688
©The Korean Society of Food Science and Technology
홍삼 사포닌 분획의 Nrf2 Keap1 신호전달체계 조절을 통한
지방축적 및 활성산소종 억제효과
김채영
1·강보빈
1·황지수
1·최현선
2,*
1고려대학교 보건과학대 의생명융합과학과, 2서울여자대학교 식품공학과
Red ginseng-derived saponin fraction inhibits lipid accumulation and
reac-tive oxygen species production by activating nuclear factor
(erythroid-derived 2)-like 2 (Nrf2)/Kelch-like ECH-associated
protein 1 (Keap1) pathway
Chae-Young Kim1, Bobin Kang1, Jisu Hwang1, and Hyeon-Son Choi2,*
1Department of Integrated Biomedical and Life Science, Graduate School, Korea University 2Department of Food Science and Technology, College of Natural Science, Seoul Women’s University
Abstract This study aimed to investigate the effects of red ginseng-derived saponin fraction (SF) on lipid accumulation, reactive oxygen species (ROS) production, and nuclear factor (erythroid-derived 2)-like 2 (Nrf2)/Kelch-like ECH-associated protein 1 (Keap1) signaling during adipocyte differentiation. SF effectively inhibited lipid accumulation, with the down-regulation of adipogenic factors such as peroxisome proliferator-activated receptor gamma (PPARγ) and CCAAT/enhancer-binding protein alpha (C/EBPα). A high dose of SF decreased the protein levels of PPARγ and C/EBPα by over 90% compared to the control. SF-mediated downregulation of adipogenic factors was due to the regulation of early adipogenic factors including C/EBPβ and Krüppel-like Factor 2 (KLF2). In addition, SF (200 μg/mg) decreased intracellular ROS generation by 40% during adipocyte differentiation. However, the SF significantly upregulated Nrf2 and its target proteins, hemoxygenase-1 (HO-1) and NADPH dehydrogenase quinone 1 (NQO1). Furthermore, SF (200μg/mg) promoted the nuclear translocation of Nrf2. The SF-mediated reduction of lipid accumulation was associated with the regulation of the Nrf2/Keap1 pathway.
Keywords: red ginseng-derived saponin fraction, lipid accumulation, ROS production, Nrf2/Keap1 pathway
서
론
인삼은 두릅나뭇과(Araliaceae), 인삼속(Panax) 다년생 식물의 뿌리를 말하며 민간 또는 한방 약초로 오랫동안 이용되어왔고 여 러 종류의 인삼 중에서 한국의 인삼(Panax ginseng C. A. Meyer) 이 가장 유명하다(Chang, 2003; Chang 등, 2006). 인삼은 특유의 사포닌 성분인 진세노사이드를 함유하고 있으며 그 함량과 조성 은 산지와 재배 및 가공방법에 따라 다르다(Chang, 2003; Lee와 Do, 2001). 특히 높은 온도와 증기의 가공과정을 거친 인삼을 홍 삼이라 하며 이때 진세노사이드의 조성과 함량이 가공 전보다 더 우수한 것으로 알려져 있다(Nam, 2005). 홍삼은 세포와 동물실험 에서 다양한 생리활성을 나타내는 것으로 보고되고 있는데 면역 시스템을 활성화하며, 심혈관질환을 예방하고, 중추신경계를 보 호하는 기능은 물론 항암, 항당뇨, 산화방지 등의 효과를 나타내 는 것으로 알려져 있다(Keum 등, 2000; Kim 등, 2004; Kim 등, 2012; Konno와 Hikino, 1987; Yoon과 Joo, 1993). 이러한 홍삼의 생리활성들은 주로 홍삼에서 유래하는 사포닌 성분에 의한 것으 로 보고되고 있다(Attele 등, 1995). 비만은 에너지 불균형으로부터 오는 과도한 지방의 축적에서 비롯되며 현대의 대표적인 대사질환이라고 말할 수 있다(Spiegelman 등, 1993). 비만은 그 자체보다는 다른 질환들, 즉 당뇨병, 심혈 관 질환, 고혈압, 암 등의 성인병을 유발 할 수 있는 주요 원인 으로 지목되고 있기 때문에 비만에 대한 예방 및 관리가 중요하 게 여겨진다(Spiegelman 등, 1993). 비만 시 지방의 축적은 주로 지방세포에서 지방분화라는 과정을 거쳐 이루어진다(Lowe 등, 2011). 지방분화과정 동안 PPARγ와 C/EBPα와 같은 다양한 지방 분화 전사인자들의 발현과 상호작용이 일어남으로써 지방대사관 련 유전자들이 발현되어 지방합성이 이루어진다(Kersten, 2001). ROS는 세포대사과정에서 자연적으로 생산되는 분자로 세포의 정상적인 기능과 신호전달시스템에서 중요한 역할을 한다(Halliwell 등, 1992). 그러나 과도한 ROS의 생산은 산화스트레스를 유도하 고 세포 내 분자들의 손상을 일으켜 세포의 정상적인 기능과 역 할을 방해한다(Halliwell 등, 1992; Oakley 등, 2009). 산화스트레 스는 비만, 염증질환, 암 등의 다양한 질환의 발병과 진행에 중 *Corresponding author: Hyeon-Son Choi, Department of Food
Sci-ence and Technology, College of Natural SciSci-ence, Seoul Women’s University, Seoul 01797, Korea
Tel: +82-2-970-5683 Fax: +82-2-970-5977 E-mail: [email protected]
Received September 18, 2018; revised November 24, 2018; accepted November 28, 2018
요한 역할을 하는 위험요인중의 하나로 인식되고 있다(Halliwell 등, 1992). 특히 비만은 ROS로 인한 산화스트레스가 항진되어 있 는 질병으로 알려져 있다(He 등, 2012). 하지만 세포는 과도한 ROS로 인한 세포 내 스트레스와 독성을 완화하는 조절시스템을 가지고 있으며 대표적인 것이 nuclear factor (erythroid-derived 2)-like 2 (Nrf2)/Kelch-2)-like ECH-associated protein 1 (Keap1) 신호 전달체계이다(He 등, 2012). Nrf2는 산화스트레스로부터 세포를 보호하는 역할을 하는 전사인자로 정상적인 상태에서는 세포질 에서 Keap1과 복합체를 형성하다가 ROS나 친전자체들의 자극에 의해 Keap1이 분해되고 이로 인해 Nrf2가 복합체로부터 분리되 어 핵 안으로 들어가서 ROS조절 및 세포독성을 보호할 수 있는 유전자들의 발현을 촉진하는 것으로 알려져 있다(He 등, 2012). 따라서 지방축적 및 비만의 중요한 원인으로 과도한 ROS에 인 한 산화스트레스를 들 수 있으며 세포 내 ROS 수준을 통제 할 수 있는 Nrf2의 조절을 통해 지방축적 및 비만을 조절할 수 있 을 것이다. 실제 비만과 Nrf2의 관계에 대한 연구들이 다수 보고 되어 왔다(Gaikwad 등, 2001; Pi 등, 2010; Shin 등, 2007). 또한, 홍삼 유래 항비만에 대한 연구도 종종 보고되고 있지만 홍삼사 포닌 분획의 지방분화 및 지방축적억제효과와 Nrf2/Keap1 신호 전달체계와의 관계에 대한 연구는 미비하다. 본 연구에서는 홍삼 유래 사포닌 분획의 지방축적에 대한 효과가 ROS조절 및 Nrf2/ Keap1 신호전달체계와 밀접한 관련이 있음을 보여준다.
