유
유기 기물 물원 원 항 항온 온배 배양 양 온 온도 도가 가 토 토양 양미 미생 생물 물체 체량 량과 과 효
효소 소활 활성 성 및 및 P PLLFFA A함 함량 량에 에 미 미치 치는 는 영 영향 향
Effect of Temperature on Soil Microbial Biomass, Enzyme Activities, and PLFA Content During Incubation Period of Soil Treated
with Organic Materials
좌재호*·문두경·전승종·김천환·최경산·현해남1·강위금2
Jae-Ho Joa
*, Doo-Gyung Moon, Seung-Joung Chun, Chun-Hwan Kim, Kyung-San Choi, Hae-Nam Hyun
1, and Ui-Gum Kang
2국립원예특작과학원, 1제주대학교, 2국립식량과학원
National Institute of Horticultural & Herbal Science, RDA
1
Major of Plant Resources and Environment, Jeju National University
2
National Institute of Crop Science, RDA
This study was carried out to evaluate the effect of temperature on soil microbial biomass, enzyme activities, and PLFA content in the volcanic(VAS) and the non-volcanic ash soil(NVAS). The soils were treated with organic materials such as organic fertilizer pelleted(OFPL), organic fertilizer powdered(OFPD), pig manure compost(PMC), and food waste compost(FWC). Two grams of organic materials were well mixed with 30g of dried volcanic and non-volcanic ash soil(< 2 mm) with 50% of soil moisture content. And the soils were incubated at 10, 20, 30℃ ℃ in incubator. Soils were analysed on the incubation times as followed; soil pH, total nitrogen, organic matter(at 75, 150, 270 days), microbial biomass C and PLFA (at 75, 270 days), microbial biomass N and soil enzyme(at 150, 270 days). pH values of soils treated with PMC and FWC had no changes on soil type, and incubation temperature. However, the pH was increased with temperature in the soils treated with OFPL. The changes in NVAS was higher than in VAS. Soil microbial biomass C content were high in the condition of high temperature and organic fertilizers treatment in VAS. But the contents were gradually decreased with incubation period in both NVAS and VAS. Soil microbial biomass N was high in NVAS treated with organic fertilizers and in VBS treated with PMC and FWC. PLFA content was higher in NVBS than in VBS at 75 days but showed high in VBS at 270 days. Urease activity of NVBS treated with OFPL showed 10℃ ℃ (75.0)> 20℃ ℃ (16.3)>30℃ ℃ (4.6 ug NH
4-N g
-12h
-1) at 150 days. It were decreased gradually high temperature and time passes. And it showed high at 10℃ ℃ in VBS. β β -glucosidase activity was higher in NVBS than in VBS. Correlation coefficient of between soil microbial biomass C and microbial activity indicators showed that PLFA was high significantly at r
2=0.91 in NVBS and β β -glucosidase was r
2=0.83 in VBS. Soil microbial activities showed differences in the relative sensitivities of soil type and soil temperature.
Key words: Volcanic ash soil, PLFA, Microbial biomass C, Soil enzyme, Organic matrials
서 언
최근 친환경농업이 확산되면서 유기질비료와 가축분 뇨를 원료로 한 부산물퇴비의 사용이 증가하고 있다.
토양에 유기물이 시용되면 미생물의 작용에 의해 유 기탄소 분해와 질소 무기화가 일어나며, 유기질비료는
속효성 화학비료와 달리 미생물 분해활동에 의하여 시비효과가 느리게 나타난다. 분해된 양분은 토양내에 서 미생물과 작물의 영양원으로 이용되고 미생물의 활성을 증가시킨다. 토양유기물의 분해와 무기화과정 은 미생물의 개체수, 유기물 분해효소활성, 미생물의 양분요구도 등 생물적 요인과 토양특성, pH, 온도, 수 분함량, 유기물의 화학적 조성 등 비생물적 요인이 복 합적으로 상호작용하여 일어난다(Alvarez et al., 1995;
접 수 : 2009. 10. 20 수 리 : 2009. 11. 19
*연락저자 : Phone: +82647412581, E-mail: [email protected]
Manzoni and Porpotato, 2007).
토양유형과 경지이용 형태는 토양미생물의 기능에 크게 영향을 주며, 화산회토양은 유기물과 allophane 이 복합체를 형성하여 알루미늄 독성에 의하여 토양 미생물의 활성이 낮고(Anderson, 1979; Henmi et al., 1976; Nanzyo, 2002; Satti et al., 2007; Song, 1990;
Tokashiki and Wada, 1975; Ugolini and Dahlgren, 2002), 비화산회토양 보다 질소 무기화율(Deenik, 2006; Saito, 1990)과 유기탄소 무기화율(Takenaka and Hayano, 1999)이 낮으며 K2SO4 침출 미생물체량 C는 화산회토양에서 비화산회토양보다 높다고 하였다 (Nishiyama et al., 2001). 토양에 유기물이 시용되면 토양산도를 교정하고 pH 완충력을 높이지만 일시에 다량의 유기물시용은 분해과정에서 작물뿌리에 피해 를 주고 일시적으로 토양 pH를 낮추기도 하며 토양 중에서 작물과 미생물간에 질소 양분경쟁을 일으키기 도 한다. 토양 pH는 유기물의 분해과정에서 질소의 휘산, 미생물 개체수의 변화(Kelly and Henderson, 1978), 토양 질소와 탄소 순환 및 미생물군집 기능에 영향을 준다(Kemmitt et al., 2006; Nanzyo, 2002). 미 생물 활성을 평가하는 지표로 미생물체량, 토양효소, 인지질지방산이 있는데, Hu and Cao(2007)는 미생물 체량과 토양효소 활성은 토양비옥도를 반영하여 토양 질의 중요한 생물지표가 된다고 하였다. 유기물의 분 해와 미생물 활성은 토양 온도와 수분함량 등에 영향 을 받으며(Anderson, 1991; Kirschbaum, 1995; Koch et al., 2007; Nobili, 2006; Paul and Clark, 1989), 온 도가 높을수록 토양미생물체량 C와 총유기탄소간에는 상관관계가 높다고 하였다(Cerhanova et al., 2006).
토양미생물의 몸체를 구성하는 미생물체량 C보다는 작지만 미생물체량 N은 미생물의 개체수와 밀접한 관계를 가지고 있어 미생물의 밀도를 측정하는데 유 용한 수단이 되며 유기물이 투입되면 질소 무기화와 미생물체량 N이 증가한다(Goyal et al., 2000). 미생물 활성을 평가하는 방법으로 선택배지를 이용하여 배양 이 되는 미생물의 종류와 개체수를 파악하였으나 시 료 채취, 배양조건, 배지 종류, 양분요구도 등에 영향 을 받아 변이가 크고 정확도가 떨어지는 단점이 있다.
인지질 지방산은 살아있는 미생물에만 존재하기 때문 에 배양이 어려운 미생물까지도 정량할 수 있어 미생 물의 활성을 평가하는데 유용하고 (Baath and Anderson, 2003; Green and Scow, 2008), pH가 증가 하면 PLFA함량도 증가한다고 하였다(Cookson et al.
2007). 토양에 유기물을 첨가하면 미생물체량과 Urease 활성이 증가하며(Bremner and Mulvaney, 1978; Dinesh et al., 1998; Klose and Tabatabai, 1999,2000; Perucci 1990; Roscoe et al. 2000), 토양효
소활성은 자연적, 인위적 교란에 민감하게 빨리 반응 이 나타난다(Dick, 1992).
