472
서 론우리나라 연안에서
Cochlodinium polykrikoides
에 의한 적 조는1995
년도 이후로 거의 매년 발생하고 있으며 연안 양식 어업의 생산성을 저해시키고 있다. 특히1995
년도에는 약764
억원의 대규모 수산피해를 일으켰고 이후 거의 매년 발생하였으며 이로 인한 수산피해를 최소화시키기 위하여 정부 및 어업 인은 연안 가두리양식장 주변해역에 적조가 고밀도로 유입 시 적조생물 구제효율이 우수한 선별된 황토를 살포요령에 따라 살포하고 있다.
황토는 암석이 화학적 풍화작용에 의하여 변질되어 토양화 된 풍화잔류토
(Hwang, 1997; Hwang et al., 2000)
이며 우리Article history;
Received 14 June 2012; Revised 2 August 2012; Accepted 18 September 2012
*Corresponding author: Tel: +82. 55. 640. 4771 Fax: +82. 55. 641. 2036 E-mail address: [email protected]
Kor J Fish Aquat Sci 45(5) 472-479, October 2012 http://dx.doi.org/10.5657/KFAS.2012.0472 pISSN:0374-8111
ⓒ The Korean Society of Fishereis and Aquatic Science. All rights reserved
해양퇴적층에서 적조생물(
Cochlodinium polykrikoides
)의 혐기성 분해과정 중 황토가 휘발성 지방산 생성에 미치는 영향박영태 * . 이창규 . 박태규 . 이윤
1. 배헌민
2Effects of Yellow Clay on the Production of Volatile Fatty Acids during the Anaerobic Decomposition of the Red Tide Dinoflagellate
Cochlodinium polykrikoides in Marine Sediments
Southeast Sea Fisheries Research Institute, National Fisheries Research & Development Institute, Tongyeong 650-943, Korea
1
West Sea Fisheries Research Institute, National Fisheries Research & Development Institute, Incheon 400-420, Korea
2
Haema Co. LTD, Busan 612-600, Korea
The formation of volatile fatty acids(VFAs) and changes in pH, oxidation and reduction potential(Eh) and acid volatile sulfide(AVS) with the addition of yellow clay were investigated using microcosm systems to examine the effects of yel- low clay dispersion on the anaerobic decomposition of Cochlodinium polykrikoides in marine sediments. The acetate concentration reached a maximum by day 4 and was 1.2-1.8 fold less in the sample treated with yellow clay compared to the untreated sample (224-270 vs. 333 uM). The formate concentration reached a maximum by day 1 and was 1.3-2.8 fold less in the sample treated with yellow clay compared to the untreated sample (202-439 vs. 563 uM). The propionate concentration reached a maximum by day 2 and was 1.5-1.8 fold less in the sample treated with yellow clay compared to the untreated sample (32.6 vs. 57.2 uM). After the amounts of acetate, formate and propionate peaked the levels dropped dramatically due to the utilization by sulfate reducing bacteria. The Eh of the samples treated with yellow clay was similar to the untreated sample on day 0 but was higher in the sample treated with yellow clay(140-206 mV) from days 4 to 17. AVS started to form on day 3 and this was sustained until day 6, and 1.2-2.2 fold less was produced in the sample treated with yellow clay compared to the untreated sample (40.2-69.3 vs. 83.2-93.8 mg/L).
Accordingly, during the anaerobic decomposition of C. polykrikoides in marine sediments, yellow clay dispersal seems to suppress the reduction state of Eh and the formation of volatile fatty acids(acetate, formate and propionate) used as an energy source by sulfate reducing bacteria, indicating that this process controls the production of hydrogen sulfide that negatively affects marine organisms and the marine sediment environment.
Key words: Cochlodinium polykrikoides, Yellow clay, Volatile fatty acids, Sulfate reducing bacteria, Acid volatile sul- fide, Eh
Young-Tae Park, Chang-Kyu Lee, Tae-Gyu Park, Yoon Lee1 and Heon-Meen Bae2
국립수산과학원 남동해수산연구소, 1국립수산과학원 서해수산연구소, 2(주) 해마
나라의 가까운 산에서 쉽게 볼 수 있는 천연물질로 원적외선 복 사와 유해물질 흡착
-
응집-
침전으로 오래 전부터 우리의 건강 과 환경에 다양하게 이용되고 있다. 우리생활과 밀접한 관계 가 있는 황토는 여러 가지 적조구제물질 중 실제현장에 적용 할 수 있는 우수한 경제적인 방법으로 알려져 있으며(Na et al., 1996; Choi et al., 1998; Kim, 2000; Shirota 1976, 1989; Yu
2004)
, 강우로 인하여 육상으로부터 연안으로 유입되는 천연물질이다. 그러나, 황토가 비록 천연물질이지만, 적조가 발생 하는 수온이 높은 하계에 단기간 집중적으로 살포되므로 해양 생태계에 미치는 영향이 우려되어 황토살포가 수질 및 양식생 물
(Na et al., 1996; Choi et al., 1998; Kim et al., 2002; Bae et al.,1998),
이매패(Seo et al., 2008),
해양 퇴적환경(Choi et al., 1998),
연안 저서생태계에 미치는 영향(Park and Lee, 2006)
등에 관하여 다양하게 조사되어 졌다.수계에 발생한 적조생물에 황토를 살포하면 일부는 수중에서 분해되거나, 혹은 황토입자에 의하여 퇴적층으로 이동되어 분 해작용을 받게 된다. 연안역의 퇴적물 표층에서는 유기물분해 가 활발하게 일어나며
(Jørgensen et al., 1990),
수계에서 생성 된 유기물의 약20-60
%가 분해되고(Jørgensen, 1983)
그 과 정을 통하여 수괴의 생지화학적 순환에 중요한 역할을 하게 된 다. 그러나, 연안역 퇴적층은 혐기성 상태로 되기 쉬우며, 표 층에 퇴적된 유기물의 약50
% 이상이 혐기성세균인 황산염환 원세균에 의하여 분해되어(Jørgensen, 1982),
최종산물로 해 양환경에 악 영향을 미치는 황화수소를 생성하게 된다. 이들 황산염환원세균이 이용하는 에너지원은 주로 휘발성 지방산(volatile fatty acid : VFAs),
아미노산, hydrogen
등으로 다양 하지만(Sørensen et al., 1981; Winfrey and ward, 1983),
이 중 휘발성 지방산이 가장 중요한 기질로 알려져 있다(Mueller- Harvey and Parkes, 1987).