재료 및 방법
재료 및 시약 홍삼에서 유래하는 사포닌 분획(SF)은 고려인삼학회로부터 얻 었다. 진세노사이드 표준물질은 Embo 실험실(Daejeon, Korea)에 서 구입하였다. 3T3-L1 전지방세포는 American Type Cultured Collection (ATCC, Rockville, MD, USA)에서 분주 받아 실험하였 다. Dulbecco’s modified Eagle’s medium (DMEM)은 Welgene (WelGENE, Daegu, Korea)에서 구입하였고, 소의 혈청(bovine serum)과 인슐린(insulin)은 Gibco (Gaithersburg, MD, USA)에서 구입하였다. 소태아혈청(fetal bovine serum), 항생제(penicillin와 streptomycin), phosphate-buffered saline (PBS)은 Hyclone (Logan, UT, USA)에서 구입하였다. 싸이아졸릴블루 테트라졸륨 브로민화 물(thiazolyl blue tetrazolium bromide, MTT), 다이메틸설폭사이드 (DMSO), 덱사메타손(dexamethasone, DEX), 3-isobutyl-1-methylx-anthine (IBMX), Oil red O 시약은 Sigma-Aldrich Co. (St. Louis, MO, USA)에서 구입하였다. Nrf2, HO-1 항체는 Santa Cruz Bio-technology, Inc. (Santa Cruz, CA, USA), 그 외 PPARγ, C/ EBPα, C/EBPβ, FABP4, KLF2, GAPDH 등의 항체는 Cell Sig-naling (Danvers, MA, USA)사로부터 구입하였다. 그 외의 모든 시약 및 용매는 Sigma-Aldrich Co. (St. Louis, MO, USA)에서 구입하였으며 일급 또는 특급 이상의 등급을 사용하였다.HPLC 분석
SF의 진세노사이드(ginsenoside) 조성은 Imtakt Cadenza CD-C18 칼럼(4.6 mm×75 mm, 3 μm, Portland, OR, USA)이 장착된 HPLC 시스템(Agilent Technologies 1260 Infinity, Santa Clara, CA, USA)을 사용하여 분석하였다. 샘플은 UV 검출기를 사용하여 203 nm에서 측정되었으며, 이동상으로 용매 A (10% acetonitrile)와 용 매 B (90% acetonitrile)를 사용하여 진세노사이드를 40에서 농도 기울기로 분리하였다. 용매(A/B) 비율은 초기 0-3 min에 95/5, 20 min에 90/10, 50 min에 75/25 그리고 60 min에 95/5가 최종 비율
이었다. 유속은 1.3 mL/min, 시료주입량은 5 μL이었다. 사포닌 표 준물질로는 Rb1, Rb2, Rc, Rd, Re, Rf, F2, Rh1 (s), Rh2 (s), Rk1, Rg1, Rg2 (s), Rg2 (r), Rg3 (s) (r), Rg5, compound K (CK)가 각각 사용되었다.
ABTS 및 DPPH 라디칼 제거능 측정
ABTS 라디칼(ABTS+)을 형성시키기 위해, 7 mM ABTS와 2.45 mM 과황산포타슘 용액을 혼합하여 하루 동안 실온, 암소 조건 에서 방치시켰다. 734 nm에서 흡광도 값이 0.7이 되도록 100% 에탄올로 희석하여 ABTS라디칼 용액을 만들었다. 희석된 ABTS+ 용액 150 μL와 농도 별로 희석시킨 SF 50 μL를 30분간 암소에서 반응시킨 뒤 734 nm에서 흡광도를 측정하였다. DPPH 라디칼 제 거능 측정은 메탄올을 사용하여 200 μM의 DPPH 용액을 제조한 후, 517 nm에서 0.7의 흡광도 값이 되도록 희석하였다. 희석된 DPPH 용액 100 μL와 농도 별로 희석시킨 SF 100 μL를 30분간 암소에서 반응시킨 뒤 517 nm에서 흡광도를 측정하였다. 각 농 도별 라디칼 제거능 곡선을 구한 뒤 50%를 제거하는 농도(IC50)
를 산출하였다. 대조군으로는 카테킨(Sigma-Aldrich Co., MO, USA)을 농도별로 사용하였다. 라디칼 제거능은 다음 식에 의해 산출하였다.