β-glucosidase와 Urease가 토양생물학적 활성의 지표 (Hu and Cao, 2007; Timothy and Dick, 2004)로 사용 될 때 토양 온도와 수분함량은 중요한 요인이 된다 (Alvarez et al., 1995; Debosz et al., 1999; Sardans et al., 2008). 유기물의 분해특성과 조성은 유기물의 분해 와 미생물활성에 영향을 미친다. 토양별 유기물원의 항온조건배양시 분해특성과 이에 따른 미생물의 활성 비교는 유기물 시용량과 시용시기를 정하고 토양을 관리하는데 있어 중요한 요소이며, 합리적인 유기물시 용 및 시비관리를 위하여 필요하다. 유기물시용이 미 생물활성(Dinesh et al.,1998)과 토양효소활성(Debosz et al., 1999; Hu and Cao, 2007; Klose and Tabatabai, 2000; Marschner et al., 2003)에 미치는 영향과 항온 조건에 따른 유기물의 탄소와 질소 무기화 특성 (Agehara and Warncke, 2005; Cordovil et al., 2005;
Kemmitt et al., 2008; Stanford and Smith, 1972), 질 소무기화에 따른 양분의 공급력(Sharifi et al., 2008), 미생물체량에 대한 연구보고는 많이 되어왔다. 화산회 토양의 유기재배 감귤원은 관행재배보다 PLFA함량 과 미생물체량이 높았으며(Joa et al., 2008), 화학비료 +퇴비구가 무비료구 보다 Dehydrogenase와 β- glucosidase 활성이 높았다고(Joa et al., 2009) 보고한 바 있으나 제주지역 화산회토와 비화산회토양에서 유 기물시용에 따른 미생물의 활성과 변화에 대한 연구 는 많지 않다. 본 연구는 화산회토양과 비화산회토양 에서 유기물원을 항온 배양했을 때 온도가 토양미생 물체량과 효소활성 및 PLFA함량에 미치는 영향을 토양특성별로 평가하고자 수행하였다.
재료 및 방법
흑색화산회토와 암갈색비화산회토양에 유기물원으 로 제주도내 감귤재배농가에서 가장 많이 시용되고 있는 혼합유기질비료 2종류(입상과 분상), 돈분퇴비, 음식물퇴비를 토양 30 g에 2 g을 잘 혼합 후 토양수 분함량을 50%로 조절하여 밀폐된 1.2 L의 플라스틱 용기에 넣어, 10, 20, 30℃에서 항온배양을 하면서 토 양을 채취하여 pH, 토양질소, 유기물 함량(75, 150, 270일), Microbial biomass C(75, 270일), Microbial biomass N, 토양효소활성(150, 270일), 인지질지방산 함량(75, 270일) 변화를 2반복으로 분석하였다. 시험 토양은 초기 45일 동안은 7일 간격으로 잘 혼합하였 으며 이후 시료 채취시에 잘 혼합한 후 시료채취를 하였다. 시험 토양과 유기물원의 화학성분은 농촌진 흥청 표준분석법에 준하여 시료를 전처리 후 분석하 여 평균값을 산출하였다(RDA, 1988). 비화산회토양은
다년간 경작한 미사질식토의 동홍통으로 pH 5.8, 유기 물함량 26.9 g kg-1, 토양질소 0.1%, 유효인산함량 9.4 mg kg-1, 치환성 칼슘함량 5.9 cmolc kg-1, 치환성 알 루미늄함량이 150.5 mg kg-1였다(Table 1.). 화산회토 양은 비경작토양인 미사질식양토의 송당통을 이용하 였으며, 이화학성분은 pH 5.2, 유기물함량 145.2g kg-1, 토양질소 0.6%, 유효인산함량 4.0 mg kg-1, 치환성 칼 슘함량 1.0 cmolc kg-1, 치환성 알루미늄함량이 968.3 mg kg-1으로 토양질소 함량은 비화산회토양보다 높았 고 칼슘함량은 적었다. 분상유기질비료(O.F.-Powder) 는 골분, 어분, 탈지강, 유박, 입상유기질비료(O.F.- Pellet)는 유박, 쌀겨, 골분, 어분, 팜박, 돈분퇴비는 돈 분과 톱밥(3:7), 음식물퇴비는 음식물쓰레기와 톱밥 (5:5)이 혼합된 것을 이용하였다(Table 2.). 입상과 분상 유기질비료는 질소함량이 각각 3.80, 4.75%로 음 식물퇴비(2.06)와 돈분퇴비(2.52%)보다 많았다. 인산 함량은 입상유기질비료(3.92), 분상유기질비료(2.95), 돈분퇴비(3.69), 음식물퇴비(2.45%)였으며, 칼륨함량 은 돈분퇴비가 3.13%로 가장 높았다. 칼슘함량은 음 식물퇴비(5.65)>입상유기질비료 (3.13)>돈분퇴비 (2.67)>분상유기질비료(2.25%)순이었다. 돈분퇴비의 Zn과 Cu함량은 각각 478.2 , 332.1 ppm으로 가장 많 았으며, C/N비는 41.1로 입상 유기질비료 23.5, 분상 유기질 비료 19.0 보다 높았다.
미생물체량 C Vance et al.(1987)의 방법을 이용 하여 2 mm체를 통과한 토양시료를 클로로프롬으로 훈증배양한 후 Microbial biomass C를 추출하여 추출 시료액 8 mL를 250 mL 둥근바닥 플라스크에 취한
후, 2 mL의 66.7 mM K2Cr2O7 용액, 황산과 인산의 혼합액(2:1) 15 mL을 첨가하여 혼합 후 환류냉각관 으로 분해한 다음, 적정하여 Microbial biomass C 함 량을 구하였다.
미생물체량 N Microbial biomass C 추출액을 이용 하여 가용성 유기태질소의 1/2을 점유하는 아미노산태 질소량을 구하는 Ninhydrin 발색법을 이용하여 분석하 였다(Amato and Ladd, 1988). 추출액 2 mL에 ninhydran reagent 1 mL를 가한 후 끊는 수조안에서 25 분간 아미노산태질소(+암모니아태 질소)를 발색시 킨 후 냉각하여, 95% ethanol과 증류수(1:1)혼합액 4.5 mL을 가하여 혼합 후 UV-Visible Spectrophotometer (Cary 100, Varian)를 이용 570 nm에서 비색법으로 측정 후 계산하였다.
토양효소활성 Urease는 토양 5 g을 각각 50 mL 시험관에 취한 후 20 mL의 Borate buffer(0.1 M, pH 10)을 넣고, 처리구에 2.5 mL의 720 mM Urea용액을 가하고 밀봉을 하여 37℃에서 2시간 동안 배양 후 2 M KCl용액 30 mL를 가하고 30분간 진탕 후 여과하 여 Sodium salicylate/Sodium hydroxide 혼합액 5 mL 와 39.1 mM Sodium dichloroisocyanide 2 mL를 가한 후 UV-Visible Spectrophotometer(Cary 100, Varian) 를 이용하여 660 nm에서 측정하였다. 배양 현탁액을 여과하기 직전에 대조구에 2.5 mL의 720 mM Urea용 액과 2 M KCl용액 30 mL를 가하고 30분간 진탕 후 여과하여 Sodium salicylate/Sodium hydroxide 혼합액 5 mL와 39.1 mM Sodium dichloroisocyanide 2 mL를
C/N Cu
Zn CaO
K2O P2O5
--- % --- --- ppm --- N
3.80 4.75 2.52 2.06 OFPL"
OFPD PMC FWC
"
OFPL : Organic fertilizer pelleted, OFPD : Organic fertilizer powdered, PMC : Pig manure compost, FWC : Food waste compost.
3.92 2.95 3.69 2.45
1.27 1.06 3.13 0.61
3.13 2.25 2.67 5.65
62.3 63.1 478.2 85.5
21.3 25.9 332.1 42.1
23.5 19.0 31.9 41.1
Table 2. Chemical properties of organic materials.
VAS"
NVAS
5.2 145.2 0.6 4.0 0.4 1.0 0.5 0.15 968.3
5.8 26.9 0.1 9.4 1.7 5.9 2.4 0.15 150.5
Exch. Al Na
Mg Ca
Exch. cations Av.P2O5 K
T-N O.M.
pH
mg kg-1 --- cmolckg-1---
mg kg-1 g kg-1 %
1:5
"
VAS : Volcanic ash soil, NVAS : Non-volcanic ash soil.
Table 1. Chemical properties of experimented soil.
