a S J A S ■ 힐 5 l A f r ~ 배 ™
후 총 학
골손실을 조절하는 활성산소종의 역할 규명
양미혜*-t. 박효정t . 이동석* . 임미정#
숙명여자대학교 약학대학, *경복대학교 자연과학대학생명공학부 (Received March 25, 2009; Revised May 11, 2009: Accepted May 21, 2009)
The Function of Reactive Oxyegn Species in Bone Loss
Mihye Yang*, Hyojung Park, Dong-Seok Lee* and M ijung Yim부 College of Pharmacy, Sookmyung Women's University, Seoul 140-742, Korea
^College of Natural Sciences, Kyungpook National University, Daegu 702-701, Korea
Abstract — We explored the role of reactive oxygen species (ROS) in LPS-induced bone loss. LPS was shown to increase the concentration of ROS in osteoclast precursors. The antioxidant decreased osteoclast formation by LPS. Furthermore, the antioxidant decreased NFATcl expression by LPS, suggesting that ROS mediates NFATcl expression in the regulation of LPS-induced osteoclast formation. Finally, the antioxidant decreased LPS-induced RANKL mRNA expression in osteo
blasts. Taken together, these data indicate that LPS mediates ROS to induce bone loss.
Keywords □ ROS, LPS, osteoclast, osteoblast, bone loss
최근인구고령화에따라노인성 질환이 급증하고 었으며, 이 로인해노인의료비도지난 1 0년간8배이상폭발적으로증가 하고있다. 노화에따른질환중대표적인것이골격계질환으로, 특허 염증성 골손실은류마티스 관절염, 처주염, 골다공증 등을 유발하쉬 삶의질을저하시킨다.
건강한뼈는일생동안뼈가지속적으로형성되고파괴되며, 이 률뼈의 재형성이라한다.^^ 뼈의 재형성은새로운뼈를만드는 조골세포(osteoblast)와오래된뻐를파괴콩]는괴골세포(osteoclast) 에의해평형을유지한다. 조골및파골세포는밀접한관계를맺 고있어, 파골세포의 분화는조골세포에 의해 엄격하게 조절된 다.녀즉, 조골세포는파골세포분화인자인 M-CSF(Macrophage Colony-Stimulating Factor, or CSF-1) 및 RANKL(Receptor Activator of Nuclear Factor-kappaB Ligand, OPGL, ODR or TRANCE)을통해파골세포의 분화를조 절 함 ^써 체내골형성 과골파괴의 동적인평형을유지한다. 2
염^ ^ 골파괴의 병인인자로 lipopolysaccharide(LPS)를돌수 있다.®^ LPS 감염에 의한 골파괴에는 Toll-Like Receptor(TLR) 률매개한자연면역계가중요하다고생각되고있다. TLR는지
끓본 논문에 관한 문의 는 저 자에 게로
^ 화) 02-710-9572 (팩스) 02-710-9871 (E-mail) [email protected]
t양미혜와 박효정은 본 논문에 대해 동일하게 기여하였습니다.
금까지사 림 게 서 11중류, ■미우스에서는 13종류가보고된바있 다 LPS는 TLR family 중 TLR4에 결합하여 IL-6, IL-12,
TNF-a 등의 염증성사의토카인이나 케모카인의 발현을 유도한
다. TLR4의세포내도메인에는 ToML-lR 상동영역(TIR도메인) 이존재하여, TLR4가 LPS와결합하면 TUMI MyD8 8, TIRAI스 TRIP; TRAM, SARM의 adaptor 분자가결합해 세포내에 신호 를 전달한다. 특히 MyD8 8은 TLR4 신호전달계에 있어서 매우 중요한역할을담당하고있다.®
조골및파골세포도표면에 TLR4률 발현하고있다.®> 조골세 포에서 LPS는 TLR4에 걸합해 MyD8 8를 매개해 Ca^VPKC, ERK 의존적으로파골세포분화인자인 RANKL 발현을유도한 다.^® 또한 C0X-2률 활성화해 PGEg 생성을 촉진하고 그의 autocrine/paracrine 작용에의해파골세포분화억제 인자인 OPG 발현을억제함머 알려져 있다.^^^ 이기전에있어서 MyD8 8-A유 래의세포에서 LPS는 RANKL 발현을유도하지 않는 것으로부 터 , TLR4의세포내 adaptor 분자 MyD8 8이 RANKL 생성에중 요함미 밝혀졌다.^^^ 한편 LPS는성숙한파골세포에 직접 작용해 MyD8 8률매개해골흡수기능및생존을촉진하며 , 파골세포견 구세포에 작용해 파골세포로의 분화률촉진함< > 1 알려져 있다/ 3) 그러나이러한일련의보고에도불구하고, LPS가골손실을유 발하는기전은아직 층분히 밝혀지지 않았고, 이는치료제 개발 에필요한타겟분자의 발굴을 어렵게하고 있다. 이에따라 염
179
180 양미혜 • 박효정 • 이동석 ■임미정
증성 골손실은 높은 발병률에도 불구하고 소염진통제 이외에는 뚜렷한 치료제가 없는 실정이다. 이에 본 연구자는 항산화제률 이용해 활 성 산 소 종 (Reactive Oxygen Species, ROS)이 LPS 에 의한 골세포 조절에 미처는 영향을" 조사하였기에 보고하는 버어다.