ABTS or DPPH radical scavenging activity (%) =(1−sample absorbance)×100 3T3-L1 세포 분화 및 배양 마우스 유래 전지방세포(3T3-L1 preadipocyte)는 소혈청(BS, 10%)과 페니실린-스트렙토마이신(1%), L-글루타민(584 mg/L), 피 루브산소듐(110 mg/L)를 함유한 DMEM을 배지로 37oC, 5% CO 2 조건의 배양기에서 배양하였다. 2일마다 배지를 교환하였고 PBS 용액과 0.05% 트립신-EDTA를 사용하여 세포를 계대하였다. 이 들 세포를 6 well plate에 균등하게 분주하였고, 100% confluent 로부터 2-3일 후 10% FBS, 1% 페니실린-스트렙토마이신, DMEM 에 분화 칵테일용액(1 μM DEX, 0.5 mM IBMX, 1.67 μM insu-lin)을 혼합하여 분화를 유도하였다. 분화는 6일 동안 진행되었으 며 분화시작 2일 째 부터 이틀 동안 인슐린(1.67 μM)과 10% FBS 를 함유한 DMEM으로 교환해 주었고, 그 이후부터는 10% FBS 를 함유한 DMEM으로 2일마다 교환해 주었다. 세포독성 시험 지방세포에 대한 SF의 세포독성 평가는 싸이아졸릴블루 테트 라졸륨 브로민화물(MTT)를 이용하여 측정하였다. 3T3-L1 세포를 96-well plate에 각각 1×104 cells/well을 분주한 뒤, 24시간 동안 배양시켰다. SF를 포함한 배양액을 2일 마다 교체하며 6일간 배 양시켰다. 배양액을 제거한 뒤 MTT 시약(0.5 mg/mL)이 포함된 serum이 없는 배양액으로 1시간 동안 배양기에서 반응시킨 후 생 성된 MTT 포마잔을 DMSO에 녹여, 마이크로플레이트 판독기 (Spectra Max M3, Molecular Devices, Sunnylvale, CA, USA)의 550 nm에서 흡광도를 측정하였다. Oil red O 염색 SF (50, 100, 200μg/mL)를 농도 별로 처리하여 6일 동안 배양 하고 배지를 제거한 후, 10% 포말린으로 세포를 5분간 실온에서 고정시켰다. 3차 증류수로 세척한 뒤 Oil red O 용액을 첨가하여 24시간 동안 지방구 염색을 진행하였다. 염색 후 증류수로 세포 를 세척하고 지방구의 세포 내 축적을 카메라가 장착된 현미경
(Leica DM2500, Wetzlar, Germany)을 이용하여 관찰하고 사진촬 영을 통해 SF가 지방세포 분화에 미치는 영향을 관찰하였다. 이 미지J 프로그램(NIH, MD, USA)을 이용해 염색된 지방을 정량 하였다. ROS 생산양 측정 분화 과정에 따른 지방세포 내 ROS의 생성양을 측정하고자 디클로로플루오로레신 디아세테이트(dichlorofluorescin diacetate, DCFH-DA)와 나이트로 블루 테트라졸륨(nitro blue tetrazolium, NBT) 방법을 이용하였다. DCFH-DA 방법에서는 4일 또는 6 일 동안 SF 또는 vehicle 처리와 함께 분화시킨 3T3-L1에 DCFH-DA (20 µM)를 처리한 뒤 30분간 반응시킨다. 다이메틸설폭사이드 (dimethylsulfoixde, DMSO)를 주입하여 세포막이 파괴되게 하였고, 세포 내 ROS에 의해서 생성된 DCF 포마잔을 형관판 판독기 (excitation 485 nm, emission 535 nm) (Spectra Max M3, Molec-ular Devices)를 이용해 측정하였다. NBT 방법에서는 SF 또는 vehicle와 함께 6일 동안 분화된 3T3-L1 세포를 PBS로 2회 세척 하고 0.2% NBT 용액을 주입하여 CO2배양기에서 90분간 반응 시켜 ROS 생산의 지표인 검푸른 포마잔의 생성량을 확인하였다. 생성된 검푸른 포마잔은 이미지J 프로그램을 이용해 정량화 하였다. 웨스턴 블롯 지방세포의 분화과정 또는 분화 후 단백질가수분해효소와 인 산가수분해효소 억제제(aprotinin, leupeptin, benzamidine, pepstatin, sodium orthovanadate, phenylmethylsulfonyl uoride (PMSF))를 함 유하는 lysis buffer를 이용하여 세포를 용해하여 단백질 추출액을 얻었다. 이 세포 용해액을 10,000×g, 4oC에서 10분간 원심분리 하
여 상층액만 회수하였다. Bicinchoninic acid (BCA)시약을 사용하 여 단백질을 정량한 후 50 µg의 단백질을 10% 또는 12% 아크 릴아마이드에 전기이동 후 플루오린화 폴리바이닐리덴(polyvinylidene uoride, PVDF) 막으로 옮겼다. 5% 탈지분유로 30분간 membrane 을 blocking 후 1차 항체를 넣고(1:1000) 4oC에서 24시간 반응시
켰다. 다음 TBST (tris-buffered saline+0.1% Tween 20)로 세척한 후 2차 항체를 실온에서 2시간 동안 반응시켰다. TBST로 세척 한 후 단백질 검출은 LAS imaging software (Fuji, New York, NY, USA)를 사용하여 화학발광으로 확인하였고. 또한, 각 그룹 별 단백질양은 이미지 J 프로그램을 이용해 정량하였다.
핵과 세포질의 분리
분화시킨 3T3-L1를 PBS로 2회 세척 후 harvest buffer (10 mM N-2-hydroxyethylpiperazine-N-2-ethanesulfonic acid (HEPES), 50 mM 염화소듐, 0.5 M 설탕, 0.1 mM 에틸렌다이아민테트라아세트 산(ethylenediaminetetraacetic acid, EDTA), 0.5% Triton X-100, 1 mM dithiothreitol (DTT), 10 mM 피로인산 테트라소듐, 100 mM 플루오린화소듐, 17.5 mM 글리세로인산, 1 mM PMSF, 4 mg/ mL 아프로티닌(aprotinin), 2 mg/mL 펩스타틴(pepstatin) A를 이용 하여, 세포를 microtube (1.5 mL)에 회수한 후 피펫으로 세포를 용해하였다. 세포 용해액을 120×g, 4oC에서 10분간 원심분리 하
였다. 핵 분획을 위해, cell pellet을 buffer A (10 mM HEPES, 10 mM KCL, 0.1 mM EDTA, 0.1 mM ethylene glycol-bis (2 amino ethylether)-N,N,N’,N-tetraacetic acid (EGTA), 1 mM DTT, 1 mM PMSF, 4 mg/mL aprotinin, 2 mg/mL pepstatin A)로 다시 현탁시 켰다. 120×g, 4oC에서 5분간 원심분리한 후, pellet을 buffer C
(10 mM HEPES, 0.5 M sodium chloride, 0.1 mM EDTA, 0.1 mM EGTA, 0.1% NP-40, 1 mM DTT, 1 mM PMSF, 4 mg/mL
aprotinin, 2 mg/mL pepstatin A)로 4oC에서 10분간 vortex하여 다 시 현탁시켰다. 10000×g, 4oC에서 10분간 원심 분리 한 후 생성 된 상층액을 핵 분획으로 사용 하였다. 세포질 분획을 위해, 세 포 용해물을 120×g, 4oC에서 5분간 원심분리한 후 얻은 상층액 을 10000×g, 4oC에서 15분간 원심분리하고 생성된 상층액을 세 포질 분획으로 사용하였다. 통계분석
모든 실험 결과는 SPSS (Statistical Package for the Social Science, SPSS Inc., Chicago, IL, USA) 프로그램을 이용하여 통 계분석 하였다. 모든 측정항목의 결과는 평균±표준편차로 표시 하였다. 유의성 분석은 one-way ANOVA 검정을 실시하였으며, Duncan’s multiple range test로 유의성은 p<0.05 수준에서 검정하 였다.