가한 후 측정하여 처리구와 대조구의 차로 효소활성 을 측정하였다. β-glucosidase는 토양 1 g을 50 mL 삼 각플라스크에 취하고 4 mL의 MUB 완충용액(12.1 g trishydroxymethyl aminomethane, 11.6 g maleic acid, 14.0 g citric acid, 9.3 g H3BO3, 488 mL 1N NaOH, pH 6.0), 1 mL의 0.025 M p-nitrophenyl-β-D- glucoside용액을 가한 후 플라스크에 마개를 하고 37
℃에서 1시간 동안 배양 후 1 mL의 0.5 M CaCl2용액 과 4 mL의 0.1 M THAM완 충 용 액 (12.2 g trishydroxymethylaminomethane, 0.5M NaOH, pH 12) 을 첨 가 한 다 음 에 여 과 하 여 UV-Visible Spectrophotometer (Cary 100, Varian)를 이용 400 nm에서 흡광도를 측정하였다.
인지질 지방산 분석 2 mm체로 선별한 토양을 동 결건조 후 인지질 지방산(Phospholipid fatty acid, PLFA)함량은 Bligh/Dyer first-phase extraction(Bligh and Dyer, 1959)방법을 이용하여 미생물 세포막 지방 산을 추출한 다음 Silicic acid column chromatography 를 이용하여 중성지질과 당지질을 순차적으로 제거한 후 인지질을 분획하였다. 분획된 시료는 methylation 시킨 후 MIDI Sherlock Microbial Identification System(MIDI Inc., Newark, DE)으로 인지질 지방산 을 정량하였다.
통계분석 SAS프로그램을 이용하여 온도별로 미 생물체량 C와 N, 토양효소활성, 인지질 지방산함량을 비교하기 위하여 던칸의 다중범위 검정(DMRT)으로 유의수준 5%에서 통계분석을 하였다.
결과 및 고찰
온도별 토양화학성의 변화 암갈색 비화산회토양 에서 온도별 유기물원의 항온배양에 따른 시험토양의 화학성분 변화는 Table 3.에 나타냈다. 돈분과 음식물 퇴비는 온도에 상관없이 토양 pH 변화가 크지 않았 다. 분상유기질비료는 온도가 높아짐에 따라 pH가 낮
아졌으나 20℃와 30℃는 비슷한 경향이었다. 입상유기 질비료는 시간이 경과하면서 pH가 낮아졌고 변화폭 이 컸으며, 고온으로 갈수록 pH가 낮아졌다. 토양중 C/N비는 토양의 탄소와 질소 비율을 나타내는 것으 로 미생물 몸체구성분인 미생물체량과 관계가 깊으 며, 유기물 분해정도를 측정하는 지표로서 토양 유기 물이 분해가 되면 C/N비는 낮아진다(Friedel et al., 2006; Lagomarsino et al., 2009). 돈분퇴비는 10℃, 75일 에 179.5로 가장 높았으며 시간이 경과 할수록 낮아졌 다. 20℃는 150일에 높았다가 270일에 낮아졌으며, 30℃
는 75일에 높았다가 점차 낮아졌다. 음식물퇴비는 10℃
에서 돈분퇴비와 같은 경향을 나타냈으나, 20℃는 150 일에 낮아졌다가 270일에 약간 높아졌으며, 30℃는 차 이가 없었다. 분상과 입상유기질비료는 10℃와 20℃에 서 비슷한 경향을 나타냈으며, 분상유기질비료는 30℃
에서 150일에 낮았다가 270일에 높게 나타났고, 입상 유기질비료는 30℃에서 점차 낮아져 270일에는 34.0을 나타냈다. 비화산회토양은 유기질비료형태에 따라 pH 변화가 다르게 나타났으며, 분상유기질비료는 pH가 낮았고, 입상유기질비료는 토양 pH변화 폭이 컸으며, 토양 C/N비는 30℃에서 시간이 경과할수록 낮아졌다.
이는 토양과 혼합시 분상은 토양접촉 단면적이 넓고 분해가 빨리 이루어진 반면 입상은 접촉면이 작고 분 해가 되면서 일시에 H+ 이온이 증가하기 때문으로 생각되며 온도가 높고 시간이 지날수록 유기질비료의 분해가 잘되어 토양 C/N 비가 낮아지는 것으로 판단 된다.
흑색 화산회토양에서 토양 pH는 돈분과 음식물퇴비 가 분상과 입상유기질비료보다 높았으나 온도와 상관 없이 pH의 변화가 크지 않았으며, 시험토양보다 pH 가 높아졌다. 분상유기질비료는 온도에 상관없이 차 이가 없었고, 입상유기질비료는 온도가 높고 시간이 경과할수록 토양 pH는 낮아졌으나 변화폭은 크지 않 았고 시험토양보다 pH가 낮아졌다(Table 4.). C/N비 는 돈분과 음식물퇴비가 10℃에서 분상과 입상유기질 비료보다 높았으나 시간이 경과하면서 낮아지는 경향
Treatment
CON"
PMC‡ FWC OFPD OFPL
5.8 5.6 6.3 5.2 6.5
5.7 5.6 6.3 5.6 6.6
5.6 5.5 6.3 4.6 5.0
5.7 5.6 6.2 4.7 5.5
5.7 5.5 6.9 5.3 6.6
5.6 5.5 6.1 4.6 5.2
5.5 5.4 6.0 4.7 5.1
5.9 5.6 6.1 5.3 5.7
5.5 5.5 5.9 4.6 4.7
131.5 179.5 122.1 93.2 70.1
130.7 132.1 99.2 109.2 74.4
123.1 96.6 96.9 118.9 97.9
128.9 93.3 104.7 173.5 149.5
121.0 124.1 79.0 142.2 87.4
113.6 104.9 86.3 97.7 64.7
115.6 141.3 109.6 103.8 78.5
122.6 125.4 108.9 93.2 61.3
111.5 125.6 107.7 123.7 34.0 270d 150d 75d 270d 150d 75d 270d 150d 75d 270d 150d 75d 270d 150d 75d 270d 150d 75d
10℃
"
CON : Control
‡See table 2.
20℃ 30℃ 10℃ 20℃ 30℃
C/N ratio pH(1:5)
Table 3. The seasonal change of soil chemical properties in non-volcanic ash soil during incubated experiment period.
을 나타냈다. 20℃에서 돈분과 음식물퇴비는 분상과 입상유기질비료 보다 낮았다가 점차 높아졌으며, C/N 비는 입상유기질비료에서 감소 폭이 컸다. 30℃에서 돈분과 음식물퇴비는 분상과 입상유기질비료 보다 높 았으나 점차 낮아졌고, 분상유기질비료는 C/N비가 조 사시기에 비슷하게 나타났다. 돈분과 음식물퇴비는 토양 pH가 증가하였으나 입상과 분상유기질비료는 낮아졌다. 유기물이 일정한 온도와 수분조건이 유지 되면 미생물에 의하여 분해가 일어나며 토양 pH가 높아졌다가 낮아지는 것은 유기물의 분해가 일어나면 서 토양중 NO3-N 함량과 H+ 이온의 감소와 증가에 기인하는 것으로 토양내에서 유기물의 무기화와 유기 화가 동시에 일어나는 것으로 추정된다. Kelly and Henderson(1978)은 양분의 공급은 토양 pH변화에 의 하여 간접적으로 미생물활성에 영향을 준다고 하였 다. 또한 화산회토양의 알루미늄독성에 기인하여 항 온배양과정에서 미생물의 개체수와 활성이 영향을 받 아 온도, 유기물 종류와 형태, 분석시기에 따라 C/N 비가 다르게 나타나는 것으로 생각된다(Nobili, 2006).
비화산회토양의 pH가 화산회토양 보다 높은 것은 유 기물이 무기화되면서 NH4-N의 함량이 증가하면서 토 양에 흡착되고, 화산회토양은 탄소와 질소의 무기화 율이 낮았기 때문으로(Saito, 1990; Takenaka and Hayano, 1999), 온도와 토양특성, C/N비를 고려하면 비화산회토양이 화산회토양보다 분해가 잘 되는 것으 로 판단된다.
Microbial biomass C와 N 함량 변화 유기물이 분해 되면 탄소와 질소성분의 일부는 미생물 몸체 구성에 이용되는데 미생물체량 C와 N은 미생물의 밀도를 나 타내어 미생물의 활성을 평가하는 지표가 된다.
