용되고 있다. * ^
특히,
당말단7H
수분해효소 중 가장 연구가 활발한
neuraminidase의 반응성에 관한 연구에
le ctin의 당걸합 특이성을 이용한 결과가 보고되고 있다.**^
본
연구에서는 기질로 당단백질의 일종인
asialofe- tu in을이용하여
Aoryzae,
bovine liver,S. fragilis 유래의
p-galactosidase에 대한 반응최적
pH ,유효농 도, 반응적정온도, 반응시간, 번성제 및 금속이온등의
영향을
lectin의 당 결합 특이성을 이용하여 실시하였
다.
A sia lo fe tu in은
fe tu in의 버환원 말단으로부터
sialic acid틀 가수분해하여
G a lp l나
4G lcNAc이 말단인
iV -linked trian te nn ary com plex-type의 당쇄가 약
74%, M a n a l— 3가지중
G a ip i나
3G lcN AC을 포함하 는 이성체가 약
9%, G aipi-->4G lcN Ac잔기률 가지는
biantennary chain및
G a ip i-< 3 G a lN A c의
0 -lin k e d disaccharide를 일부 포함하는 분자량
48 kD a의 당단 백질로 대표적인 당의 구조는
Fig. 1에 나타내었다
꾸2 0> 또 한 ^ 본연구에서 이용한
le c tin은 Ricinus communis 유래로 당말단의
galactose률 특이적으로 결합하는 RCA|
2o과 Bandeiraea simplicifolia 유래로 당말단의
G lcN A c과 결합하는
B S -II이며, 이들
le c tin의당 결합 특이성을 이용하여
M icroplate Reader를 이용한
enzym e-linked lection assay법을적용하였다■ 고 ** 즉,
P-galactosidase를 기질에 반응시키 면
galactose가7|
수분해되어 새로운 당
말단인 GlcNAc과 결합하는
peroxidase-labeled B S -II lectin이반응
을 하게 되므로발색제로 발색한
후 415 nm에서 흡광도률 측정할 수 었게 되며
,
p-galactosidase에 의해가수분해
반응이
진행되지 않은경우에는
galactose에 혁의적으로
결힘하는RCA,m
이번응하게된다(Fig. 1).p-galactosidase 의 빈움 조건 199
실 험방 법
시약 및 기기 -
본 실험에서 사용한asialofetuin (fetal calf serum), bovine serum albumin(BSA), polyethylene sorbitan monolaurate(Tween 20), pero
xidase-labeled RCAj2o \ec\m{Ricinus communis ag
glutinin), peroxidase-labeled BS-H \ectin(Bandeiraea simplicifolia), ^gala.ciosi6ase(/\spergilltts oryme, bovine liver, Saccharomyces fragilis), 2,2'-azinobis(3-ethyl- benzthiazoline-G-sulfonic add)(ABTS), sodium dodeccyl sulfate(SDS
)는SIGMA
사로부터 구입하였다.
그외의 시약은 모두 특급 흑은 일급을 사용하였다. P- galactosidase
반응성 확인에는Microplate Reader (Model 550, Bio-Rad
)를사S'
하였으며,
단백질의 정량 에는Cary 3 UV/VIS Spectrophotometer(Varian)
률 사 하 였 다.
Asialofetuin에 대한 lectin의 결할 농도 착인 - Asialofetuin
을0.625, 1.25, 2.5, 5, 10, 20 |xg/m
/의 농도로10 mM phosphate buffer, 0.15M NaCl, pH 7,4(PB
욕)에 용해하고 각각을50
씩mkroplate well
에 코팅한 후37T
에서1
시간 동안 반응시켰다.
코팅된 용액은aspiration
에의해 제거하고 각well
은PBS-0.1% Tween 20(PBST) 100 pJS. 3
회 세척하 였다.
그 후1% BSA
를 포함한PBST 100 pi
를well
에 넣고37°C
에서1
시간 반응시킨 후aspiration
에 의해 제거하고 각well
을100 PBST
로3
회 세척하였다.