휘발성 지방산은 퇴적층에서 유기 물의 혐기적 분해과정 중 발효세균에 의하여 생산되는 중간대 사물(Kondo et. al., 1990a)
로 황산염이 풍부한 해양환경에서 는 대부분 황산염환원세균에 의하여 이용되어 진다(Canfield et al., 2005).
본 연구는
C. polykrikoides
적조발생 시 살포한 황토가 해 양 퇴적물 표층으로 침강된 적조생물의 혐기적 분해과정에서 휘발성 지방산의 생성과pH,
산화환원전위(Eh)
와 산휘발성 황화물(acid volatile sulfide : AVS)
의 변동에 미치는 영향을microcosm
실험계를 이용하여 간접적으로 조사한 결과이다.재료 및 방법
Microcosm 실험계 구성
Microcosm
실험계 구성을 위한 퇴적물과 해수는2008
년8
월 에 남해 연안역에서 표층해수와 개량형Ekman
채니기로 퇴적 물을 채취하여 표층부(
상부1 cm)
를 시료로 이용하였으며, 채 취된 시료는ice-box
에 보관하여 연구실로 옮겨 처리하였다.황토는
C. polykrikoides
실내구제효율(3,000 cells/mL
밀도의C. polykrikoides
배양액1 L
에 황토10 g
투여10
분 후의 적조 생물 구제효율)이80
% 이상을 보여준 남해와 완도주변에서 채 취된 것을 사용하였다. 모든 실험계는250 mL
삼각비이커로3
개의 반복구로 구성하였으며, 대조구는 황토미첨가 실험계로 퇴적물15 g
을, 황토첨가 실험계는 퇴적물과 황토의 비율을 습 윤/건조무게로15 g:15 g (1:1)
과15 g:30 g (1:2)
으로 하였다.또한, 모든 실험계에 대한 황산염환원세균에 의한 휘발성 지 방산의 이용을 파악하기 위하여, 황산염환원세균을 선택적으 로 억제
(Oremland and Taylor, 1978)
하는sodium molybdate (
최종농도20 mM)
를 첨가 혹은 미첨가 실험계로 각각 구성하 였다.F/2
배지(Guillard and Ryther, 1962)
로 대량 배양한C.
polykrikoides
를plankton net
로 농축하여 각 실험계에 균일하 게 첨가하였으며 유기물 보완으로glucose
와pepton
을 최종농 도가 각각0.001
%로 되도록 첨가 후 현장해수를 상부까지 가 득 채우고 혐기적 조건이 되도록 실리콘마개로 밀봉 하였다.각 실험계의 유기물의 농도는
Kim et al. (2006)
과Bisutiiet al.