결과 및 고찰
세포독성 평가 홍삼 사포닌 분획(SF)의 지방세포에 대한 세포독성을 평가한 결과는 Fig. 1과 같다. 처리한 SF의 농도 200 μg/mg까지는 지방 세포에 유의적인 독성 및 생장저해 효과를 보이지 않은 반면 400 μg/mg 이상의 처리시에는 지방세포에 대해 독성을 보이기 시작 하는 것으로 관찰되었다. 따라서 이후 실험은 세포독성이 관찰되 지 않은 50, 100, 200 μg/mg의 농도에서 진행되었다. 홍삼사포닌 분석 SF의 사포닌 조성을 HPLC로 분석한 결과 전체 진세노사이드 총량은 추출물 mg당 523.26 μg (Table 1)으로 50% 이상이 진세 노사이드로 구성되어 있음을 알 수 있고 그 중 가장 많은 함량 을 차지하는 진세노사이드는 Rb1으로 135.31 μg/mg이었다. 반면, compound K (CK)와 Rh2는 본 분석에서는 검출되지 않았다. Rb1 은 항 염증을 비롯한 다양한 생리활성을 가지고 있는 것으로 알 려지고 있다(Wu 등, 2018; Yuan 등, 2014). Rb1은 쥐(rat)에서 LPS로 유도된 폐의 염증에 대한 억제효과를 발휘하는 것으로 알 려져 있으며(Yuan 등, 2014). 최근 한 논문은 Rb1이 비만 쥐의 뇌에서 leptin에 대한 민감성을 향상시킴을 보고하였다(Wu 등, 2018). SF에는 Rb1이외에도 다양한 진세노사이드가 포함되어 있 기 때문에 본 연구에서 보이는 지방축적억제효과와 라디칼 제거Fig. 1. Effect of saponin fraction (SF) on viability of 3T3-L1 cells. Cells were treated with SF or vehicle (DMSO) for 8 days. MTT assay was performed in triplicate according to manufacturer’s protocol.
능 및 Nrf2 신호전달체계조절능력은 주로 Rb1에 의해서 이루어 진다고 볼 수 없으며 다양하게 조성되어 있는 사포닌들의 조합 에 의한 효과일 가능성이 큰 것으로 판단된다. 따라서 최적화된 사포닌의 조합을 발굴하는 작업이 추후에 이루어 져야 할 것이다. 홍삼사포닌 분획의 지방축적에 대한 영향 SF는 지방분화 시 생성되는 지방 축적의 정도를 농도 의존적 으로 억제하였다(Fig. 2A). 200 μg/mL의 SF를 처리할 때 지방에 대한 Oil red O의 염색이 55.5% 감소하였다(Fig. 2B와 C) 이러 한 SF의 지방축적 억제효과는 지방분화인자들인 PPARγ, C/EBPα 와 이들의 타겟 인자인 FABP4의 단백질의 발현이 감소되는 것 과 일치하였다(Fig. 2B). 이들 지방분화인자들의 단백질 양은 SF 에 의해 농도의존적으로 감소하였는데 높은 농도의 SF에서 이들 의 단백질의 발현은 90%이상 감소를 보였다(Fig. 2C). 이는 SF 가 지방분화인자들의 발현을 억제함으로써 지방분화 및 축적을 억제함을 보여준다. 대부분의 지방분화 및 축적에 대한 연구들에 서 지방분화인자들의 발현이 직접적으로 지방축적에 절대적인 영 향을 미치는 것으로 관찰된다(Kim 등, 2016; Suh 등, 2015). 이 것은 대부분 지방분화인자들의 타겟 유전자들이 지방 형성과 관 련 있는 유전자들이기 때문인 것으로 보인다(Suh 등, 2015). 홍 삼 및 인삼의 사포닌 성분 중에서 특히 Rg3는 지방분화 중 지 방분화인자들 뿐만 아니라(Zhang 등, 2017) 글리세롤 3인산 수소 제거효소(glycerol-3-phosphate dehydrogenase)의 활성과 FABP4의 mRNA의 발현을 억제하는 것이 보고되었으며(Lee 등, 2011), Rg2 도 지방분화관련 유전자들의 발현을 조절하는 것으로 알려져 있 다(Liu 등, 2018). 따라서 홍삼사포닌 중 특정사포닌 성분이 아니 라 다양한 사포닌성분들이 효과를 발휘하는 것으로 판단되며 특 히, 추출물 및 분획에서는 이러한 다양한 사포닌들의 조합에 의 해 지방축적억제 효과가 발휘되는 것으로 판단된다. 홍삼사포닌 분획의 지방분화시기와 초기 지방분화인자에 대한 영향 지방분화는 시기별로 특정 분화 유전자들의 발현이 순차적으 로 다르게 이루어지며 시기별 이들의 발현양상이 궁극적으로 전 체 지방분화에 중요하게 작용한다(Rosen 등, 2000). SF의 분화시 기별 효과를 측정한 결과는 Fig. 3에 나타내었다. 분화 초기에만 SF를 처리한 군에서 분화기간 종료 후 축적된 지방의 양이 대조 군에 비해 55%의 감소효과를 보였고 전체 분화시기를 모두 처 리한 군에서는 76%의 감소효과를 보였다(Fig. 3C). 하지만 분화 후기에만 처리한 군에서는 단지 10%정도의 감소효과를 보임으 로써 SF는 주로 지방 분화 초기에 관여하여 지방분화를 억제하 는 것으로 관찰되었다(Fig. 3). C/EBPβ/δ와 KLF2 등의 지방분화 초기인자들은 나중에 PPARγ나 C/EBPα 등의 지방분화 후기인자 들의 발현에 영향을 미침으로서 전체적으로 지방분화 및 축적에 영향을 미치는 것으로 보고되고 있다(Rosen 등, 2000). 본 연구 에서 이들 초기인자들의 단백질 발현양에 대한 SF의 효과를 측 정해 본 결과 C/EBPβ는 억제되었고(90%), KLF2의 단백질은 증 가(~30%)되는 것이 관찰되었다(Fig. 4). 이 결과는 SF가 초기 지 방분화인자인 C/EBPβ는 억제하고 초기 항지방분화인자인 KLF2 는 활성화시키는 조절 작용을 통해서 전체적인 분화 및 지방축 적을 억제하는 것으로 판단되었다. 몇몇 연구들에서도 식품 유래 파이토케미칼류들이 주로 지방분화 초기부터 지방분화초기인자 들을 조절함으로써 지방축적억제 효과를 발휘하는 것으로 보고 되고 있다(Choi 등, 2012; Choi 등, 2014; Suh 등, 2015).