실험 방법
파골세포의 분화유도
마우스 macrophage cell line인 Raw 264.7 세포 2x10크 cells/
well을 RANKL 50 ng/m/(peprotech) 존재하에서 36시간 배양하 여 파골 견구세포률 생성하였다. 파골세포 형성을 위해 추가적 으 로 300 ng/m/ LPS(Sigma, Germany)와 표기된 양의 N- Acetyl-L-cysteine(NAC, Fluka, Germany)을 처러하여 5일간 배 양하였다. 배양이 끝난 세포는 10% formalin으로 10분간 고정한 후 ethanol-aceton(l: 1)로 1 분 간 재고정 하여 TRAP(tartrate- resistant acid phosphatase) staining을: 했다. 3개 이상의 핵을 가 진 TRAP+ 세포률 다핵 파골세포로 관정했다.
Western blot 분석
파골전구세포를 LPSdng/m/)와 NAC(20mM>을 처리하여 일 정시간 배양 후 , lysate buffer로 용해하고 원심분리 하였다. 단 백질 농도는 BSA(bovine serum albumin)을■ 표준화하여 protein assay kit(Bio-Rad)를 사용하여 정량하였다. 20 나요의 단백질을 SDS-PAGE(poly acrylamide gel electrophoresis)로 번성 분리하 고, 이룰 80v에서 1시간 45분 처리하여 PVDF membrane에 이 견 시 켰 다. 이룰 5% skim milk가 함유된 PBST 용 액 으 로 blocking하고, 1차 항체로서 anti-NFATcl(l: 1000, Santa cruiz), C0X-2(1:2000, R&D systems), IkB(1 :2000, Cell signaling), 또는 p-actin(l; 4000, Sigma) 항체와 각각 반응시켰다. PBST로 5희 세정하31 HRPCHorseradish peroxidase)가 결합된 2차 항체 와 반응시킨 무 ECL Advance(Amersham. CO.)로 발색시켜 분 석하였다.
마우스 조골세포의 초기 배양
생후 0~1일의 ICR mouse로부터 두개골 피부를 벗긴 후 두 개골을 적출하였다. 이때 후두골 근육 부착부분은 제거하였다.
부착된 근육, 혈구 등을 제거한 후 a-MEM으로 가볍게 세척하 였다. 0.1 % collagenase와 0.2% dispase 흔합효소 용액에 넣어 37T에서 5분간 진탕시킨 후 상등액을 버리고 새로운 효소 용액 을 가하였다. 37°C에서 약 10분간 진탕하^ 상등액을 모으는 조 작을 4희 반복하였다. 원심 분리한 후 10% FBS 함유 a-MEM
^ 약 3~5xl(y* 세포/100 mm plastic dish가 되도록 접종하였 다. 5% C02, 37X 에서 3-4일간 배양한 세포률 이후 조골세포 로 실험에 용하였다.
RNA 분리 및 RT-PCR 분석
Total RNA 추출은 easy-Blue(iNtRON Bio technology co., Ltd, Korea)를 이 용 하 였 다 . cDNA는 l|ig의 total RNA를 oligodT primer, 10 mM dNTR 1 unit RNase inhibitor 그리고 4 unit Script reverse transcriptase(Fermentas, Life science)로 42°C에서 sag: 처리하여 합성한 후 , 70°C에서 10분 가열함으로 써 반응을 중지 시 켰 다 . Polymerase chain reaction(PCR)은 RANKL에 특이적인 서 열 (forward: 5'-ccagccatttgcacacctc-3', reverse: 5'-agcagggaagggttggaca-3')을 사용하여 94°C 3분간 처 리한 후 , 94°C 30초 , 58°C 45초 72°C 1분의 과정을 32희 반복 하였다. PCR 결과물은 견기영동으로 확인하였다.