(2004)
의 방법으로CHN analyzer(EA1112)
로 분석하였으며 각 실험계의TOC(total organic carbon)
와TON(total organic nitrogen)
은 각각14.5-15.5 mg/mL, 1.8-1.9 mg/mL
범위였 다. 모든 실험계는24
℃의 암소에서 배양하였으며 황토와 퇴 적물이 잘 혼합되도록 약하게 진탕 배양을 하며 경시적으로 시 료를 채취하였다. 퇴적물과 해수가 균질하도록 충분히 흔든 후 혐기적으로 시료를 채취하여 분석하였다.분석
pH
와Eh
는 각각 휴대용pH meter (Hanna HI99121)
와Eh meter (TOA RM-209)
로 측정하였으며 산휘발성 황화물은 시 료채취 후 검지관으로 바로 측정하였다.휘발성 지방산
(acetate, butyrate, formate, lactate, propio- nate, succinate)
은Mueller-Harvey (1987)
의 방법에 의하여HPLC(Shimadzu JP/LC-20A)
를 이용하여ODS
칼럼(5
μm;4.6×250 mm; Restec)
으로 분리하여UV
검출기(Shimadzu SPD-M20A, 400 nm)
로 측정하였다. 이동상은27
%aceto- nitrile
을1.2 mL/min
로 하였으며 컬럼 내부온도는24±2
℃를 유지하였다.Sodinium molybdate
를 첨가한 실험계는Banat et al. (1981)
의 방법으로 전처리 후 휘발성지방산을 유도화하 여 분석하였으며, 황산염화원세균에 의하여 이용되어진 휘발 성지방산은sodium molybdate
를 첨가한 실험계와 미첨가 실 험계간의 농도 차로 구하였다.결과 및 고찰
분해과정 중 환경인자 변동
분해과정 중
pH
의 변동을Fig. 1
에 나타내었으며, 각 실험계 의 초기pH
는 황토 첨가 실험계는7.1-7.2
의 범위로 황토미첨가 실험계
7.6±0.05
에 비하여 약간 낮았으며, 적조배양액에 황 토첨가 시pH
의 감소를 보여준Kim (2000)
의 결과와 일치하 였다. 또한 채취 지역이 다른34
개 황토의pH
가4.3-6.2
의 넓 은 범위로 산성을 보인Hwang et al. (2000)
의 연구결과를 고 려하면, 황토첨가 시pH
의 하락은 황토자체의 산성으로 인한 영향으로 판단된다. 배양4
일째부터 모든 실험계의pH
는 조금 씩 증가하여6
일째부터 배양17
일까지7.5-7.8
범위를 유지하 였으며 황토첨가 실험계가 미첨가 실험계에 비하여 약간 낮거 나 유사하였다. 또한, 완도 황토는 첨가비율이 높을수록 약간 낮은pH
를 보였으나 남해 황토는 첨가 비율에 따라 뚜렷한 차 이가 없었다산화환원전위
(Eh)
는 산소가 존재하는 산화적 환경에서는 양 의 값을, 산소가 결핍된 환원적 환경에서는 음의 값을 나타낸 다(Shiozawa, 1979)
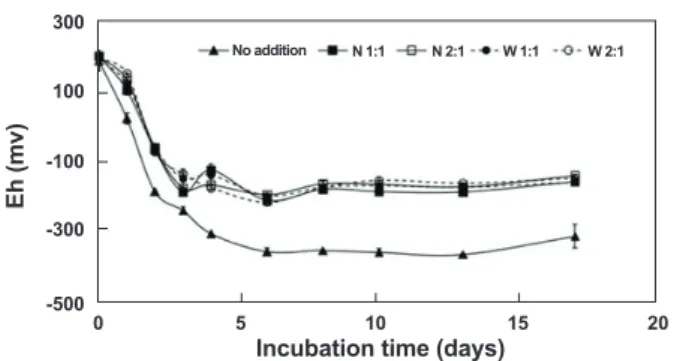
. 산화환원전위가 양의 값을 보이는 수계 등의 산화적 환경에서는 주로 호기성 분해가 진행되며, 음의 값을 보이는 연안퇴적층 등의 환원적 환경에서는 휘발성지방 산, 황화수소 및 메탄 등이 생성되는 혐기성 분해가 진행된다.Fig. 2
는 배양기간 산화환원전위의 변동을 보여주며, 황토첨가 실험계는 배양 3일까지 감소하여
17
일까지는-128~-191
의 범위를 유지하였으며, 미첨가 실험계는 배양6
일까지 감소하 여-317~-369
의 범위였다. 배양 시작일에는 황토첨가 실험계 와 미첨가 실험계간에 차이가 없었으나, 배양1
일부터 황토 미 첨가 실험계의 산화환원전위가 낮아져 배양4
일 경과 후부터 황토 미첨가 실험계의 산화환원전위가 첨가 실험계에 비하여-140~-206 mV
정도 낮았다. 또한, 황토첨가 실험계간에는 황 토 채취산지와 황토 첨가비율에 따른 산화환원전위의 변동차 이가 없었다. 유기물 분해과정 중 산화환원전위의 변동으로 황 토첨가는 환원적 환경조성을 억제하는 효과가 있음을 알 수 있었으며, 이는 유기물 분해에 관여하는 황토입자에 서식하는 미 생물군집 등에 의한 산소소모 및 황토 자체가 가지고 있는 화학 적 구성성분 등의 영향으로 사료되며,
Nita et al. (1985)
는 해 양 퇴적층에 점토광물 살포시 산소소비속도를 천천히 하는 효 과가 있다고 보고하였다.산휘발성 황화물의
(AVS: acid volatile sulfide)
변동을Fig. 3
에 나타내었으며, 산휘발성 황화물은 연안 퇴적층 유기물의 혐 기성 분해과정에서 생성되는 황화수소 등의 황산화물의 총칭Fig. 1. Fluctuations of pH during anaerobic decomposition of
Cochlodinium polykrikoides in the microcosm system.