그리고10, 25, 50 |Vm
/의 농도로PBS
에용해한peroxidase-labeled lectin RCA^o
과pero
xidase-labeled lectin BS-II
를50
씩 각well
에 넣 고37°C
에서1
시간동안 반응시키고, 100
의PBST
로5
회 세적후ABTS 0.55mg/m/
용액100m
/에H2O2 1 p
/률 넣고 각well
에100
썩 넣은후 즉시Microplate Reader
를사용하여415 nm
에서 각well
의흡광도 값을측정하고10
분후 재측정하여 각각의 상대값을 구하식 시약을 넣는 순서에 의한 흡광도의 오차를즐였다.
P-galactosidaseS| 반응성 확인 - Asialofetuin
의농 도는5 |xg/m/, peroxidase-labeled lectin BS-II
의 능 도는10 [ig/m
/로 하여asialofetuin
에 대한lectin
의 결합방법에준하여 실시하였으나, lectin
을가하기 전 에 다옴에 나타내는 각 조건에 따라p-Galacto- sidase(A. oryzae, bovine liver, S. fragilis)# 'il'i
•시켰다
.
완충액의 조제
-본 실험에서 시용한 완충액은100 mM citric acid
에200 mM Na2HP04
■틀29.4%, 44.9%, 56.3%, 70.9%, 92.0
%씩 각각 가하늬pH
룰3.5, 4.5, 5.5, 6.5, 7.5
로조절하였다.
P"galactosidase의 최적 pH 조건
一pH 3.5, 4.5, 5.5, 6.5, 7.5
의 완중액에 용해한p-galactosidase
를0.8 unit/100 10/
농도로 하고37°C
에서24
시간동안 기질과 반응시킨 후반응성을 확인하였다.
p-galactosidase^ 유효농도 및 반응 적정온도 - P- galactosidase
를pH 3.5
의 완충액에0.05, 0.1, 0.2, 0.4, 0.8 unit
률100 p
/에 용해한 후37°C
에서24
시 간동안 반응시킨 후 반응성을 확인하였다.
반응은20°C
와37X
로 한incubator
에서 실시하였다.
P-galactosidase의 반등시간 oryzae, S. fragilis
유래 P-galactosidase
인 경우에는 0.8 unit/100 \xl, bovine liver
유래 P-galactosidase
인 경우에는 0.05 unit/100 ^i
/로 한 후 2, 8, 16, 24
시간 반응시킨 후
37^C
에서 활성을 측정하였다.
변성제 및 금속시약어 p-galactosidase의 활성에 미 치는 영향 -
단백질번성제로는SDS,
금속이온으로는KC1, NaCl, C11CI2
,MnCl2, MgCl2, CaCl^, BaCl^, ZnCl2, C0CI2, FeCl2, FeClg
를 각각 최중농도가0.1, 1, 10, 100 mM
이되도록가하고 A ofyzae, S. fragilis
유래인 경우에는 0.8 unit/100 |oi, bovine liver
유래인
경우에는 0.05 imit/100
빠 한 후 pH 3.5
의 완층
액에서 37X, 24
시간 반응•을시킨후 반응성을 확인하
였다. p-galactosidase
의상대적인 활성도는 시약을가
하지 않은 경우의 흡광도률
100
%로 하고 얻어지는흡광도의 상대적인 %룰 구하였다
.
실 험 결 과 및 고 찰
Asialofetuin에 대한 lectin의 결합 농도 - Asialofe- tuin
의 농도률0.625~20 |xg/m
/로 희석하고lectin
의 농도률10, 25, 50
으로 하여415 nm
에서Microplate Reader
률 이용하식 흡광도률 측정하였다.
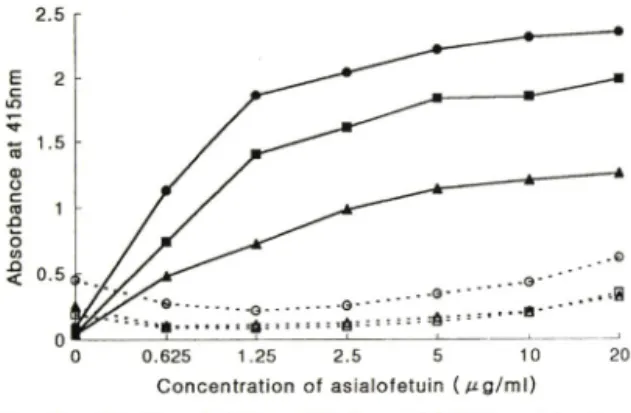
그 결과, asialofetuin
의 비환원 말단에 존재하는galactose
와의 특이적으로 결함하는RCAi^o
과의 반응 에서는asialofetuin
의농도가증가함에 따라흡광도도 증가하였다.