ROS 생성농 측정 (DCF 법)
ROS 생성능I: 측정하기 위해 DCFH-DA(2',7"dichlorofluorescin- diacetate)률 사용하였다. Raw 264.7 세포률 RANKL이 포함된 배지에서 36시간 동안 배양하고, 0.5% FBS가 포함된 배지에서 12시간 동안 배양하였다. 0.05 mM DCFH-DA를 30분 동안 처 리하고, PBS로 washing한 후 시간대별로 LPS(300 ng/m/)률 처 리 하 였다. 생성된 DCF 형광의 번화는 excitation, 488 nm 및 emission, 515—540 nm에서 Fluorescence Microplate Readei로 측정하였다.
통계처리
실험결과는 평균 + 표준편차로 표기하였고, Student's t-test로 분석하여 P 값이 0.05 미만일 때 통계적으로 유의하다고 판단하 였다.
실험 결과 및 고찰
LPS에 의한 파골전구세포내 활성산소종의 증가
먼저 LPS에 의한 파골세포 분화에 관여하는 활성산소종의 역 할을 규명하기 위해, 파골전구세포률 LPS로 처리하였을 때 세포 내 활성산소중의 번화를 시간대별로 조사하였다. 파골전구세포 로는 마우스 macrophage cell line인 Raw 264.7 세포률 파골세 포 될수 분화인자인 RANKL로 36시간 처리하식 형성된 세포를 사용하였다. 파골전구세포는 LPS로 처리하여 5일간 배양하면 다 핵의 성숙 파골세포로 분화한다(data not shown). 파골견구세포 내 활성산소중의 농도는 300 ng/m/의 LPS 처리 후 10분부터 유 의적으로 증가하였으며 2시간에서 최고치룰 나타냈다(Fig. 1). 그 후 점차적으로 감소하였으나, 처리 후 6시간에서도 비교적 높은 세포내 활성산소중 농도률 유지하였다. 이상의 결과로 LPS가 파 골견구세포내에서 활성산소종 농도률 증가시킴을 알 수 있으며, 이는 활성산소종이 LPS에 의한 파글세포 분화에 관여할 가눙성 을 보여주는 것이라 할 수 있다.
2 - The involvement of ROS in LPS-induced osteoclast for
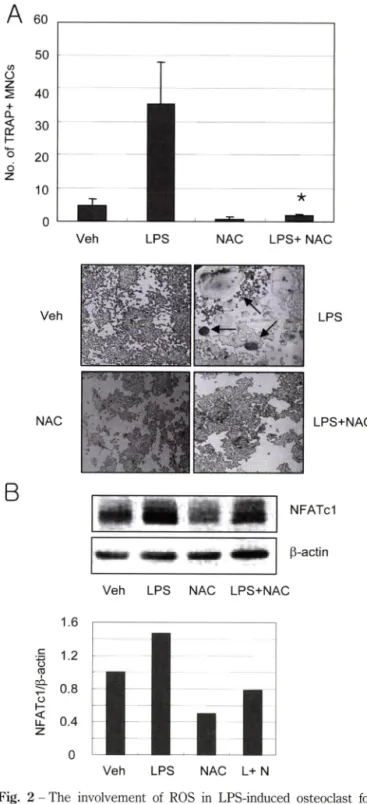
mation. A, Osteoclast precursor cells were further cultured with 300 ng/ml LPS for 5 days in the absence or presence of 10 mM NAC pretreatment. Cells were then fixed and stained for TRAE TRAP-positive (+) multinucleated cells (MNCs) were counted. The experiments were performed 3 times, and the reproducibility was confirmed. Values are the mean±SD of triplicate cultures in a representative experiment. Veh: vehicle, *:^<0.05, significantly different from LPS. B, Osteoclast precursor cells were cultured with 300 ng/ml LPS for 2 days in the absence or presence of 20 mM NAC pretreatment. The expression of NFATcl was determined by western blotting assay. Western blot images are representative of 3 experiments with similar results.