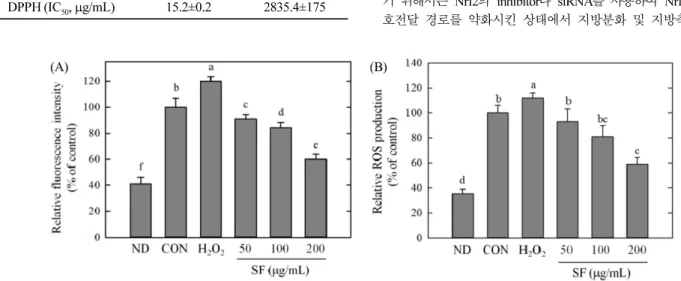
홍삼사포닌 분획의 ROS 생성에 대한 효과
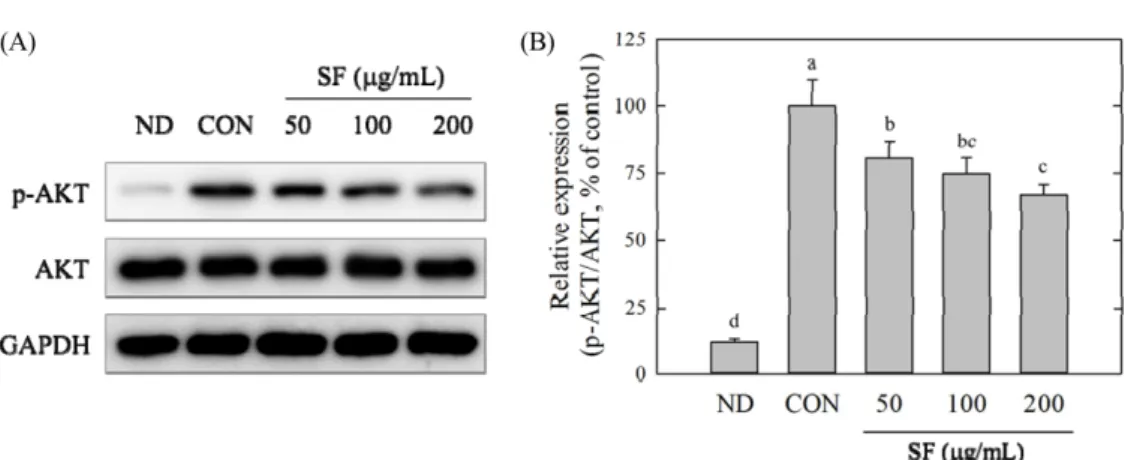
지방분화 기간 동안 지방의 축적은 과도한 ROS의 생성이 동 반되는 것으로 알려져 있으며(Furukawa 등, 2017; Lee 등, 2009). 이러한 ROS의 생성은 지방분화의 초기에 세포의 클론 확장 유 사분열(mitotic clonal expansion)을 가속화함으로써 분화를 촉진하 는 것으로 보고되고 있다(Lee 등, 2009). 또한 비만 및 비만 관 련 질병들에서 ROS에 의한 산화스트레스가 항진되어 있는 것으 로 알려지고 있다(Furukawa 등, 2017). 본 연구에서는 지방분화 시 생성되는 ROS의 생성에 대한 SF의 효과를 DCFH-DA법(Fig. 5A)과 NBT법(Fig. 5B)을 통해 측정하였으며 SF가 효과적으로 지 방분화 동안 생성되는 ROS의 생성을 억제하는 것으로 관찰되었 다(Fig. 5). 온전히 분화된 대조군에 비해 ROS의 한 형태인 과산 화수소를 처리하였을 때 오히려 지방의 축적이 15~20% 가량 증 가하였다(Fig. 5). 이는 세포 내 ROS의 증가가 지방축적을 촉진 한 것으로 보인다. 반면, SF를 처리하였을 때 농도의존적으로 ROS의 생성량이 감소하였으며 200 μg/mL의 SF를 처리하였을 때 대조군에 비해 40% 감소하였다. 이는 SF에 의한 지방축적 억제 효과가 ROS생성 억제능과 밀접하게 관련 있음을 보여준다. SF 에 의한 세포 내 ROS의 감소가 직접적인 ROS의 제거 때문인지 아니면 세포 내 ROS생성 관련 메커니즘 기구의 조절효과 때문 인지 알기 위해 SF가 ABTS와 DPPH라디칼에 대해 어느 정도의 제거작용이 있는지 알아본 결과(Table 2), ABTS와 DPPH라디칼 에 대한 제거작용을 나타내는 IC50값이 각각 1053.6±96, 2835.4± 175μg/mL으로 대조군인 카테킨보다 훨씬 높은 값을 나타내어 카 테킨보다 직접적 라디칼 제거 작용은 크지 않은 것으로 관찰되 었다. 따라서 SF의 라디칼에 대한 직접적인 제거작용으로 인해 세포 내 ROS의 생성이 억제되었다고 할 수 없다. 홍삼 사포닌 분획의 Nrf2/Keap1 pathway에 대한 영향 SF의 지방축적억제효과와 ROS 조절효과가 확인됨에 따라 SF 에 의한 세포 내 ROS의 조절작용이 지방축적억제효과와 밀접한 관련이 있는 것으로 판단되었다. 그러나 SF의 라디칼 제거 작용 을 측정한 결과, 홍삼 사포닌분획은 직접적인 ROS 제거보다는 Table 1. Ginsenoside composition from saponin fraction
No. Standards Ginsenoside (μg/mg)
1 Rb1 135.31 2 Rb2 44.03 3 Rc 58.71 4 Rd 18.14 5 Re 42.76 6 Rf 39.14 7 Rh1(s)+Rg2(s) 10.01 8 Rh2(s)+Rh2(r) -* 9 Rg1 32.15 10 Rg2(r) 17.03 11 Rg3(s) 36.58 12 Rg3(r) 15.08 13 Rg5 20.42 14 Rk1 21.22 15 CK -* 16 F2 32.69 Total 523.26 * not detected
다른 간접적인 방법으로 세포 내 ROS를 조절하는 것으로 판단 된다. 세포 내 ROS를 조절하는 대표적인 시스템으로 Nrf2/Keap1 신호전달체계를 꼽을 수 있으며(Taguchi 등, 2011), 이 신호전달 체계에 대한 SF의 효과를 측정하였다. Nrf2는 충분히 분화된 지 방세포(대조군)에서는 분화되지 않은 군에 비해 그 수준이 현저 하게 낮은 것을 확인할 수 있으며 SF를 처리함에 따라 Nrf2 단 백질의 세포 내 발현 수준이 증가하였다(Fig. 6A). 200 μg/mL 농 도의 SF는 대조군에 비해 2배 이상 증가하였다(Fig. 6B). 또한 Nrf2의 타겟 단백질인 hemoxygenase-1 (HO-1)와 NADPH dehy-drogenase 1 (NQO1)의 양도 SF에 의해 농도의존적으로 증가하였 으며 200 μg/mL의 농도에서 HO-1와 NQO1 단백질 양이 대조군 에 비해 각각 3.3배, 1.8배 증가하였다(Fig. 6A와 B). 반면 Nrf2 의 복합체 파트너인 Keap1은 홍삼 사포닌 분획에 의해 감소하는 것으로 나타나 Nrf2의 양과 대조를 이루었다 (Fig. 6A). Nrf2는 전사인자로서 핵 내로 들어가 ROS 생산에 영향을 미치는 유전 자들의 발현을 조절하는 것으로 알려져 있기 때문에 Nrf2의 핵 내로의 진입이 중요하다(Taguchi 등, 2011). 따라서 Nrf2의 핵 내 로의 진입에 대한 SF의 효과를 측정하기 위해 세포 추출물을 핵 분획과 세포질 분획으로 분리하여 Nrf2의 양을 측정하였다(Fig. 6C와 D). 분화된 지방세포에서는 대부분의 Nrf2가 핵 내 보다는 세포질에 존재하는 것으로 관찰되었고 SF을 처리함에 따라 핵 내 존재하는 Nrf2의 양이 증가하여 높은 농도의 SF (200 μg/mL) 을 처리할 때 대조군에 비해 Nrf2의 양이 3배 가량 증가하였다 (Fig. 6D). 이러한 결과는 SF가 Nrf2의 핵 내 진입을 촉진함으로 써 타겟 단백질인 HO-1, NQO1의 양을 증가시킴을 보여준다. Nrf2의 핵 내로의 이동증가는 Nrf2/Keap1 신호전달 체계가 활성 화 됨을 의미하며 몇몇 연구들에서 Nrf2가 상위 신호전달 시스 템에 의해 인산화 되어 핵 내 진입이 이루어짐을 보고 하였다 (Chen 등, 2012; Wang 등, 2008). 