▲: No addition of Yellow clay, ■:Yellow clay collected Namhae area added to the sediment in the dry/wet weight ratio of 1:1, □:
Yellow clay collected Namhae area added to the sediment in the dry/wet weight ratio of 2:1, ●: Yellow clay collected Wando area added to the sediment in the dry/wet weight ratio of 1:1, ○: Yel- low clay collected Wando area added to the sediment in the dry/
wet weight ratio of 2:1. All data are expressed as the mean±SE (n=3)
Fig. 2. Fluctuations of Eh during anaerobic decomposition of Cochlodinium polykrikoides in the microcosm system.
▲: No addition of Yellow clay, ■:Yellow clay collected Namhae area added to the sediment in the dry/wet weight ratio of 1:1, □:
Yellow clay collected Namhae area added to the sediment in the dry/wet weight ratio of 2:1, ●: Yellow clay collected Wando area added to the sediment in the dry/wet weight ratio of 1:1, ○: Yel- low clay collected Wando area added to the sediment in the dry/
wet weight ratio of 2:1. All data are expressed as the mean±SE (n=3)
Fig. 3. Fluctuations of acid volatile sulfide (AVS) during anaero- bic decomposition of Cochlodinium polykrikoides in the micro- cosm system.
▲: No addition of Yellow clay, ■:Yellow clay collected Namhae area added to the sediment in the dry/wet weight ratio of 1:1, □:
Yellow clay collected Namhae area added to the sediment in the dry/wet weight ratio of 2:1, ●: Yellow clay collected Wando area added to the sediment in the dry/wet weight ratio of 1:1, ○: Yel- low clay collected Wando area added to the sediment in the dry/
wet weight ratio of 2:1. All data are expressed as the mean±SE (n=3)
pH Eh (mv)
Incubation time (days) Incubation time (days)
Incubation time (days)
9.0 300
100 -100 -300
-5000 5 10 15 20
8.0
7.0
6.00 5 10 15 20
No addition N 1:1 N 2:1 W 1:1 W 2:1 No addition N 1:1 N 2:1 W 1:1 W 2:1
AVS (mg/L)
150 100 50
00 5 10 15 20
No addition N 1:1
W 2:1 N 2:1
W 1:1
으로 양식장 및 연안역 퇴적층의 유기물 오염지표로 이용된다.
산화환원전위가
-100 mV
이하로 환원된 연안 퇴적층에서 혐 기성세균인 황산염환원세균(Postgate, 1984)
이 에너지원으로acetate
및propionate
등 휘발성 지방산을 이용하여 최종산물 로 황화수소를 생산한다(Winfrey and Ward, 1983).
발생된 황 화수소는 주변에 서식하는 해양생물에 직접적인 영향을 미치 거나 해양생물에 유해한 환경을 조성한다. 산휘발성 황화물은 모든 실험계에서 배양3
일 경과 후 생성되기 시작하여6
일까지 급속히 증가하여 그 이후로는 일정한 농도였다.6
일 이후 산휘 발성 황화물 농도는 황토첨가 실험계가40.2-69.3 mg/L
의 범 위로 황토미첨가 실험계83.2-93.8 mg/L
에 비하여 약1.2-2.2
배 낮게 생성되었다. 분해과정 중 황토첨가 실험계의 산휘발 성 황화물이 미첨가 실험계에 비하여 낮게 생산되었으며, 이는Park and Lee (1998)
의 실내실험계를 통하여C. polykrikoi- des
분해과정 중 해양세균 군집변동 조사에서 황토첨가 실험계 가 미첨가 실험계에 비하여 산휘발성 황화물의 생성농도가 낮 았다는 결과와 일치하며,Choi et al. (1998)
이 진주만의 퇴적 층에 황토를 살포 후 산휘발성 황화물을 조사한 결과 황토를 살 포한 지역에서 산휘발성 황화물의 농도가 낮았다는 조사결과 와 일치한다. 또한, 황토 첨가비율이 높은 실험계에서 산휘발 성 황화물이 낮게 생성되었으며, 이는 황토첨가가 유기물 분해 과정에서 환원적 환경으로의 진행을 억제하여 황산염환원세균 의 활성저해와 황화수소 발생을 억제한 것으로 사료된다. 또 한, 완도 황토를 첨가한 실험계에서 산휘발성 황화물의 생성농 도가 남해 황토첨가에 비하여 높았으며, 이는 황토 채취지역에 따라 황토의 구성광물 비율 및 화학적 조성의 차이(Hwang et al., 2000)
가 있으며 이와 관련이 있는 것으로 사료된다.분해과정 중 휘발성 지방산의 변동
휘발성 지방산은 유기물의 혐기성 분해과정에서 생성되는 중 간 대사산물로서 수계퇴적물, 폐수처리 반응조 및 동물의 장 내 등 다양한 혐기성 환경에서 검출되며
(Balba and Nedwell,
1982)
, 유기물의 혐기성 분해과정의 최종단계인 메탄생성과황산염환원에 중요한 에너지원이 된다. 그러나 해수역에서는 전자수용체인 황산염이 풍부하여 해양환경에 악 영향을 미치 는 황화수소가 생성되는 황산염환원에 우선적으로 이용된다.