이는asialofetuin
의경우Gaipi-4GlcNAc
이버환원말단에 결합되어galactose
가표면에 노출되Vol. 44, No. 2, 2000
5.5 pH
Fig. 3 - Effect of pH on the activity of p-galactosidases from
A. oryzae
( • ) , bovine liver (■ ) andS.
fragilis
(A ) at 37*'C for 24 hrs. Each value represents mean ± S.D. (n=3).0 0.05 0.1 0.2 0.4 0.8
C o n c e n tr a tio n ( U n it)
Fig. 4 - Effect of concentration and temperature on the activity of p-galactosidases from
A. oryzae
( • ) , bovine liver (■ ) andS. fragilis
( A ) at 3 7 X (fillled) and2(fC
(open). The protein concentration represented 9.0, 0.17 and 4.6 unit/mg protein. Each value represents mean + S.D.
(n=3).
산하면 oryzae, bovine liver, S, fragilis 유래 9.0, 0.17, 4.6 unit가 각각 효소 1 mg에 해당하고, 반응성 이 높은 0.8, 0.05, 0.8 unit률 유효농도로 다시 환산 하면 각각 0.09, 0.29, 0.17 mg/100 ^i/에 해당한다.
1
unit 이상의 경우에는 p-galactosidase가 완충액에 완전히 용해되지 않아 실험을 실시하지 않았다. 또한, 같은 실험을 20X 에서 실시한 결과, 37X에 비해 반 응이 거의 이루어지지 않았옴을 알 수 있었다.
p"galactosidase의 반응 서 간 - p-galactosidase를 2,
8, 16, 24시간 반웅시킨 걸과, 빈움시간이 길어질수 록 반응성이 높게 나왔다(Fig. 5). 이는 P-galac- tosidase가 반응을 나타내는데에는 24시간 흑은 그 이
0 0.625 1.25 2.5 5 10 20
C o n c e n tra tio n of a sialo fe tu in ( ^ g / m l )
F ig. 2 - Binding of RCA, 2 0 (filled) and BS-II (open) lectin to asialofetuin as a concentration of 5 0 ( 0 ) , 25(B), and 1 0( A ) ^g/m/.
어 있으므로
R C A ,20과 galactose가 강한 결합력을 나 타낸 것을 나타내며, GlcNAc과 결합하는 BS-II는 asialofetuin과 거의 결합을 하지 않은 것을 알 수 있 었다(Fig. 2). 이 결과에 의해 비교적 낮은 농도의 asialofetuin 의 경우에도 RCAj^o 은 asialofetuin 의 galactose와 결합력을 나타낸다는 것을 알 수 있었고, 이하 asialofetuin의 농도는 5 ^g/m/, lectin의 농도는
1 0
로 s H 실험을 실시하였다.
^galactosidase 의 반등 최적 pH - pH 3.5, 4.5, 5.5, 6.5, 7.5의 각 완충액에 용해하겨 p-galactosidase 의 빈움성을 확인한 결과, A oryzae 유래는 pH 3.5 와 pH 6.5에서 비교적 높은 반응성을 나타내었으며, bovine liver, S. fragilis 유래는 pH 3.5, 4.5, 5.5에 서는 비숫한 반응성을 나타낸 반면 pH 6.5이상서1서는 현저하게 번움성이 저하하였다(Fig. 3). 이 결과로 앞 에서 서술한 다른 유래의 P-galactosidase가 반응을 나타내는 데에 최적 pH가 그 기원에 따라 처의가 나 는 것과 같미 본 효소들도 적정 pH가 각각 다른 것 을 확인하였다.