Alvarez et al.(1995)은 미생물체량 C가 토양 생물학 적 활성의 지표로서 사용될 때 온도의 영향은 중요하 다고 하였다. 토양별 유기물원의 항온배양에 따른 미 생물체량 C 함량 변화는 Fig. 1.에 나타내었다. 미생 물체량 C 는 비화산회토양에서 75일에 10℃와 20℃는 각각 분상유기질비료(320.1, 567.7mg kg-1)가 돈분퇴비
(167.0, 250.1mg kg-1)보다 약 2배정도, 30℃는 음식물 퇴비가 189.9 mg kg-1 으로 돈분퇴비 120.1 mg kg-1보 다 많았으며, 20℃에서 미생물체량 C 함량이 높게 나 타났다. 270일은 75일보다 미생물체량 C함량이 낮아 졌으며 10℃는 입상유기질비료(99.6)가 돈분퇴비(48.3 mg kg-1)보다 약 2배정도, 20℃는 입상유기질비료 (114.4)가 돈분퇴비(89.2 mg kg-1)보다 많았으며, 온도 가 높을수록 증가하는 경향이었다. 화산회토양에서 75 일에 입상유기질비료는 10℃(559.7), 20℃(660.5), 30 (689.6 mg kg-1)에서 돈분퇴비 보다 10℃(214.3), 20℃
(320.6), 30℃(344.4 mg kg-1)에서 2배 이상 많았으며 분상유기질비료를 제외하고 온도가 높을수록 미생물 체량이 증가하는 경향을 나타냈다. 270일은 75일보다 미생물체량 C함량이 낮았으며 온도가 높을수록 증가 하였으나 10℃에서 입상유기질비료(117.1)가 음식물퇴 비(39.6 mg kg-1)보다 약 3배, 20℃에서 분상유기질비 료(153.5)가 음식물퇴비(105.9 mg kg-1)보다 약 0.5배 많았다. 30℃에서는 입상유기질비료(195.1)가 대조구 (85.3 mg kg-1)보다 약 2배 많았다. 시험결과 비화산 회토양보다 화산회토양에서 온도가 높고 유기질비료 처리시 미생물체량 C 함량이 높은 경향이었으며 유기 물이 분해가 될 수록 미생물체량은 토양에 상관없이 감소하였다. 비화산회토양에서 75일은 20℃에서 미생 물체량이 가장 많았으나 270일은 30℃에서 미생물체 량이 많아지는 경향을 나타냈다. 화산회토양은 온도 가 높을수록 미생물체량이 증가하여 비화산회토양보 다 높다고 보고하였으며(Nishiyama et al., 2001), Takenaka and Hayano(1999)는 화산회토양은 유기물 복합체를 형성하고 있어 분해가 어려워 유기탄소 무 기화율이 2.5배정도가 낮다고 보고하였다. 따라서 미 생물체량 C 추출시 수용성 유기탄소의 영향으로 미생 물체량이 높게 나타난 것으로 판단되며, 화산회토양 에서 미생물체량 분석에 대한 새로운 방법이 강구되 어야 할 것으로 생각된다. Mondini et al.(2008)은 어 분과 골분의 토양공급은 미생물의 활성과 양을 증가 시킨다고 했는데, 미생물체량이 유기질비료에서 높은 것은 돈분과 음식물퇴비는 수분조절제로 톱밥을 이용 Treatment
CON"
PMC FWC OFPD OFPL
5.0 5.4 5.6 4.7 4.7
5.1 5.5 5.7 4.8 4.6
4.9 5.4 5.6 4.6 4.5
5.1 5.5 5.7 4.6 4.5
5.1 5.5 5.6 4.7 4.6
5.1 5.5 5.5 4.5 4.5
5.0 5.4 5.4 4.7 4.3
5.0 5.5 5.6 4.7 4.7
5.0 5.3 5.4 4.6 4.4
148.8 171.0 169.3 137.1 134.5
144.8 154.7 154.5 130.9 131.8
140.4 153.4 159.9 123.1 129.4
187.6 141.8 130.8 165.6 190.3
138.7 160.8 155.7 134.5 129.4
142.9 148.0 150.1 128.2 131.6
135.2 172.2 162.1 134.5 156.9
146.6 162.1 158.2 131.3 127.0
141.0 155.0 154.7 134.9 133.6 270d 150d 75d 270d 150d 75d 270d 150d 75d 270d 150d 75d 270d 150d 75d 270d 150d 75d
10℃
"
See table 3.
20℃ 30℃ 10℃ 20℃ 30℃
C/N ratio pH(1:5)
Table 4. The seasonal change of soil chemical properties in volcanic ash soil during incubated experiment period.
한 반면에 유기질비료는 어분과 골분을 이용함으로써 분해가 빠르게 나타나 토양중 유기탄소함량을 증가시 키기 때문으로 생각된다. 토양 미생물체량 C는 토양 총유기탄소 함량과 상관관계가 높고, 토양 pH, 온도, 수분함량, 기질이용성, 미생물의 다양성, 토양 이화학 적특성 등에 영향을 받으며, 유기질비료 시비는 비료 를 시비하지 않는 것 보다 토양호흡활성을 높인다고 보고하였다(Anderson, 1999; Cerhanova et al., 2006;
Goyal et al., 2000; Mondini et al., 2008; Paul and Clark, 1989).
토양미생물체량 N의 함량변화는 Fig. 2.에 나타내었 다. 비화산회토양은 150일에 미생물체량 N함량이 10
℃에서 음식물퇴비(270.4)가 분상유기질비료(66.7)보 다 4배, 돈분퇴비(28.6 mg kg-1)보다 9배 높게 나타났 다. 20℃와 30℃에서 입상 유기질비료는(232.0, 176.9 mg kg-1) 돈분퇴비(17.5, 32.2 mg kg-1)보다 각각 12 배, 5배 높았으며, 음식물퇴비는 10℃>20℃>30℃, 분상 과 입상유기질비료는 30℃>20℃>10℃ 순으로 높게 나
타났다. 270일에는 150일보다 미생물체량 N함량이 낮 았으며 10℃와 20℃에서 입상유기질비료가 각각 122.9, 96.7 mg kg-1 으로 높았으며 30℃에서는 유기물 원별로 통계적인 유의성이 나타나지 않았고, 입상유기 질비료를 제외하고 20℃>10℃>30℃ 순으로 높게 나타 났다. 화산회토양에서 150일에 10℃에서는 음식물퇴비 가 174.7, 20℃에서는 입상유기질비료가 158.2, 30℃에 서는 돈분퇴비가 336.7 mg kg-1으로 가장 높았으며, 돈 분퇴비는 온도가 높을수록 증가하였고 20℃>10℃>30℃
순으로 높게 나타났다. 분상과 입상유기질비료에서는 270일이 150일보다 낮았으며 10℃는 돈분퇴비(43.9)와 입상유기질비료(43.3)가 비슷하였으며 대조구(14.8 mg kg-1)보다 약 3배정도 높았다. 20℃는 음식물퇴비(58.6) 가 대조구(17.3 mg kg-1)보다 3배나 높았고, 30℃는 음 식물퇴비(16.9)가 돈분퇴비(2.5 mg kg-1)보다 6배나 높았으며 음식물퇴비를 제외하고 10℃>20℃>30℃ 순 으로 높게 나타났다. 150일에 비화산회토양의 10℃, 음식물퇴비에서 미생물체량 N이 높게 나타난 것은
Fig. 1. The change of microbial biomass C content during incubated period. (NVBS : Non- volcanic ash soil, VBS : Volcanic ash soil,
DMRT(p=0.05))분석시료에 NH4-N 함량이 많았기 때문으로 추정되 며, 화산회토양의 30℃, 돈분퇴비에서도 다른 유기물 원과 비교하여 느리게 무기화 되면서 나타난 결과로 생각된다. 이상의 결과 미생물체량 N은 비화산회토양 에서는 유기질비료가, 화산회토양에서는 돈분과 음식 물퇴비가 높았고, 유기물이 분해가 될 수록 토양에 상 관없이 감소하는 것으로 나타났다. 유기물이 분해가 되면 일부는 미생물 몸체를 구성하는 질소원으로 사 용되는데 유기물원과 토양에 따라 미생물체량 N이 차이가 나는 것은 토양특성, 유기물원의 구성과 시용 형태, 토양질소의 무기화와 관계가 깊은 것으로 생각 된다. Manzoni and Porpotato(2007)는 토양에서 질소 순환은 환경적인 요인과 생물적 활성에 의하여 진행 되고 토양유기물의 화학적 조성과 미생물의 양분요구 도가 영향을 준다고 보고하였고, Saito(1990)와 Deenik(2006)은 화산회토양의 유기태질소는 미생물분 해에 대하여 높은 저항성을 가져 비화산회토양보다 질소 무기화율이 2배나 낮다고 보고하였다.