따라서 SF가 Nrf2의 상위 신호 전달 체계를 활성화시키는지를 알아보기 위해 AKT에 대한 SF의 활성을 분석하였다(Fig. 7). SF는 AKT의 인산화를 약간 억제함 으로써 오히려 그 활성을 감소시키는 것으로 관찰되었다(Fig. 7). Fig. 2. Effect of saponin fraction (SF) on lipid accumulation during adipocyte differentiation. After differentiation with the SF or vehicle for 6 days, lipid accumulation levels of cells were visualized by Oil red O staining and quantified by ImageJ software (A). Total protein was obtained from the cell, and expression levels of PPARγ, C/EBPα and FABP4 protein were analyzed by Western blot (B) and quantified by ImageJ software (C). Data are expressed as means±SE. Experiments were performed in three replicates.
이러한 결과는 SF가 AKT의활성을 통해서 Nrf2/Keap1 신호전달 경로를 활성화 시키는 것이 아니라 SF가 직접 Nrf2 시스템을 활 성화 시키거나 AKT 이외의 다른 신호전달 시스템을 통해서 Nrf2 의활성화를 이룰 가능성이 있음을 보여준다. 따라서 Nrf2의 상위 신호전달 시스템 또는 직접적인 SF와 Nrf2의 관계에 대해서는 추후 연구가 필요 할 것으로 판단된다. 많은 연구들에서 HO-1는 ROS의 생성을 억제함으로써 산화스트레스를 감소시키는 것으로 보고되고 있어(Tang 등, 2014; Zhong 등, 2013) SF에 의한 ROS 조절이 지방축적 억제효과의 유력한 메커니즘으로 판단된다. 더 나아가, Nrf2는 HO-1뿐만 아니라 초과산화물제거효소(SOD), 카 탈레이스, 글루타싸이온 과산화효소(Gpx)와 같은 ROS를 조절하 는 산화 방지 효소들의 발현을 촉진시키는 것으로 알려져(Zhang 등, 2013) 있으므로, 이들 산화방지 효소들의 활성화가 복합적으 로 SF의 ROS조절에 크게 기여하는 것으로 보여진다. 따라서 SF 에 의한 Nrf2 활성화가 지방분화 시 과도하게 생산되는 세포 내 ROS를 억제하여 지방분화 및 지방축적을 억제하는 것으로 판단 된다. 지방세포분화와 Nrf2/Keap1 신호전달체계와의 관계에 대해 서 비교적 많은 연구가 이루어져 왔다. Nrf2 유전자 제거가 지방 분화를 감소시킬 뿐만 아니라 지방분화인자들의 발현을 감소시 켰다는 보고(Pi 등, 2010)가 있는 반면, Shin 등(2007)은 Nrf2가 제거된 마우스 태아모세포에서 지방분화가 크게 촉진되었다는 연 구결과도 보고되고 있다. 이러한 서로 상반되는 결과들이 충돌하 고 있는 가운데 많은 연구들에서 Nrf2를 활성화 시키는 파이토 케미칼들의 지방분화 및 지방축적 억제효과가 보고되었다(Gaikwad 등, 2001; Lee와 Do, 2001; Panchal 등, 2012). 특히 설포라판은 Nrf2와 타겟단백질인 NQO1을 활성화함으로써 지방축적을 억제 Fig. 3. Effect of saponin fraction (SF) on adipogenic stages. Cells were differentiated with treatment of SF for indicated times. After 6 days, effect of SF (200μg/mL) on lipid accumulation were visualized by Oil red O staining (A) and quantified by the ImageJ software (B). FBS: differentiation medium, MDI: hormonal cocktail (1 µM DEX, 0.5 mM IBMX, 1.67 µM insulin), INS: insulin, BSM: growth medium, DMSO: dimethyl sulfoxide. Data are expressed as means±SE. Experiments were performed in three replicates.
Fig. 4. Effect of saponin fraction (SF) on early adipogenic factors. SF (50, 100 and 200μg/mL) was treated for 4 h in the 3T3-L1 differentiation, and cells were harvested for protein extraction. Expression levels of C/EBPβ and KLF2 proteins were analyzed by Western blot (A) and quantified using ImageJ software (B). Data are expressed as means±SE. Experiments were performed in three replicates.