P-galactosidase의 유효농도 및 적정온도 -(i- galactosidase를 각각 0, 0.05, 0.1, 0.2, 0.4, 0.8 unit로 하여 37T에서 반응시킨 결과, A oryzae, S.
fragilis 유래는 농도가 높아질수록 반응성이 높아지는 결과가 나타냈으나, bovine liver 유래의 경우에는 0.05 unit에서 반응성이 가장 높았으며 그 이상으로 농도가 커져도 큰 차이가 없었다(Fig. 4). 여기에서 나 타내는 unit는 1.0 |xm이의 o-nitrophenyl (3-D-ga- lactoside를 o-nitrophenol과 Z)-galactose로 가수분해하 는 양을
1unit로 하였으며, unit를 효소의 농도로 환
200 윤재경 • 이영재 • 구본융 * 은상영 • TT청수 • 김하형
5 3 5 2 5 1
3 K 2 h 1 K
0
• 0
0 0 0
0
^ l Eu
얏In o u u« q j o u q<
^ l Eu
.학Ira u« qJO »c 0o
셔<
Eu g nr
la 9U
U
법qJO OT qv
p-galactosidase 의 빈융 조건 201
Table I - Continued
R e a c tio n tim e (h r)
Fig. 5 - Effect of reaction time on the activity of p- galactosidases from A. oryzae ( • ), bovine liver (■) and S. fragilis
(A),Each value represents mean ± S.D. (n=3).
Table I - Effects of metal ions on the activity of p- galactosidases from A oryzae, bovine liver and S.
fragilis
Final Relative activity (%)
None
SDS
KCl
NaCl
CuCU
MnCl.
MgCl^
CaCU
conc.
(mM) A. oryzae Bovine
liver S. fragilis
10 0 10 0 10 0
0 .1 68
70 42
1
41 19 13
1 0
25 13 9
10 0
33
1 09
0 .1
76 92 105
1
85 92 104
1 0
94 92
1 0 110 0
141 103 51
0 .1
75 87 106
1
81 87 104
10
91
8875
10 0
158 74 75
0 .1
90 97 104
1
79 87
10 21 0
44 81 49
10 0
81 71
660 .1
77 104 92
1
80 104
1 0 21 0
73 92 109
10 0
64 85 52
0 .1
64 98 108
1
74 96 105
1 0
75
10 0109
10 0 1 1 2
115
860 .1
79 99 106
1
91 99
1 0 11 0
81
10 0103
10 0
107
1 1 092
Compound Final conc.
(mM)
Relative activity (%) A. oryzae Bovine
liver 5. fragilis
BaCl,
ZnCl-i
CoCL
Fed.
FeCU
0 .1
84 104
1 1 01
82
1 1 1 1 1 01 0
70 114 103
10 0
84 127
880 .1
76 95 107
1
65 97
1 1 11 0
45 70 93
10 0
62
1 1 0125
0 .1 1 1 1
107
1 2 21 10 0 10 2
119
1 0
84
1 0 1 1 2 210 0 1 0 1
99 80
0 .1
79 87 94
1
135
2 113
10 86
7 4
10 0
N.D. N.D. N.D
0 .1
74 85 96
1
144 29 16
1 0
113
6 010 0
44 52 78
N.D. is not determined
상의 층분한반응 시간이 필요함을 나타내는것이다
. 변성제 및 금속이온이 |3-galactosidase2j 활성에 미 치는 영향- Table I
에그결과를 나타내었으며,
금속이 온을첨가하지 않은경우를100% aj
응성이 있는 것으 로 한 경우,
변성제인SDS
를 가하였을때3
중류P- galactosidase
의빈응성이 현저하게 저하하였다.
A
oryzae
유래는 100 m M IC , Na"■이온의 경우 각 각 141%, 158%로 반응성이 증가하였으나,
Cu^-",Ba2",
이온의 경우 반 이 현저하게 감소 하였다.
특히 10 m M C u고■는44%, 10 m M Z n2■은 45%로 감소하였다. 그러나,
M g2우 , C2?\ 는 큰
영향을 미치지 않았다.