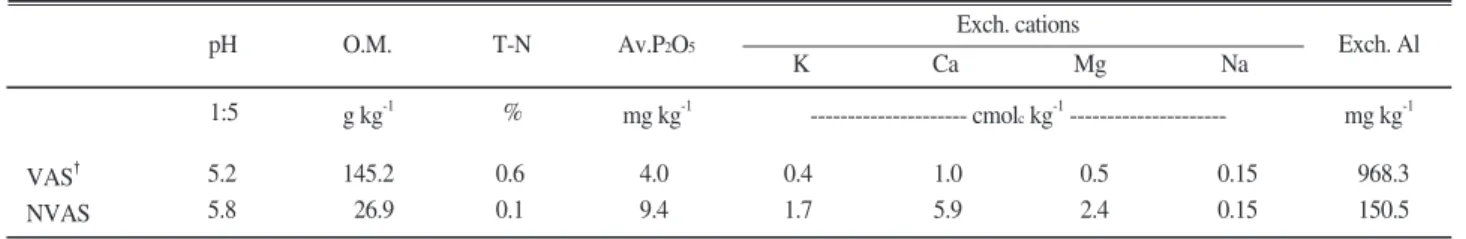
인지질 지방산 함량 변화 온도별 유기물원의 항 온배양에 따른 총 인지질 지방산함량의 변화는 Fig.
3.에 나타냈다. 비화산회토양은 75일에 인지질 지방산 함량이 10℃에서 입상유기질비료(164.3)가 음식물퇴비 (89.3 n mol g-1)보다 약 2배정도 높았다. 20℃, 30℃
에서는 음식물퇴비가 각각 170.5, 157.5 n mol g-1로 가장 높았으며, 유기물처리에 따른 인지질지방산함량 은 20℃에서 높게 나타났다. 270일은 75일보다 낮았으 며, 10℃에서는 분상유기질비료(127.4)가 돈분퇴비 (53.6 n mol g-1)보다 2배정도, 20℃에서는 입상유기질 비료(83.0)가 돈분퇴비(35.9 n mol g-1)보다 2배 이상 높았다. 30℃에서는 음식물퇴비가 82.6 n mol g-1를 나 타냈으며 유기질비료는 10℃, 돈분과 음식물퇴비는 30
℃에서 높았다. 화산회토양은 75일에 인지질 지방산 함량이 10℃와 20℃에서 입상유기질비료가 73.6, 203.5 n mol g-1로 돈분퇴비 20.9, 60.2 n mol g-1보다 3배 이 상 높았다. 30℃에서는 분상유기질비료가 84.7 n mol g-1가장 높았으며 유기물원별로는 분상유기질비료>입 상유기질비료>음식물퇴비>돈분퇴비 순이었고, 유기물
Fig. 2. The change of microbial biomass N content during incubated period. (See Fig. 1.)
처리에 따른 인지질지방산함량은 20℃에서 높게 나타 났다. 270일은 75일보다 낮았으며 10℃에서는 분상유 기질비료 169.4 n mol g-1, 20℃에서는 음식물퇴비 167.2 n mol g-1, 3℃에서는 돈분퇴비가 153.6 n mol g-1로 높았다. 10℃에서는 유기질비료, 30℃에서는 돈 분과 음식물퇴비가 높게 나타났으나 75일보다는 증가 하였으며, 20℃에서 돈분과 음식물퇴비는 증가하였고 분상과 입상유기질비료는 감소하였다. 비화산회토양 은 75일, 20℃를 제외하고 화산회토양보다 높았으나 270일 경과후 화산회토양은 모든 유기물처리에서 인 지질지방산의 함량이 증가하였다. 살아있는 미생물의 활성을 측정하는 인지질지방산 함량을 분석하였을 때, 항온배양초기에 미생물밀도가 낮은 풍건토양을 이용함으로써 토양특성보다는, 유기물이 분해가 되면 서 비화산회토양에서는 인지질지방산 함량이 높게 나 타났으며, 시간이 경과하면서 화산회토양에서 유기태 질소가 무기화되고, 통기성이 양호하여 인지질지방산 함량이 증가한 것으로 생각된다. Sharifi et al.(2008) 은 토양 유기태질소의 활성형태는 경작방법의 영향을
받으며 비경작지토양에서 이분해성 무기화질소함량이 관행경작지보다 높았고, 활성화된 유기태질소형태가 증가하는 경향이었다고 하였다. 화산회토양은 유기물 과 allophane이 복합체를 형성하여 다량의 알루미늄에 의하여 토양미생물의 활성이 낮다(Tokashiki and Wada, 1975; Henmi et al., 1976; Masami, 2002;
Ugolini and Dahlgren, 2002)고 하였는데 이는 경작토 와 비경작토, 건토와 습토, 토양 C와 N함량 차이 등 이 토양 유기물 분해와 미생물활성에 영향을 주는 것 으로 생각된다. 인지질 지방산은 미생물체량과 군집 구성 등에 대한 정보를 제공한다고 하였으며(Green and Scow, 2008), 토양 pH는 토양미생물 활성에 영향 을 나타내며, pH가 증가하면 PLFA함량도 증가한다 고 보고하였다(Baath and Anderson, 2003; Kemmitt, 2006). 비화산회토양의 pH가 화산회토양 보다 높았으 나 인지질 지방산함량이 낮은 것은 토양중의 질소순 환 미생물의 밀도가 높아 토양 중 NO3-N함량이 낮 고, 용적밀도가 크고 점토함량이 높아 통기성이 불량 하기 때문이라고 판단된다. Cookson et al.(2007)은 배
Fig. 3. The change of total-PLFA content during incubated period. (See Fig. 1.)
양온도, 토양 pH, 유기물함량은 PLFA 패턴을 예측하 는데 있어 중요하며 토양 무기태 질소함량과 상관이 높다고 보고하였다. 따라서 질소 무기화 포텐셜이 높 은 유기물원을 이용했을 경우에는 인지질지방산함량 이 증가하는 것으로 판단된다.