따라서 오래 전부터 연안 퇴적층에서의 분포
(Parkes and Tay- lor, 1983; Molongoski and Taylor, 1985; Mueller-Harvey and Parkes, 1987; Kondo et al., 1990a)
, 해양 퇴적층의 유기 물 분해과정에서 거동(Balba and Nedwell, 1982; Boshker et al., 2001; Finke et al., 2007; Kondo et al., 1990b, 1993a; Tor et al., 2003; Valdemarsen and Kristensen, 2010)
및 생성에 미치는 환경요인(Kondo et al., 1993b; park et al., 1996)
등 다 양한 연구가 되어졌다.해양 퇴적층에서의 유기물의 혐기성 분해과정 중 많이 생성 되는 휘발성 지방산
acetate, butyrate, formate, lactate, propi-
onate
및succinate
를 분석하였으나, 본 연구의C. polykrikoi- des
분해과정에서는acetate, formate, propionate
로3
종의 휘 발성 지방산만 생성되었다. 이 중acetate
는 해양퇴적층에서 생성되는 주요한 발효산물(Mueller-Harvey and Parks, 1987;
Sansone and Martens, 1982)
로서, 유기물의 혐기성 분해과 정에서 가장 많이 생성되는 휘발성지방산으로 보고(Zeikus, 1983)
되었으며, 황산염환원세균이 이용하는 기질 중 약60
% 를 차지하는 주요 에너지원으로 알려져 있다(Banat et al., 1981; Winfrey and Ward, 1983; Skyring, 1988; Kondo et al., 1990b).
Fig. 4
는 배양동안acetate
의 변동을 나타낸 것으로 황산염환 원세균의 억제제인sodium molybdate
미첨가 실험계에서 배 양1
일부터4
일까지 모든 구간에서 생성되기 시작하였으며 황 토첨가 실험계가 미첨가 실험계에 비하여 약1.2-1.8
배 정도 적 게 생성되었지만, 첨가된 황토의 산지와 첨가비율에 따라서는acetate
의 생성 정도의 차이를 보이지 않았다. 또한,sodium molybdate
미첨가 실험계에서 배양4
일째부터6
일까지 급격히 감소하였으나, 첨가한 실험계에서acetate
의 농도는4
일 이후Fig. 4. Fluctuations of acetate during anaerobic decomposition of Cochlodinium polykrikoides in the microcosm system without sodium molybdate (A) and with sodium molybdate (B).
▲: No addition of Yellow clay, ■:Yellow clay collected Namhae area added to the sediment in the dry/wet weight ratio of 1:1, □:
Yellow clay collected Namhae area added to the sediment in the dry/wet weight ratio of 2:1, ●: Yellow clay collected Wando area added to the sediment in the dry/wet weight ratio of 1:1, ○: Yel- low clay collected Wando area added to the sediment in the dry/
wet weight ratio of 2:1. All data are expressed as the mean±SE (n=3)
Incubation time (days)
Incubation time (days)
Acetate (μM)
(A)
(B)
Acetate (μM)
400
400 200
200 0
00 2 4 6 8 10 12
2 4 6 8 10 12
No addition
No addition
N 2:1 N 1:1
W 1:1 W 2:1
N 2:1 W 2:1
N 1:1 W 1:1
Fig. 6. Fluctuations of Propionate during anaerobic decomposi- tion of Cochlodinium polykrikoides in the microcosm system without sodium molybdate (A) and with sodium molybdate (B).
▲: No addition of Yellow clay, ■:Yellow clay collected Namhae area added to the sediment in the dry/wet weight ratio of 1:1, □:
Yellow clay collected Namhae area added to the sediment in the dry/wet weight ratio of 2:1, ●: Yellow clay collected Wando area added to the sediment in the dry/wet weight ratio of 1:1, ○: Yel- low clay collected Wando area added to the sediment in the dry/
wet weight ratio of 2:1. All data are expressed as the mean±SE (n=3)
Propionate (μM)
No addition N 1:1
W 1:1 N 2:1
W 2:1 100
80 60 40 20
00 2 4 6 8 10
Propionate (μM)
0 2 4 6 8 10
No addition N 1:1
W 1:1 N 2:1
W 2:1 100
(B)
80 60 40 20 0
(A)
Incubation time (days) Incubation time (days)
로 약간 증가하거나 변동이 없어, 분해과정 중 생성된acetate
는 황산염환원세균에 모두 이용되어졌음을 일수 있었다. 이는
Kondo et al. (1993a)
및Laanbroek and Pfenning (1981)
의 실내실험계를 통한 해양 퇴적층에서 생성된acetate
는 황산염 환원세균에 의하여 대부분 소모되어졌다는 결과와 일치하였 다. 따라서 해양 퇴적층의C. polykrikoides
혐기성 분해과정 에서 황토 첨가는 황화수소를 생성하는 황산염환원세균의 주 요 에너지원인acetate