한편, Fe^^, Fe
^■"은 Im M
을
가한 경우에는 반응성이 증가하였으나 10 m M 흑은
100
미보에서는오히려 반응성이 저하하였다.
Bovine liver
유래는Mg
그Ca^*, Ba
고■•■이온의경우,
농도가높아질수록 반응성이 약간증가하였으며, Na^
Cu^
■는농도가 높아질수록 반■
융"성이 저하하였다. Fe
이온의 경우는2
가, 3
가 모두1 mM, 10 mM
에서 반응성이 현저하게 감소하는 것을 확인할수 있었다.
한편
, S. fragilis
유래는,
금속이온의 중류에 관계없Vol. 44, No. 2, 2000
윤재경 • 이영재 • 구본웅 • 윤상영 ■ 유%1수 • 김하형
이
0 . 1mM과
1mM에서 반응성이 약간 증가하였으
나, 10 mM 흑은 100 mM에서는 반응성이 감소하였 다. 특허 100 mM 1C과 100 mM 이온에서 각 각 51%, 52%로 현저하게 감소하였으며 10 mM Cu고 ■ •■ 은 49%까지 감소하였다. Co고-미온은 10 mM에서 활성이 약간 증가하였나,
1 0 0피서에서는 번응성이 저 하하였다. Fe이온의 경우
27>,3가 모두 bovine liver 유래와 같이 1 mM, 10 mM에서 거의 반응'성을 나 타내지 못하였다.
결 론
당말단
7}수분해효소 중 당단백질에 대한 ifr옹■조건 이 현재까지 보고되고 있지 않은 A oryzae, bovine liv
05S. fragilis 유래의 P-galactosidase룰 대상으로 반응조건을 연구하였으며, 특허 반응성을 확인하는데 특정 당을 인식하는 lectin을 이용하여 소량의 시료로 확인할 수 있는 유용한 방법임을 확인하게 되었다. 그 결과 반응 최적 pH는 3.5, 3.604. oryzae), 3.5~5.5 (bovine liver, S. fragilis) 반응 적정온도는 37°C, 반 응시간은 24시간이며, 유효농도는 요 oryzae 유래 0.09 mglOO \d, bovine liver 유래 0.29 mg/100 \il, S.
fragilis 유래 0.17 mg/100 p/임을 알 수 있었다. 또
한, 번성제와 이온에 의해서는 번움성이 현저하
게 저하되며 기타 금속이온에 의해서는 빈응성의 향상 흑은 저하가 각각 금속이온 및 그 농도에 따라 다른 결과를 나타내었다.
본 연구에서 실시한 lectin의 당 결합 특이성을 이용 한 p'galactosidase의 반응성 확인은 당의 구조틀 연구 하는데 매우 유용하며 당단백질에 있어서 당이 단백질 의
구조, 기능에 미치는 영향등을 연구하는데 있어 대 단히 중요한 자료를 제공할 것이며, 특히, S. fragilis 유 래의 P*galactosidase에 대한 연구는 그 유•§•성이 거의 보고된 바가 없는 것으로 부터 이 효소에 대한 사용 가 능성이 보다 확대 될 것으로 기대된다. 현재는 asialofetuin중에서 •/V-linked triantennary complex-type 만의 구조를 갖는 당웹타의드률 분리하고 형광@석법을 적용하여 lectin에 의한 결과와 비교 연구하고 있다.
감사의 말씀
이 연구는 1999학년도 중앙대학교 연구기자재 구입
지원프로그램의 도움을 받';>> 수행한 결과의며 이에 감 사드럽니다.
문 헌
1) Kobata, A. : Use of endo- and exoglycosidases for structural studies of glycoconjugates. Anal Biochem., 100, 1 (1979).
2) Rademacher T. W, Parekh R. B., and Dwek R. A. ; Glycobiology. Annu. Rev. Biochem., 57, 785 (1988).
3) Hiraiwa M., and Uda Y. : GMl ganglioside beta- galactosidases from Bovine liver, Jpn /. Exp. M ed, 58, 129 (1988).