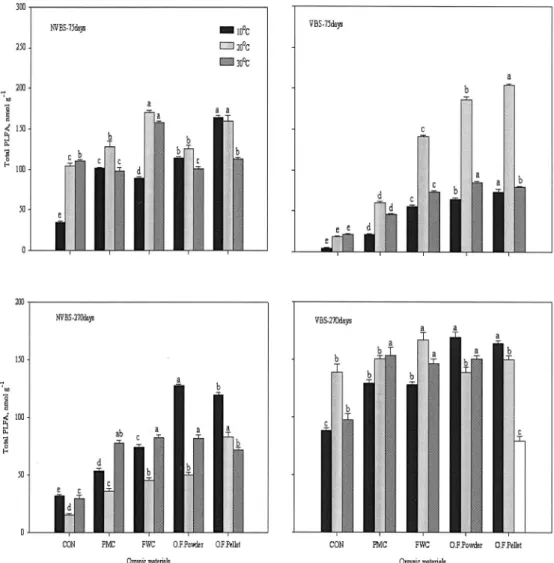
토양효소활성 변화 토양에 유기물을 첨가하면 미 생물체량과 효소활성이 증가하며 토양 pH, 유기탄소, 질소함량은 Urease활성과 상관관계가 높다고 보고하 였다(Dinesh et al., 1998). Urease는 요소를 CO2와 NH3로 분해과정에 촉매역할을 하는데, 토양별 유기물 원에 따른 Urease활성 변화는 Fig. 4.에 나타냈다. 비 화산회토양은 150일에 모든 온도처리에서 입상유기질 비료가 가장 높게 나타났으며 10℃에서는 입상유기질 비료(75.0)가 돈분퇴비(3.0 ug NH4-N g-1 2h-1)보다 25배, 20℃에서는 입상유기질비료(16.3)가 돈분퇴비 (3.1 ug NH4-N g-1 2h-1)보다 5배, 30℃에서는 입상유 기질비료(4.6)가 돈분퇴비(2.8 ug NH4-N g-1 2h-1)보 다 약간 높게 나타났으며 온도가 낮을수록 효소활성
은 낮아졌다. 270일은 150일보다 낮아졌으며, 입상유 기질비료와 10℃에서 높게 나타났다. 화산회토양은 150일에 10℃에서 Urease활성이 높았으며 입상유기질 비료(18.9)>음식물퇴비(10.0)>돈분퇴비(5.5 ug NH4-N g-1 2h-1)순으로 낮았다. 20℃에서는 유기물원간에 효 소활성차이가 없었고 30℃에서는 음식물퇴비가 2.2 ug NH4-N g-1 2h-1로 높았다. 270일은 150일보다 Urease활성이 낮았고, 10℃와 30℃에서는 유기물원간 에 효소활성의 차이가 없었으며, 20℃에서는 음식물퇴 비(0.28)가 입상유기질비료(0.13 ug NH4-N g-1 2h-1) 의 2배정도 높게 나타났다. 토양에 유기물을 첨가하면 토양 유기태질소는 무기화되면서 NH4-N을 방출하는 데 NH4-N는 초기에 높고 분해시간이 오래되고 온도 가 높을수록 낮게 나타나며 Urease 활성은 증가한다 고 보고하였다(Klose and Tabatabai, 2000). 항온배양 시험결과 Urease 활성은 비화산회토양, 유기질비료처 리, 10℃에서 높았고 시간이 경과할수록 토양에 상관 없이 낮아지는 경향을 나타내었는데, 10℃에서 높게 나타난 것은 30℃보다 무기화가 느리게 진행됐기 때
Fig. 4. The change of urease activity during incubated period. (See Fig. 1.)
문으로 생각되며, 유기질비료 토양처리시 미생물체량 N과 Urease활성은 상관관계가 높다고 보고한 결과와 일치하는 경향을 나타냈다(Bremner and Mulvaney, 1978; Roscoe et al., 2000).
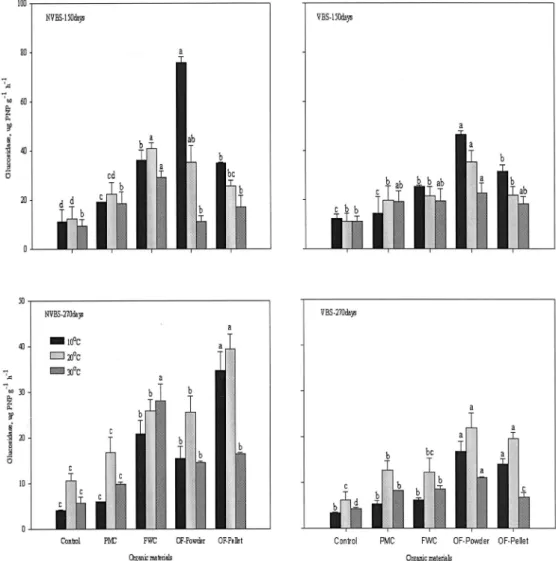
유기물의 분해 과정에서 촉매역할을 하는 β- glucosidase의 활성 변화는 Fig. 5.에 나타냈다. 비화산 회토양은 150일에 10℃와 20℃에서는 입상유기질비료 (34.7, 39.5 ug PNP g-1 h-1)가 돈분퇴비(6.1, 16.7 ug PNP g-1 h-1)보다 각각 5배, 2배 이상 높았다. 30℃는 음식물퇴비(28.2)가 돈분퇴비(9.8 ug NH4-N g-1 2h-1) 보다 약 3배정도 높게 나타났다. 돈분과 음식물퇴비는 20℃에서 높게 나타났으며 유기질비료는 10℃에서 높 았고 온도가 낮을수록 효소활성은 낮아졌다. 270일은 150일보다 높았으며 10℃에서는 분상유기질비료(34.7, 39.5 ug PNP g-1 h-1)가 돈분퇴비(6.1, 16.7 ug PNP g-1 h-1)보다 각각 5배, 2배 이상 높았다. 30℃에서는 음식물퇴비(28.2)가 돈분퇴비(9.8 ug NH4-N g-1 2h-1) 보다 약 3배정도 높게 나타났다. 화산회토양은 150일 에 10℃와 20℃에서는 분상유기질비료(46.4, 35.3 ug
PNP g-1 h-1)가 돈분퇴비(14.3, 19.8 ug PNP g-1 h-1) 보다 각각 3배, 1.5배, 30℃에서는 분상유기질비료 (22.7)가 대조구(11.1 ug NH4-N g-1 2h-1)보다 2배정 도 높게 나타났다. 돈분과 음식물퇴비는 20℃에서 높 게 나타났으며 유기질비료는 10℃에서 높았고 온도가 낮을수록 효소활성은 낮아졌다. 270일은 150일보다 낮 았으며 모든 온도에서 분상유기질비료가 높았으며 10
℃, 20℃, 30℃에서 각각 16.8, 22.0, 11.1 ug PNP g-1 h-1로, 돈분퇴비 5.4, 12.7, 8.3 ug PNP g-1 h-1보다 높 았다. β-glucosidase 활성은 비화산회토양이 화산회토 양보다 높았고 시간이 경과할수록 낮아졌다. 유기질 비료는 150일과 270일에 각각 10℃와 20℃에서 높았 으며, 음식물퇴비는 비화산회토양의 30℃에서 높게 나 타났다. 토양에 투입된 유기물은 미생물에 쉽게 분해 가 되거나 분해에 대한 저항을 나타낸다고 보고하였 는데(Alvarez et al., 1995), 이는 톱밥을 많이 혼합 한 돈분퇴비 보다 골분, 어분 등의 이분해성 유기물이 포 함된 유기질비료가 쉽게 분해가 되는 것으로 판단된 다. Sardans et al.(2008)은 Urease와 β-glucosidase 활
Fig. 5. The change of β
β-glucosidase activity during incubated period. (See Fig. 1.)
성은 겨울에는 토양온도와 정의상관, 여름에는 부의 상관을 나타내며, 온난화는 겨울에 토양효소활성을 증가시키지만 토양유기물 함량의 변화를 가져오지 않 는다고 보고하였다. 이것은 여름철 고온보다 겨울철 저온상태에서의 온도의 증가는 효소의 활성을 높이는 것으로 생각되어 지며, 이 결과는 Knorr et al.(2005) 이 토양유기탄소는 온도에 민감하다고 보고한 결과와 유사하였다. 화산회토양에서 β-glucosidase 활성이 낮 은 것은 Anderson(1979)이 Al3+ 등 금속이온이 효소 활성에 영향을 주어 유기물분해를 저해시킨다고 보고 한 결과와 일치하는 경향을 나타냈다.