하는 것으로 알려져 있다(Gaikwad 등, 2001). 본 연구에서도 SF 에 의한 Nrf2/Keap1 신호전달체계의 활성화가 지방분화 및 지방 축적을 억제시키는 것으로 판단된다. 하지만 이를 확실히 검증하 기 위해서는 Nrf2의 inhibitor나 siRNA를 사용하여 Nrf2/Keap1 신 호전달 경로를 약화시킨 상태에서 지방분화 및 지방축적을 측정
Fig. 5. Effect of saponin fraction (SF) on the production of ROS in 3T3-L1 cells. Cells were treated with different concentration of SF or 100μM hydrogen peroxide for 6 days and the cells were incubated with 20 μM DCFH-DA for 30 min. The fluorescence intensity was detected using fluorescence plate reader at excitation 485nm and emission 535 nm (A). After differentiation for 6 days, production of ROS was also determined by NBT assay and quantified using ImageJ software (B). Data are expressed as means±SE. Experiments were performed in three replicates.
Fig. 6. Effect of saponin fraction (SF) on the protein expression level of Nrf2, Keap1, HO-1 and NQO1, and Nrf2 nuclear translocation in 3T3-L1 cells. Cells were differentiated and treated with different concentrations of SF for 4 days. Expression levels of Nrf2, Keap1, HO-1 and NQO1 proteins were analyzed by Western blot (A) and quantified using ImageJ software (B). Nuclear and cytosolic fractions were prepared as described in methods, and abundances of nuclear and cytosolic Nrf2 proteins were analyzed by Western blot (C) and quantified using ImageJ software (D). Data are expressed as means±SE. Experiments were performed in three replicates.
Table 2. Radical scavenging activity of saponin fraction
Item Catechin SF
ABTS (IC50, μg/mL) 19.3±0.16 1053.6±96 DPPH (IC50, μg/mL) 15.2±0.2 2835.4±175
하는 것이 필요하다. 특히 Nrf2에 대해 특이성이 상대적으로 좋 은 siRNA를 사용하여 검증하는 것이 바람직하다고 할 수 있으 며, 이는 다른 신호전달체계에 대한 분석과 함께 추후 확인 가능 할 것으로 판단된다.
요
약
본 연구에서는 홍삼 사포닌 분획(SF)으로부터 진세노사이드의 조성을 분석하고 지방세포의 분화 및 지방축적에 대한 효과를 측 정하였다. SF는 지방분화인자인 PPARγ, C/EBPα의 단백질 양을 억제함으로써 지방분화 동안 효과적으로 지방축적을 억제하였으 며 주로 지방분화 초기시점부터 지방분화 초기인자인 C/EBPβ, KLF2의 조절작용을 통해 지방축적을 억제하는 것으로 관찰되었 다. SF는 또한 지방분화 동안 생성되는 ROS의 생성을 효과적으 로 억제하였는데 이는 SF가 산화방지 시스템인 Nrf2/Keap1 경로 를 활성화하기 때문으로 판단되며 특히 Nrf2의 핵 내로의 진입 을 활성화 함으로써 Nrf2의 타겟 산화방지 분자들인 HO-1, NQO1 의 발현을 촉진하였다. 이는 지방분화 동안 SF의 지방축적 억제 효과가 Nrf2의 활성화와 밀접하게 관련이 있음을 보여준다.감사의 글
시료를 제공해주신 고려인삼학회에 감사드리며, 본 연구는 서 울여자대학교 교내 학술연구비(2018)로부터 지원 받았음.References
Attele AS, Wu JA, Yuan CS. Ginseng pharmacology: multiple con-stituents and multiple actions. Biochem. Pharmacol. 58: 1685-1693 (1995)
Chang H. Effect of processing methods on the saponin contents of Panax ginseng leaf-tea. J. Food Sci. Nut.16: 46-53 (2003)
Chang YS, Chang YH, Sung JH. The effect of ginseng and caffeine products on the antioxidative activities of mouse kidney. J. Gin-seng Res. 30: 15-21 (2006)
Chen HH, Chen YT, Huang YW, Tsai HJ, Kuo CC. 4-Ketopinores-inol, a novel naturally occurring ARE activator, induces the Nrf2/ HO-1 axis and protects against oxidative stress-induced cell injury via activation of PI3K/AKT signaling. Free Radic. Biol. Med. 52: 1054-1066 (2012)
Choi HS, Jeon HJ, Lee OH, Lee BY. Indole-3-carbinol, a vegetable
phytochemical, inhibits adipogenesis by regulating cell cycle and AMPKα signaling. Biochimie. 104: 127-136 (2014)
Choi KM, Lee YS, Sin DM, Lee S, Lee MK, Lee YM, Hong JT, Yun YP, Yoo HS. Sulforaphane inhibits mitotic clonal expansion during adipogenesis through cell cycle arrest. Obesity. 20: 1365-1371 (2012)
Furukawa S, Fujita T, Shimabukuro M, Iwaki M, Yamada Y, Naka-jima Y, Nakayama O, Makishima M, Matsuda M, Shimomura I. Increased oxidative stress in obesity and its impact on metabolic syndrome. J. Clin. Invest. 114: 1752-1761 (2017)
Gaikwad A, Long DJ, Stringer JL, Jaiswal AK. In Vivo Role of NAD (P) H: quinone oxidoreductase1 (NQO1) in the regulation of intracellular redox state and accumulation of abdominal adi-pose tissue. J. Biol. Chem. 276: 22559-22564 (2001)
Halliwell B, Gutteridge JM, Cross CE. Free radicals, antioxidants, and human disease: where are we now?. J. Lab.Clin. Med. 119: 598-620 (1992)
He HJ, Wang GY, Gao Y, Ling WH, Yu ZW, Jin TR. Curcumin attenuates Nrf2 signaling defect, oxidative stress in muscle and glucose intolerance in high fat diet-fed mice. World J. Diabetes 3: 94-104 (2012)
Kersten S. Mechanisms of nutritional and hormonal regulation of lipogenesis. EMBO Reports. 2: 282-286 (2001)
Keum YS, Park KK, Lee JM, Chun KS, Park JH, Lee SK, Kwon HJ, Surh YJ. Antioxidant and anti-tumor promoting activities of the methanol extract of heat-processed ginseng. Cancer Letters. 150: 41-48 (2000)
Kim DH, Kwak KH, Lee KJ, Kim SJ, Shin YC, Chun BG, Shin, KH. Effects of Korea red ginseng total saponin on repeated unpredictable stress-induced changes of proliferation of neural progenitor cells and BDNF mRNA expression in adult rat hip-pocampus. J. Ginseng Res. 28: 94-103 (2004).