4) Stahl, R Rodman, J., Miller, J. and Schlesinger, E : Evidence for receptor-mediated binding of glycopro
teins, glycoconjugates, and lysosomal glycosidase by alveolar macrophages. Proc, Nat. Acad. S ci, 75, 1399 (1978).
5) Martinez-Bilbao, M. and Huber, R. E. : The activa
tion of p-galactosidase (E. coli) by Mg(2+) at lower pH values. Niochem Cell Biol., 74, 295 (1996).
6
) Zeleny R, Altmann E, and Praznik : A capillary electrophoretic study on the specificity of p- galactosidases from Aspergillus oryzae, Escherichia coli Strptococcus pneuminiae, and Canavcdia ensiformis (Jack bean). Anal Biochem. 246, 96 (1997).
7) Kiyohara X Terao T, Shioiri-Nakano K, Osawa T : Purification and properties of a neuraminidase from Streptococcus K 6646. Arch. Biochem. Biophys. 164, 575 (1977).
8
) Tanaka, Y., Kagamiishi, A., Kiuchi, A. and Horiuchi, T. : Purification and Properties of p-galactosidase from Aspergillus oryzae. J. Biochem,, 77, 241 (1975).
9) Harris, R J., Henry, R. J., Blakeney, A. B., and Stone, B. A. : An improved procedure for the methylation analysis of oligosaccharides and polysaccharides, Carbohydr. Res., 127, 59 (1984).
10) Taylor, S. A., Richardson, A. C., Smith, B. V and Price, R. G. : Comparison of several new chromo- genic galactosides as substrates for various p-D- galactosidases. Biochem Biophys Acta, 1163, 54-60 (1993).
11) M. Potier, L. Mameli, M. Bilisle, L. Dallaire and S. B.
Melancon : Fluorometric assay of neuraminidase
with a sodium (4-methylumbelliferyl-a-D-N-acetylne-
(5-galactosidase 의 빈용 조건 203
uraminate) substrate. Anal. Biochem., 94, 287 (1979).
12) Bhavanandan, V E, Yen, A. K, and Carubelli, R. : Neuraminidase assay utilizing sialyl-oligosaccharide substrates with tritium-labeled aglycone. AnaL Biochem,y 69, 385 (1975).
13) K. Ogura, M. Ogura, R. L. Anderson and C. C.
Sweely: Peroxidase-amplified assay of sialidase activity toward ganglioside. Anal Biochem., 200, 52 (1992).
14) T. Nagai and H. Yamada : Assay for sialidase using erythrocytes and peroxidase-labeled peanut lectin.
Chem. Pharm. Bull., 37, 2243 (1989).
15) Mandal, D. K. and Brewer, C. E : Cross-linking activity of 14-kilodalton p-galactoside-specific verte
brate lectin with asialofetuin : Comparison with several galactose-specific plant lectins. Biochemistry, 31, 8465 (1992).
16) Parkkinen, J. and Oksanen, J. : A lectin-Immuno- fluorometric assay using an immobilized Bandeiraea simplicifolia H lectin for the determination of galactosylation variants of glycoproteins. Anal
Biochem., 177, 383 (1989).
17) Nagai, T. and Yamada, H. : Assay ofr sialidase using erythrocytes and peroxidase-labeled peanut lectin.
Chem. Pharm. Bull., 37, 2243 (1989).
18) Onodera, S .: A microplate assay for sialidase activity using plant lectin binding to A^-acetylgalactosamine.
BioL Pharm. Bull., 17, 29 (1994).
19) J. V Baenziger and D. Fiete : Structure of the complex oligosaccharides of fetuin. J. BioL Chem., 254, 289 (1979).
20) S, Takasaki and A. Kobata : Asparagine-linked sugar chains of fetuin : Occurrence of tetrasialyl trian
tennary sugar chains containing the Galpl SGlcNAc sequence. Biochemistry, 25, 5709 (1986).
21) Hampel D. J, Kottgen B, Dudenhausen J. W, Kottgen E : Fetal fibronectin as a marker for an imminent (preterm) delivery. A new technique using the glycoprotein lectin immunosorbent assay. J. Immunol Methods, 224, 31 (1999).
Vol. 44, No. 2, 2000