50
40 30
20 10
Veh
Veh LPS NAC LPS+ NAC
LPS
NAC LPS+NAC
n i l m u NFATcl
P-actin Veh LPS NAC LPS+NAC
A
600 10 20 30min 1 2 4 6hr
Fig. 1 - Effects of LPS on the concentration of ROS in osteoclast precursor cells. Raw 264.7 cells were cultured with 50 ng/
ml RANKL for 36 hrs to produce osteoclast precursor cells.
After starvation with 0.5% FBS for 12 hrs, cells were treated with 300 ng/ml LPS for indicated times. The concentration of ROS were measured as described in Materials and methods. The experiments were performed 3 times, and the reproducibility was confirmed. Values are the mean±SD of triplicate cultures in a representative experiment. *: /)<0.05, significantly different from 0 hr.
항산화제에 의한 LPS 유도성 파골세포 분화의 억제
LPS 유도성 파골세포 분화에 있어서 활성산소종의 기능을 규 명하기 위해, LPS와 항산화제률 함께 처리하였을 때 파골세포로 의 분화 번화률 주적하였다. 항산화제로는 N-Acetyl-L-cysteine (NAC)을 사용하였다. 파골전구세포는 300n@/m/의 LPS 처리로 TRAP+ 다핵 파골세포로 분화되었으며, 이는 10 mM NAC 처 리로 유의성 있게 억제되었다(Fig. 2A). 이로써 LPS는 파골전구 세포내 활성산소중을 데개하여 파골세포로의 분화를 촉진함이 밝혀졌다.
파골세포 분화에 관여하는 많은 유전자가 밝혀졌으며, 그 중 에서 특히 Nuclear Factor of Activated T cells(NFAT)cl은 파 골세포의 분화률 조절하는 중요한 견사인자로 알려져 있다.14) 항 산화제의 처리가 LPS에 의한 NFATcl의 발현을 번화시키는지 여부를 항 NFATcl 항체를 이용한 western blotting assay로 조 사하였다. 파골전구세포를 LPS로 48시간 처리하였을 때 NFATcl 의 발현은 증가하였으며, 이는 NAC 처리로 감소하는 것으로 나 타났다(Fig. 2B), 이는 항산화제가 LPS에 의한 파골세포 분화률 억제하는 것과 일치하는 결과로, 활성산소종은 NFATcl 발현을 조 절 함 써 LPS에 의한 파골세포 분화를 매개하는 것으로 추 정되 었다.
LPS에 의한 파골세포 분화에 cydoxygenease-2(COX-2)페와 nuclear factor kappa B(NF-kB)페가 관여함이 보고된 바 있다.
따라서 LPS에 의해 유발된 활성산소종이 C0X-2와 NF-kB률 조 절하는지의 여부를 항산화제 처리에 의한 western blotting assay 로 조사하였다. LPS는 파골전구세포에서 C0X-2 발현을 증가시
Vol. 53, No. 4, 2009
골손실을 조절하는 휠 성 산 석 역할 규명 181
N NAC Vet P
0
SONI/M+dvy.uo
O N
B
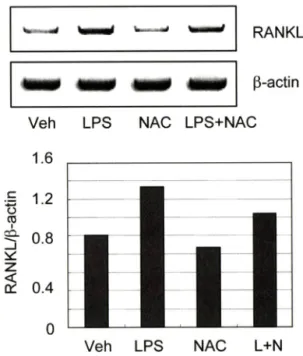
Fig. 4 - The involvement of ROS in LPS-induced RANKL expres
sion. Mouse calvarial osteoblasts were treated with 300 ng/
mi LPS in the absence or presence of 20 mM NAC pretreatment for 3 hrs. The mRNA expression of RANKL were determined by RT-PCR. Band images are representative of 3 experiments with similar results.