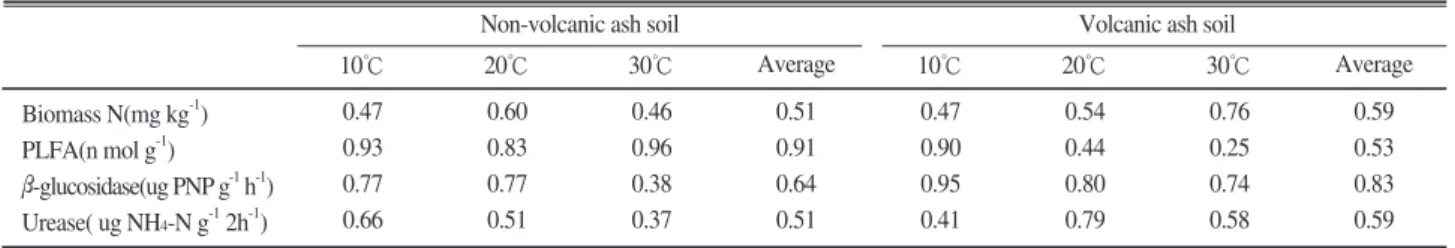
Microbial biomass C와 미생물활성 지표간의 상관관계 화산회토양과 비화산회토양의 토양미생물체량 C와 미 생물활성지표 간의 상관관계를 분석한 결과는 Table 5.에 나타냈다. 비화산회토양에서 Microbial biomass N 함량은 20℃에서 0.60으로 가장 높았으며, PLFA는 10℃(0.93), 20℃(0.83), 30℃(0.96)으로 상관계수가 높 게 나타났다. β-glucosidase는 10℃와 20℃에서 각각 0.77로 비슷하였으며 30℃에서는 0.38로 상관계수가 낮았다. Urease는 10℃에서 0.66을 나타냈으며 온도가 낮을수록 상관계수도 낮아졌다. 화산회토양에서 Microbial biomass N함량은 온도가 10℃ 이상 올라갈 수록 높아져 30℃에서 0.76으로 가장 높았으며, PLFA와 β-glucosidase는 10℃에서 각각 0.90, 0.95로 높았으며 온도가 올라갈수록 상관계수는 낮아졌다.
Urease는 20℃에서 0.79로 높았다가 30℃에서 0.58로 낮아졌다. 결론적으로 토양미생물체량 C와 미생물활 성 지 표 간 의 상 관 계 수 는 비 화 산 회 토 양 에 서 PLFA(r2=0.91), 화산회토양에서 β-glucosidase (r2=0.83)가 높았고 미생물활성지표는 토양과 온도에 따라 상관관계가 다르게 나타나 토양별로 각각 PLFA와 β-glucosidase를 이용하여 미생물의 활성을 평가할 수 있을 것으로 생각된다. 이는 토양 전체 Urease활성의 46%가 미생물체량과 관계가 있으며, 미 생물체량 C와 상관관계가 높다는 결과와 일치하였다 (Klose and Tabatabai, 1999, 2000; Perucci, 1990). 토 양효소활성과 온도별 상관계수의 차이는 토양 유기물 을 분해하는데 있어 효소활성에 영향을 주는 온도의 상대적인 차이에 기인하는 것으로 생각되며(Koch et
al., 2007), 비화산회토양에서 30℃가 10℃ 보다 낮은 것은 유기물 종류차이, 풍건토양의 낮은 미생물밀도, 수분첨가에 따른 토양수분함량 변화, 높은 온도로 조 사시기보다 빠르게 유기물분해가 진행된다는 보고와 일치하는 경향을 나타내었다(Debosz et al., 1999;
Sardans et al., 2008).
요 약
유기물원을 항온 배양했을 때 온도가 토양미생물체 량과 효소활성 및 PLFA함량에 미치는 영향을 토양 특성별로 평가하고자 입상과 분상 혼합유기질비료, 돈분퇴비, 음식물퇴비를 화산회토양과 비화산회토양 30 g에 2 g을 잘 혼합 후 10, 20, 30℃에서 항온배양 을 하면서 pH, 토양질소, 유기물 함량, Microbial biomass C와 N, 토양효소활성, 인지질지방산 함량을 분석하였다. 돈분과 음식물퇴비는 토양종류와 온도에 상관없이 토양 pH의 변화가 크지 않았으나 입상유기 질비료는 온도가 높을수록 낮아졌으며, 비화산회토양 에서 변화폭이 컸다. 미생물체량 C는 화산회토양의 경우 배양 온도가 높고 유기질비료를 처리할 때 높아 지는 경향이었지만 토양종류에 상관없이 점차 감소하 였다. 미생물체량 N은 비화산회토양에서 유기질비료, 화산회토양에서는 돈분과 음식물퇴비를 처리할 때 높 게 나타났다. 인지질 지방산함량은 항온배양 75일후 비화산회토양이 화산회토양보다 높았고, 270일후 화산 회토양에서 높게 나타났으나 점차 낮아지는 경향이었 다. Urease활성은 150일에 비화산회토양의 입상유기질 비료처리에서 10℃(75.0)>20℃(16.3)>30℃(4.6 ug NH4-N g-1 2h-1)순으로 10℃에서 가장 높게 나타났으 며 온도가 높고, 시간이 경과할수록 낮아졌으며 화산 회토양의 Urease활성은 10℃에서 높게 나타났다. β- glucosidase 활성은 비화산회토양이 화산회토양보다 높았고 시간이 경과할수록 낮아졌다. 토양미생물체량 C와 미생물활성지표간의 상관계수는 비화산회토양에 서 PLFA (r2=0.91), 화산회토양에서 β-glucosidase (r2=0.83)가 높았으며, 미생물활성지표는 토양종류와 온도에 따라 상대적으로 민감도가 다르게 나타났다.
Biomass N(mg kg-1) PLFA(n mol g-1) β-glucosidase(ug PNP g-1h-1) Urease( ug NH4-N g-12h-1)
Average 30℃
20℃
10℃
Average 30℃
20℃
10℃
0.47 0.93 0.77 0.66
0.60 0.83 0.77 0.51
0.46 0.96 0.38 0.37
0.51 0.91 0.64 0.51
0.47 0.90 0.95 0.41
0.54 0.44 0.80 0.79
0.76 0.25 0.74 0.58
0.59 0.53 0.83 0.59
Non-volcanic ash soil Volcanic ash soil
Table 5. Correlation coefficient between microbial activity indicators and microbial biomass C content.
인 용 문 헌
Agehara, S. and D.D. Warncke. 2005. Soil moisture and temperature effects on nitrogen release from organic nitrogen sources. AM. J.
Soil Sci. Soc. 69:1844-1855.
Alvarez, R., O.J. Santanatoglia, and R. Garcja. 1995. Effect of temperature on soil microbial biomass and its metabolic quotient in situ under different tillage systems. Biol. Fertil. Soils. 19(2):227- 230.
Amato, M. and J.N. Ladd. 1988. Assay for microbial biomass based on Ninhydrin -reactive nitrogen in extracts of fumigated soils. Soil Biol. Biochem. 20:107-I14.
Anderson, D.W. 1979. Processes of humus formation and transformation in soils of the Canadian Great Plains. Euro. J. Soil Sci. 30:77-84.
Anderson, J.M. 1991. The effects of climate change on decomposition processes in grassland and coniferous forests. Ecol.
Appl. 1:326 347.
Baath, E. and T.H. Anderson. 2003. Comparison of soil fungal/bacterial ratios in a pH gradient using physiological and PLFA-based techniques. Soil Biol. Biochem. 35(7):955-963.
Bligh, E.G. and W.J. Dyer. 1959. A rapid method for total lipid extraction and purification. Can. J. Biochem. Physiol. 37:911-917.
Bremner, J.M and R.L. Mulvaney. 1978. Urease activity in soil.
In:Burns RG(ed) Soil enzymes. Academic Press, London, pp 149- 196.
Cerhanova, D., J. Kubat, and J. Novakova. 2006. Respiration activity of the soil samples from the long-term field experiments in Prague. Plant Soil Environ. 52:21-28.
Cordovil, C.M.D.S., J. Coutinho, M. Goss, and F. Cabral. 2005.
Potentially mineralizable nitrogen from organic materials applied to a sandy soil: fitting the one-pool exponential model. Soil Use Manage. 21:65-72.
Cookson W.R., M. Osman, P. Marschner, D.A. Abaye, I. Clark, D.V. Murphy, E.A. Stockdale, and C.A. Watson. 2007. Controls on soil nitrogen cycling and microbial community composition across land use and incubation temperature. Soil Biol. Biochem.
39:744-756.
Debosz, K., P.H. Rasmussen, and A.R. Pedersen.1999. Temporal variations in microbial biomass C and cellulolytic enzyme activity in arable soils : effects of organic matter input. Appl. Soil Ecol.
13:209-218.
Deenik, J. 2006. Nitrogen mineralization potential in important agricultural soils of Hawaii. Uni. Hawaii, CTAHR. Soil Crop Manage. 15:1-5.