Kim JH, Kang S, Jung YN, Choi H-S. Cholecalciferol inhibits lipid accumulation by regulating early adipogenesis in cultured adipo-cytes and zebrafish. Biochem. Biophys. Res. Commun. 469: 646-653 (2016)
Kim JY, Park JY, Kang HJ, Kim OY, Lee JH. Beneficial effects of Korean red ginseng on lymphocyte DNA damage, antioxidant enzyme activity, and LDL oxidation in healthy participants: a randomized, double-blind, placebo-controlled trial. Nutr. J.11: 47 (2012)
Konno C, Hikino H. Isolation and hypoglycemic activity of panaxans M, N, O and P, glycans of Panax ginseng roots. Int. J. Crude Drug Res. 25: 53-56 (1987)
Lee H, Lee YJ, Choi H, Ko EH, Kim JW. Reactive oxygen species facilitate adipocyte differentiation by accelerating mitotic clonal expansion. J. Biol. Chem. 284: 10601-10609 (2009)
Lee JW, Do JH. Antioxidative activity of ethanol extraction fraction from the Korean red tail ginseng. Kor. J. Food Sci. Technol. 33: 497-500 (2001).
Fig. 7. Effect of saponin fraction (SF) on the protein expression level of AKT, P-AKT in 3T3-L1 cells. SF (50, 100 and 200μg/mL) was treated for 3 h in the 3T3-L1 differentiation, and cells were harvested for protein extraction. Expression levels of AKT and P-AKT proteins were analyzed by Western blot (A) and quantified using ImageJ software (B). Data are expressed as means±SE. Experiments were performed in three replicates.
Lee M, Kim I, Kim C, Kim Y. Effects of ginsenoside Rg3 on adipo-cyte fatty acid binding protein mRNA expression and glycerol-3-phosphate dehydrogenase activity during adipocytes differentia-tion. Kor. J. Lipidol. 21: 67-75 (2011)
Liu H, Wang J, Liu M, Zhao H, Yaqoob S, Zheng M, Cai D, Liu J. Antiobesity effects of ginsenoside Rg1 on 3T3-L1 preadipocytes and high fat diet-induced obese mice mediated by AMPK. Nutri-ents 10: 830-844 (2018)
Lowe CE, O'Rahilly S, Rochford JJ. Adipogenesis at a glance. J. Cell Sci. 124: 2681-2686 (2011)
Nam KY. The comparative understanding between red ginseng and white ginsengs, processed ginsengs (Panax ginseng CA Meyer). J. Ginseng Res. 29: 1-18 (2005)
Oakley FD, Abbott D, Li Q, Engelhardt JF. Signaling components of redox active endosomes: the redoxosomes. Antioxid. Redox Sig-nal. 11: 1313-1333 (2009)
Panchal SK, Poudyal H, Brown L. Quercetin ameliorates cardiovas-cular, hepatic, and metabolic changes in diet-induced metabolic syndrome in rats. J. Nutr. 142: 1026-1032 (2012)
Pi J, Leung L, Xue P, Wang W, Hou Y, Liu D,Yehuda-Shnaidman E, Lee, C, Lau J, Kurtz T W, Chan JY. Deficiency in the nuclear factor E2-related factor 2 transcription factor results in impaired adipogenesis and protects against diet-induced obesity. J. Biol. Chem. 285: 9292-9300 (2010)
Rosen ED, Walkey CJ, Puigserver P, Spiegelman BM. Transcriptional regulation of adipogenesis. Genes Dev. 14: 1293-1307 (2000) Shin S, Wakabayashi N, Misra V, Biswal S, Lee GH, Agoston ES,
Yamamoto M, Kensler TW. NRF2 modulates aryl hydrocarbon receptor signaling: influence on adipogenesis. Mol. Cell. Biol. 27: 7188-7197 (2007)
Spiegelman BM, Choy L, Hotamisligil GS, Graves RA, Tontonoz P. Regulation of adipocyte gene expression in differentiation and syndromes of obesity/diabetes. J. Biol. Chem. 268: 6823-6826 (1993)
Suh HJ, Cho SY, Kim EY, Choi HS. Blockade of lipid accumulation
by silibinin in adipocytes and zebrafish. Chem. Biol. Interact. 227: 53-62 (2015)
Taguchi K, Motohashi H, Yamamoto M. Molecular mechanisms of the Keap1–Nrf2 pathway in stress response and cancer evolution. Genes Cells. 16: 123-140 (2011)
Tang W, Yan J, Wang T, Xia X, Zhuang X, Hong K, Li R, Liu P, Jiang H, Qiao J. Up-regulation of heme oxygenase-1 expression modulates reactive oxygen species level during the cryopreserva-tion of human seminiferous tubules. Fertil. Steril. 102: 974-980 (2014)
Wang L, Chen Y, Sternberg P, Cai J. Essential roles of the PI3 kinase/Akt pathway in regulating Nrf2-dependent antioxidant functions in the RPE. Invest. Ophthalmol. Vis. Sci. 49: 1671-1678 (2008)
Wu Y, Huang XF, Bell C, Yu Y. Ginsenoside Rb1 improves leptin sensitivity in the prefrontal cortex in obese mice. CNS Neurosci. Ther. 24: 98-107 (2018)
Yoon S, Joo C. Study on the preventive effect of ginsenosides against hypercholesterolemia and its mechanism. Kor. J. Ginseng Sci. 17: 1-12 (1993)
Yuan Q, Jiang YW, Ma TT, Fang QH, Pan L. Attenuating effect of Ginsenoside Rb1 on LPS-induced lung injury in rats. J. Inflam-mation. 11: 40 (2014)
Zhang L, Zhang L, Wang X, Si H. Anti-adipogenic effects and mechanisms of ginsenoside Rg3 in pre-adipocytes and obese mice. Front. Pharmacol. 8: 113 (2017)
Zhang M, An C, Gao Y, Leak RK, Chen J, Zhang F. Emerging roles of Nrf2 and phase II antioxidant enzymes in neuroprotection. Prog. Neurobiol.100: 30-47 (2013)
Zhong Y, Liu T, Lai W, Tan Y, Tian D, Guo Z. Heme oxygenase-1-mediated reactive oxygen species reduction is involved in the inhibitory effect of curcumin on lipopolysaccharide-induced monocyte chemoattractant protein-1 production in RAW264.7 macrophages. Molecular Med. Reports. 7: 242-246 (2013)