항산화제에 의한 LPS 유도성 RANKL 발현의 억제
LPS는 파골전구세포에 직접 작용하여 파골세포 분화률 촉진 할 뿐 아니라, 조골세포내 RANKL 발현을 유도함으로써 파골세 포 분화를 간접적으로도 촉진하는 것이 알려져 있다.^® 따라서 LPS에 의한 조골세포내 RANKL 발현이 활성산소종을 매개하는 지 규명하고자 하였다. 이를 위해 항산화제 처리에 의한 조골세 포내 RANKL 발현 번화를 RT-PCR로 조사하였다. 조골세포로 는 미우스의 두개관에서 분리한 초기배양 조골세포를 사용하였 다. LPS 처리는 조골세포내 RANKL 발현을 촉진하였으며, 그 발현 촉진 효과는 항산화제 처리로 다소 감소하는 것^ 나타 났다(Fig. 4). 따라서 활성산소중은 LPS의 파골전구세포에 대한 직접적인 작•§뿐 아니라 조골세포에 대한 간접적인 작용도 매개 하는 것으로 밝혀졌다. 본 연구률 통해 LPS에 의한 파골세포 분 화 촉진을 매개하는 활성산소종의 기눙이 처음으로 밝혀졌으며, 이는 체내 LPS에 의한 염증성 골손실의 새로운 발병기견을 밝 히는 것과 동시에, 항산화제의 염증성 골손실 치료제로서의 가 능성을 제시해주는 것이라 할 수 있다.
결 론
LPS는 파골전구세포내 활성산소중의 농도률 증가시켰으며, 활 성산소중은 LPS에 의한 파골세포 분화 촉진을 매개함이 밝혀졌 다. 활성산소종은 파골세포 분화에 필수적인 견사인자 NFATcl
Veh LPS NAC L+N
Fig. 3 - The involvement of ROS in LPS-activated signaling path
ways. Osteoclast precursor cells were cultured with 300 ng/m/ LPS for 24 hrs (COX-2) or 30 min (IkB) in the absence or presence of 20 mM NAC pretreatment. The expression of COX-2 (A) or IkB (B) was determined by- western blotting assay. Western blot images are re
presentative of 3 experiments with similar results.
켰으나, 항산화제 처리는 런 번화를 보이지 않았다(Fig. 3A).
또한 LPS는 IkB 분해를 촉진하여 NF-kB를 활성화시키는 것으 로 나타났으나, 항산화제는 이에 대해 아무 영향을 머치지 않았 다(Fig. 3B). 이상의 결과는 LPS에 의해 증가된 활성산소중이 C0X-2 또는 NF-kB의 신호전달경로와는 무관하게 파골세포 분 화를 조절함을 시사한다. 향후 추가적인 실험을 통해 활성산소 중이 매개하는 LPS 유도성 파골세포 분화 기견이 보다 명확하 게 규명되기를 기대한다.
양미혜 • 박효정 ■이동석 • 임머정
A
설찰률들텔!헬!^^ 설블설igil텔제률/ / 출 W P
J 톰 흡 헬 월 ^ ^ ^ ^ ^ 실
Veh LPS NAC LPS+NAC COX-2
P-actin
'아-*"^ RANKL
m m m m t m am m m
P-actin Veh1.6
LPS NAC LPS+NAC
NAC
벨황 P-actin
Veh LPS NAC LPS+NAC
2
0.80.4
c!PB-drwNvd
0
4
d>
u!}oe- 란q>t|
iKi
2840
1 1 u!}oe-tl/2-xoo
B
골손실을 조절히는 활성산소종적 역할 규명
을 조절함으로써 LPS의 작용을 매개하는 한편, C0X-2와 NF-kB 의 조절과는 무관한 것으로 밝혀졌다. 또한 활성산소중은 LPS에 의한 조골세포내 RANKL 발현도 매개하는 것으로 나타나, 파골 견구 및 조골세포에 대한 LPS의 작용에 모두 관여하는 것으로 추정되었다.
감사의 말씀
본 연구는 보건장학회의 지원으로 수행되었으므로 이에 감사 드럽니다.
참고문헌
1) Takahashi, N., Akatsu, T, Udagawa, N., Sasaki, T, Yamaguchi, A., Moseley, J. M., Martin, T. J. and Suda, T.: Osteoblastic cells are involved in osteoclast formation. Endocrinology 123, 2600 (1988).
2) Suda, X, Takahashi, N., Udagawa, N., Jimi, E., Gillespie, M. T.
and Martin, T. J. : Modulation of osteoclast differentiation and function by the new members of the tumor necrosis factor receptor and ligand families. Endocr. Rev. 20, 345 (1999).