의 생성을 억제함을 간접적으로 알 수 있었다.Fig. 5
는formate
의 변동을 보여주는 것으로acetate
와 유사 한 농도로 생성되었다.Kondo et al. (1990b, 1993a)
에 의한 유기물 분해실험에서는 본 연구결과와 같이acetate
와 유사한 농도로 생산되었으나,Laanbroek and Pfenning(1981), Tay- lor and Parkes(1984)
,Park et al. (1997)
에 의한 해양 퇴적물 의 유기물 분해실험에서는 생성되지 않아, 퇴적물에 따라 유기 물 분해기작에 차이가 있음을 알 수 있었다.C. polykrikoides
혐기성 분해에서formate
는 황토첨가 및 미첨가 모든 실험계에 서acetate
에 비하여 빠른 속도로 생성되어 배양1
일째에 최고농도에 도달하였다. 황토첨가 실험계에서
formate
가 미첨가 실험계에 비하여1.3-2.8
배 낮은 농도로 생성되었으며, 황토첨 가 실험계 중 남해와 완도황토 모두 첨가 비율이 높은 실험구가 낮은 실험구에 비하여 낮은 농도로 생산되어 유기물의 혐기적 분해과정에서 황토구성 성분이formate
생산을 억제 함을 알 수 있었다. 또한,sodium molybdate
를 첨가한 실험계에서는 배양1
일 이후로 일정 한 농도를 유지하였으나,sodium mo-
lybdate
를 첨가하지 않은 실험계 중 황토미첨가 실험계에서는급격히 소모되어 배양
2
일째에는 검출되지 않았으나 황토첨가 실험계는 미첨가 실험계에 비하여 서서히 소모되어 배양3
일째 에 거의 소모되었다. 따라서 해양 퇴적층에서C. polykrikoides
혐기성 분해과정에서 황토첨가가 미첨가에 비하여formate
의 생성농도가 작았고 황산염환원세균에 의하여 천천히 이용됨 을 알 수 있었다.Propionate
도 해양 퇴적층 및 간석지 등의 혐기적 환경에서 분포하는 중요한 휘발성 지방산으로 보고(Banat et al., 1981)
되었으며, 대부분의 해양 퇴적층의 유기물 분해과정에서 생산 되는 것으로 보고(Kondo et al., 1993a; Laanbroek and Pfen-
Fig. 5. Fluctuations of formate during anaerobic decomposition of Cochlodinium polykrikoides in the microcosm system without sodium molybdate (A) and with sodium molybdate (B).
▲: No addition of Yellow clay, ■:Yellow clay collected Namhae area added to the sediment in the dry/wet weight ratio of 1:1, □:
Yellow clay collected Namhae area added to the sediment in the dry/wet weight ratio of 2:1, ●: Yellow clay collected Wando area added to the sediment in the dry/wet weight ratio of 1:1, ○: Yel- low clay collected Wando area added to the sediment in the dry/
wet weight ratio of 2:1. All data are expressed as the mean±SE (n=3)
Formate (μM) Formate (μM)
600
600 400
400 200
200 0
0 0
0
2
2
4
4
6
6
(A)
No addition N 1:1W 1:1 N 2:1
W 2:1
Incubation time (days)
Incubation time (days)
No addition N 1:1
W 1:1 N 2:1
W 2:1
(B)
ning, 1981; Taylor and Parkes, 1984; Park et al., 1997)
되었 다. 본 연구에서propionate
도acetate
와formate
와 마찬가지 로 배양 후 황토첨가 및 미첨가 모든 실험계에서 생성되기 시 작하여2
일째 농도가 가장 높았으며 황토첨가 실험계가 미첨 가 실험계에 비하여1.5-1.8
배 정도 낮은 농도로 생성되었으나,
formate
와는 다르게 황토 첨가농도에 따라서 뚜렷한 차이를 보이지 않았다
(Fig. 6).
또한, 배양2
일 이후로sodium
molybdate
를 첨가한 황토첨가 및 미첨가 실험계에서 조금씩증가하는 경향을 보여주었으나, 첨가하지 않은 모든 실험계에 서 배양 4일째까지 급격히 소모되어 황산염환원세균에 의하여 이용됨을 알 수 있었다. 따라서
propionate
도 해양 퇴적층에서C. polykrikoides
혐기적 분해과정에서 황토첨가가 미첨가에 비하여propionate
의 생산량이 낮아 황산염환원세균에 의하여 이용되는 양이 적음을 알 수 있었다.Table 1
에C. polykrikoides
혐기성 분해 중 황산염환원세균 에 이용된3
종의 휘발성 지방산의 농도를 나타내었다. 생성된 모든 휘발성 지방산이 황토첨가 실험계가 미첨가 실험계에 비 하여 황산염환원세균에 의하여 이용되는 양이 적었으며, 이용 된 양은acetate
와formate
가 유사하였고propionate
가 약1/4
정도의 수준이었다. 또한 이용된 순서는 산휘발성 황화물이 생 성되기 시작하는 배양
2
일째부터propionate
가 가장 먼저, 다 음으로formate, acetate
의 순이었다.적조방제로 살포한 황토가 해양퇴적층에서
C. polykrikoides
의 혐기성 분해에 미치는 영향을microcosm
실험계를 이용하 여 간접적으로 알아본 결과를 요약하면,C. polykrikoides
의 분해과정에서 황토첨가는 환원적 환경을 나타내는 음의 산환 환원전위로 되는 것을 억제하며, 황산염환원세균의 에너지원 으로 이용되는 휘발성 지방산인acetate, formate
및propio- nate
의 생성을 낮게 하고, 최종적으로 해양생물에 직접적인 악 영향을 주거나 해양환경을 악화시키는 황화수소 생성을 제어 함을 간접적으로 알 수 있었다. 그러나 본 실험에서는 황토 첨 가가 혐기적 환경조성과 휘발성 지방산의 생성억제와 이에 따 른 황화수소생성을 억제함을 알 수 있었으나 황토의 어떤 기작 에 의한 것인지는 차후 연구되어야 할 것으로 사료된다.사 사
본 연구는 국립수산과학원 적조모니터링 및 대응연구(