Dick, R. P.1992. A review: long-term effects of agricultural systems on soil biochemical and microbial parameters. Agric. Ecosyst.
Environ. 40: 25-36.
Dinesh, R., R.P. Dubey, and G.S. Prasad. 1998. Soil microbial biomass and enzyme activities as influenced by organic manure incorporation into soils of a rice-rice system. J. Agro. Crop Sci.
181:173-178.
Friedel, J. K. E. Otto, P. Michael, S. Michael, V. Tobias, S.
Michael.2006. Soil microbial biomass and activity : the effect of site characteristics in humid temperate forest ecosystems.J. plant nutrition & soil science.169:175-184
Goyal S., K. Sakamoto, and K. Inubashi. 2000. Microbial biomass and activities along an andosol profile in relation to soil organic matter level. Microb. Environ. 15:143-150.
Green, C.T. and K.M. Scow. 2008. Analysis of phospholipid fatty acids(PLFA) to characterize microbial communities in aquifers.
Hydrogeo. J. 8:126-141.
Henmi, T. and K. Wada. 1976. Morphology and composition of allophane. Am. Mineral. 61:379-390.
Hu, C. and Z. Cao. 2007. Size and activity of the soil microbial biomass and soil enzyme activity in long-term field experiments.
World J. Agri. Sci. 3(1):63-70.
Joa, J.H., J.H. Lee, H.Y. Won, S.G. Han, and H.C. Lim. 2008.
Effect of different soil managements on physical properties and microbial activities in citrus orchard soil. Korean J. Soil Sci. Fert.
41:279-284.
Joa, J.H., S.G. Han, H.Y. Won, H.C. Lim, H.N. Hyun, and J.S. Suh.
2009. Effect of different fertilization management practices on soil microbial activities and community structure in volcanic ash citrus orchard soil. Korean J. Soil Sci. Fert. 42:222-229.
Kao, P.H., C.C. Huan, and Z.Y. Hseu. 2006. Response of microbial activities to heavy metals in a neutral loamy soil treated with biosolid. Chemosphere. 64:63-70.
Kelly, J.M. and G.S. Henderson. 1978. Effects of nitrogen and phosphorus additions on deciduous litter decomposition. Am. J.
Soil Sci. Soc. 42:972-976.
Kemmitt, S.J., C.V. Lanyon, I.S. Waite, Q. Wen, T.M. Addiscott, N.R.A. Bird, A.G. O'Donnell, P.C. Brookes. 2008. Mineralization of native soil organic matter is not regulated by the size, activity or composition of the soil microbial biomass C new perspective. Soil Biol. Biochem. 40:61-73.
Kemmitt, S.J., D. Wright, K.W.T. Goulding, and D.L.Jones. 2006.
pH regulation of carbon and nitrogen dynamics in two agricultural soils. Soil Biol. Biochem. 38:898-911.
Kirschbaum, M.U.F. 1995. The temperature dependence of soil organic matter decomposition, and the effect of global warming on soil organic C storage. Soil Biol. Biochem. 27(6):753-760.
Klose, S. and M.A. Tabatabai. 1999. Urease activity of microbial biomass in soils. Soil Biol. Biochem. 31:205-211.
Klose, S. and M.A.Tabatabai. 2000. Urease activity of microbial biomass in soils as affected by cropping systems. Biol. Fertil.
Soils. 31:191-199.
Knorr, W., I.C. Prentice, J.I. House, and E.A. Holland. 2005. Long- term sensitivity of soil carbon turnover to warming. Nature. 433 298-301.
Koch, O., D. Tscherko, and E. Kandeler. 2007. Temperature sensitivity of microbial respiration, nitrogen mineralization, and potential soil enzyme activities in organic alpine soils. Global Biogeochem. Cycles. Japan. 21(4):GB4017
Lagomarsino, A., M.C. Moscatelli, A. D. Tizio, R. Mancinelli, S.
Grego, S. Marinari. 2009. Soil biochemical indicators as a tool to assess the short-term impact of agricultural management on
ˇ
changes in organic C in a Mediterranean environment. Ecological indicators. 9:518-527.
Manzoni, S., A. Porpotato. 2007. A theoretical analysis of nonlinearities and feedbacks in soil arbon and nitrogen cycles. Soil Biol. Biochem. 39:1542-1556.
Marschner, P., E. Kandeler, and B. Marschner. 2003. Structure and function of the soil microbial community in a long-term fertilizer experiment. Soil Biol. Biochem. 35:453-461.
Mondini, C., M.L. Cayuela, T. Sinicco, M.A.S. Monedero, E.
Bertolone, and L. Bardi. 2008. Soil application of meat and bone meal. short-term effects on mineralization dynamics an soil biochemical and microbiological properties. Soil Biol. Biochem.
40:462-474.
Nanzyo, M. 2002. Unique properties of volcanic ash soils. Glob.
Environ. Res. 6(2):99-112.
Nishiyama, M., Y. Sumikawa, G. Guan, and T. Marumoto. 2001.
Relationship between microbial biomass and extractable organic carbon content in volcanic and non-volcanic ash soil. Appl. soil Ecol. 17(2):183-187.
Nobili, D.M., M. Contin, and P.C. Brookes. 2006.Microbial biomass dynamics in recently air-dried and rewetted soils compared to others stored air-dry for up to 103 years. Soil Biol. Biochem.
38:2871-2881.
Paul, E.A. and F.E. Clark. 1989. Soil microbiology and biochemistry. Academic press, San Diego, CA.
Perucci, P. 1990. Effect of the addition of municipal solidwaste compost on microbial biomass and enzyme activities in soil.Biol Fertil Soils. 10:221-226.
RDA. 1988. Methods for chemical analysis of soil. Institute of Agricultural Technology.
Roscoe, R., C.A. Vasconcellos, A.E.F. Neto, G.A.A. Guedes, and L.A. Fernandes. 2000.Urease activity and its relation to soil organic matter, microbial biomass nitrogen and urea-nitrogen assimilation by maize in a Brazilian Oxisol under no-tillage and tillage systems.Biol Fertil Soils. 32:52-59.
Saito, M. 1990. Nitrogen mineralization parameters and its availability indices of soils in Tohoku district, Japan: Their relationship. Jpn. J. Soil Sci. Plant Nutr. 61(3):265-272.
Sardans, J., J. Penuelas, and M. Estiarte. 2008. Changes in soil enzymes related to C and N cycle and in soil C and N content under prolonged warming and drought in a Mediterranean shrubland. Appl. soil ecol. 39:223-235.
Satti, P., M.J. Mazzarino, L. Roselli, and P. Crego. 2007. Factors affection soil P dynamics in temperate volcanic soils of southern argentina. Geoderma. 139:229-240.
Sharifi, M., B.J. Zebarth, D.L. Burton, C.A. Grant, G.A. Porter.
2008. Organic amendment history and crop rotation effects on soil nitrogen mineralization potential and soil nitrogen supply in a potato cropping system. Agron. J. 100:1562-1572.
Song, K. C. 1990. Andic properties of major soils in Cheju island.
Ph. D. Thesis. Seoul National University. Suwon, Korea.
Stanford, G. and S.J. Smith. 1972. Nitrogen mineralization potentials of soils. Am. J. Soil Sci. Soc. 36:465-472.
Takenaka, M and K. Hayano. 1999. Investigation on the influence of the global warming on the mineralization of soil organic matter.
Rese. Out. Agriculture, Forestry Fishery Tech. Bureau. 339:232- 236.
Timothy, R.K., and R.P. Dick. 2004. Differentiating microbial and stabilized -glucosidase activity relative to soil quality. Soil Biol.
Biochem. 36:2089-2096.
Tokashiki T. and K.Wada. 1975. Weathering implications of the mineralogy of clay fractions of two Ando soils. Kyushu.
Geoderma. 14:47-62.
Ugolini, F.C. and R.A. Dahlgren. 2002. Soil development in volcanic ash. Glob. Environ. Res. 6(2):69-81.
Vance, E.D., P.C. Brookes, and D.S. Jenkinson. 1987. Microbial biomass measurements in forest soils : the use of the chlroroform fumigation incubation method in strongly acid soils. Soil Biol.
Biochem. 19:697-702.