3) Wong, B. R., Rho, J., Arron, J., Robinson, E., Orlinick, J., Chao, M., Kalachikov, S., Cayani, E., Bartlett, E S. 3rd, Frankel, W. N., Lee, S. Y. and Choi, Y.: TRANCE is a novel ligand of the tumor necrosis factor receptor family that activates c-Jun N- terminal kinase in T cells. /. Biol Chem. 272, 25190 (1997).
4) Yasuda, H., Shima, N., Nakagawa, N., Yamaguchi, K., Kinosaki, M., Mochizuki, S., Tomoyasu, A., Yano, K., Goto, M., Murakami, A., Tsuda, E., Morinaga, T, Higashio, K., Udagawa, N., Takahashi, N. and Suda, T. : Osteoclast differentiation factor is a ligand for osteoprotegerin/osteoclastogenesis- inhibitory factor and is identical to TRANCE/RANKL. Proc.
Natl Acad. Sci USA 95, 3597 (1998).
5) Lacey, D. L., Timms, E., Tan, H. L., Kelley, M. J., Dunstan, C. R., Burgess, T, Elliott, R., Colombero, A., Elliott, G., Scully, S., Hsu, H., Sullivan, J., Hawkins, N., Davy, E., Capparelli, C., Eli, A., Qian, Y. X., Kaufman, S., Sarosi, L, Shalhoub, V, Senaldi, G., Guo, J., Delaney, J, and Boyle, W. J. : Osteoprotegerin ligand is a cytokine that regulates osteoclast differentiation and activation. Cell 93, 165 (1998).
6) Nail; S. R, Meghji, S., Wilson, M,, Reddi, K,, White, R
and Henderson, B. : Bacterially induced bone destruction:
mechanisms and misconceptions. Infect Immun. 64, 2371 (1996).
7) Erard, E and Ryffel, B. : Toll like receptor - potential drug targets in infectious disease. Infect Disord. Drug. Targets. 8, 221 (2008).
8) Pilsson-McDermott, E. M. and O'Neill, L. A. : Signal transduction by the lipopolysaccharide receptor, Toll-like receptor-4. Immunology 113, 153 (2004).
9) Itoh, K,, Udagawa, N., Kobayashi, K., Suda, K., Li, X., Takami, M., Okahashi, N., Nishihara, T. and Takahashi, N. : Lipopolysaccharide promotes the survival of osteoclasts via Toll-like receptor 4, but cytokine production of osteoclasts in response to lipopolysaccharide is different from that of macrophages. / Immunol 170, 3688 (2003).
10) Kikuchi, T, Matsuguchi, T, Tsuboi, N., Mitani, A,, Tanaka, S., Matsuoka, M., Yamamoto, G., Hishikawa, T, Noguchi, T and Yoshikai, Y. : Gene expression of osteoclast differentiation factor is induced by lipopolysaccharide in mouse osteoblasts via Toll-like receptors. J. Immunol 166, 3574 (2001).
11) Coon, D., Gulati, A., Cowan, C. and He, J. : The role of cyclooxygenase-2 (COX-2) in inflammatory bone resorption./.
Endod. 33, 432 (2007).
12) Sato, N., Takahashi, N., Suda, K., Nakamura, M., Yamaki, M., Ninomiya, T., Kobayashi, Y., Takada, H., Shibata, K., Yamamoto, M., Takeda, K., Akira, S,, Noguchi, T. and Udagawa, N, : MyD88 but not TRIF is essential for osteoclastogenesis induced by lipopolysaccharide, diacyl lipopeptide, and IL- lalpha. J. Exp Med. 200, 601 (2004).
13) Suda, K., Woo, J. T, Takami, M., Sexton, R M. and Nagai, K. : Lipopolysaccharide supports survival and fusion of preosteoclasts independent of TNF-alpha, IL-1, and RANKL./.
Cell Physiol 190, 101 (2002).
14) Takayanagi, H. : The role of NFAT in osteoclast formation.
Ann. N. Y Acad. Sci 1116, 227 (2007).
15) Kaneko, H., Mehrotra, M., Alander, C., Lemer, U., Pilbeam, C. and Raisz, L. : Effects of prostaglandin E2 and lipopolysaccharide on osteoclastogenesis in RAW 264.7 cells.
Prostaglandins Leukot Essent Fatty Acids. 77, 181 (2007).
16) Zou, W and Bar-Shavit, Z. : Dual modulation of osteoclast differentiation by lipopolysaccharide. J. Bone Miner. Res, 17, 1211 (2002).
Vol. 53, No. 4,2009