RP-
2012-OE-15)
의 연구지원에 의하여 수행되었습니다.Table1. Utilized volatile fatty acids(VFAs) by sulfide-reducing bacteria during anaerobic decomposition of Cochlodinium polykrikoides in the microcosm system
Periodes (days)
Total utilized VFAs
1-2 2-3 3-4 4-6 6-8 8-10 10-23
Acetate
No addition1 0.0 0.0 357.0 10.7 7.5 0.0 375.2
N 1:12 0.0 0.0 117.6 18.1 86.5 0.0 222.2
N 2:13 0.0 0.0 158.6 30.9 10.8 0.0 200.3
W 1:14 0.0 0.0 167.4 77.6 10.3 0.0 255.3
W 2:15 0.0 0.0 203.2 0.0 0.0 0.0 203.2
Formate
No addition 0.0 4.1 0.0 0.0 0.0 0.0 432.0
N 1:1 0.0 209.4 0.0 0.0 0.0 0.0 266.3
N 2:1 0.0 51.9 0.0 0.0 0.0 0.0 185.2
W 1:1 0.0 178.2 0.0 0.0 0.0 0.0 210.3
W 2:1 0.0 108.6 0.0 0.0 0.0 0.0 175.5
Propionate
No addition 0.0 10.4 20.8 5.8 0.0 0.0 82.3
N 1:1 0.0 30.0 2.5 5.9 0.0 0.0 53.4
N 2:1 0.0 26.1 6.4 3.4 0.0 0.0 45.7
W 1:1 0.0 16.8 9.6 2.9 0.0 0.0 50.2
W 2:1 0.0 26.9 2.2 1.3 0.0 0.0 46.5
1
No addition of Yellow clays
2
Addition of Yellow clay collected Namhae area to the sediment in the dry/wet ratio of 1:1.
3
Addition of Yellow clay collected Namhae area to the sediment in the dry/wet ratio of 2:1.
4
Addition of Yellow clay collected Wando area to the sediment in the dry/wet ratio of 1:1.
5
Addition of Yellow clay collected Wando area to the sediment in the dry/wet ratio of 2:1.
참고문헌
Bae HM, Choi HG, Lee WC and Yoon SJ. 1998. Control of the red tide by yellow clay dispersion. In: Proceedings of Korea-China Joint Symposium on Harmful Algal Blooms.
Kim HG, Lee SG and Lee CK eds. Gooduck Press, Busan, Korea. 53-60.
Balba MT and Nedwell DB. 1982. Microbial metabolism of acetate, propionate and butyrate in anoxic sediment from the Colne Point Saltmarsh. Essex UK J of Gener Microbiol 128, 1414-1422.
Banat IM, LindströM EB, Nedwell DB and balba MT. 1981.
Evidence for the existence of two distinct functional groups of sulfate-reducing bacteria in salt marsh desiment. Appl and Environ Microbiol 42, 985-992.
Bisutii I, Hilke I and Raessler M. 2004, Determination of to- tal organic carbon an overview of current method, Tends in Analy Chem 23, 716-72.
Boschker HTS, Graaf W, Koster M, Meyer-Reil LA and Cap- penberg TE. 2001. Bacterial populations and processes involved in acetate and propionate consumption in anoxic brackish sediment. FEMS Microbiol Ecol 35, 97-103.
Canfield DE, Thamdrup B and Kristensen E. 2005. Aquatic Geomicrobiology. Elsevier Academic Press Sandiego, 656.
Choi HG, Kim PJ, Lee WC, Yun SJ, Kim HG and Lee HJ. 1998.
Removal efficiency of Cochlodinium polykrikoides by yel- low loess. Kor J Fish Soc 31, 109-113.
Finke N, Vandieken V and Jørgensen BB. 2007. Acetate, lac- tate, propionate and isobutylate as electron donors for iron and sulfate reduction in Artic marine sediments Svalbarde.
FEMS Microbiol Ecol 59, 10-22.
Guillard RRI and Ryther JH. 1962. Studies of marine plankton- ic diatoms 1.Cyclotella nana HUSTEDT and Detonula con-
fervacea (CLEVE) GRAN, Can J. Microbiol 8, 229-239.
Hwang JY. 1997. Characteristics and application of Macbansuk and Hwangto. In : Proceedings of the 10 years anniversity symposium. The Miner Soc Korea, 89-98.
Hwang JY, Jang MI, Kim JS, Cho WM, Ahn BS and Kang SW.
2000. Mineralogy and chemical composition of the residual soils (Hwangto) from South Korea. J Miner Soc Korea 13, 147-163.
Jørgensen BB. 1982. Mineralization of organic matter in the sea bed-the role of sulphate reductions. Nature 296, 643-645.
Jørgensen BB. 1983. Processes at the sediments-water interface.
In : The major biogeochemical cycles and their interactions.
Bolin B and Cooks RB, eds. SCOPE Wiley, 477-515.
Jørgensen BB, Bang M and Blackburn TH. 1990. Anaerobic mineralization in marine sediments from the Baltic Sea- North Sea transition. Mar Eco Prog Ser 59, 39-54.
Kim KH, Son SK, Son JW and Ju SJ. 2006. Methodological comparison of the quantification of total carbon and organic carbon in marine sediment. Mar Environ Eng 9, 235-242.
Kim SJ. 2000. Removal of red tide organisms: 2. Flocculation
of red tide organisms by using loess. J Kor Fish Soc 33, 455-462.
Kim PJ, Heo S and Yun SJ. 2002. Adsorption and removal mechanism of dissolved inorganic nutrients in seawater by Yellow loess. J Kor Fish Soc 35, 146-154.
Kondo R, Kitada H, Kawai A and Hata Y. 1990a. Low molecu- lar fatty acids in the marine sediment of eutrophic coastal regions. Nippon Suisan Gakkaishi 56, 519-523.
Kondo R, Nishijima T and Hata Y. 1990b. Acetate-oxidizing sulfate reduction in anoxic marine sediment. Bull Jap Soc Microb Ecol 5, 29-35.
Kondo R, Nishijima T and Hata Y. 1993a. Mineralization pro- cess of glucose and low molecular fatty acid production in an anoxic marine sediment slurry. Nippon Suisan Gakkaishi 59, 105-109.
Kondo R, Nishijima T and Hata Y. 1993b. Effect of temperature on the production of low molecular fatty acids within an anoxic marine sediment slurry. Nippon Suisan Gakkaishi 59, 1189-1194.
Laanbroek HJ and Pfenning N. 1981. Oxidation of short chain fatty acids by sulfate reducing bacteria in freshwater and in marine sediments. Arch Microbiol 128, 330-335.
Molongoski JJ and taylor CD. 1985. High-performance liquid chromatography of short- and long- chain fatty acids from seawater. Applied and Environ Microbiol 50, 1112-1114.
Mueller-Harvey I and Parkes RJ. 1987. Measurement of vol- atile fatty acids in pore water from marine sediments by HPLC, Est Coast Shelf Sci 25, 567-579.
Na GH, Choi WJ and Chun YY. 1996. A study on red tide con- trol with clay suspension. J Aquaculture 9, 239-245.
Nita T, Ito S, Inoue T and Iwazaki K. 1985. Guideline for the remediation of the bottom sediment environment. Japan Fisheries Resource Conservation Association, 110.
Oremland RS and Taylor BF. 1978. Sulfate reduction and meth- anogensis in marine sediments. Geochim Cosmochim Acta 42, 209-214.
Park CH and Lee BH. 2006. Effects of loess application in coastal benthic ecosystem, J Environ Sci 15, 1035-1043.
Park YT, Nishimura M and Ohwada K. 1996. Effects of chemi- cal composition and temperature for the production of vola- tile fatty acids during anaerobic decomposition process of marine sinking particles. J Korean Fish Soc 29,888-892.
Park YT, Nishimura M and Ohwada K. 1997. Study of marine sulfate-reducing bacterial population using fluorescent in
situ hybridization method during decomposition processes
of detrial material and polypepton in microcosm. Fish Sci 63, 99-104.Park YT and Lee WJ. 1998. Changes of bacterial population during the decomposition process of red tide dinoflagellate,
Cochlodinium polykrikoides in the marine sediment addi-
tion of yellow loess. J Korean Fish Sci 31, 920-926.Parkes RJ and Taylor J. 1983. Analysis of volatile fatty acids by ion-exclusion chromatography, with special reference to
marine pore water. Marine Biology 77, 113-118.
Postgate JR. 1984. The sulfate reducing bacteria, 2nd ed, Can- bridge University Press Cambridge 208.
Sansone FJ and Martens CS. 1982. Volatile fatty acids cycling in organic rich marine sediments. Geochim Cosmochim Acta 46, 1575-1589.
Seo KS, Lee CK, Park YT and Lee Y. 2008. Effect of yellow clay on respiration and phytoplankton uptake of bivalves.
Fish Sci 74, 120-127.
Shirota A. 1976. Meeting on control of red tide with clay. Sei- kai Reg Fish Res Lab 24, 2-5.
Shirota A. 1989. Red tide problem and countermeasures (2). Int J Aquat Fish Technol 1, 195-223.
Shiozawa T, Kawana K, Hosika A, Tanimoto T and Takimura O. 1979. The Characteristics of sediment in the Seto Inland Sea. Rep Gov Ind Res Inst Chugoku 4, 1-24.
Skyring GW. 1988. Acetate as the main energy substrate for the sulfate-reducing bacteria in Lake Eliza(South Australia) hypersaline sediments. FEMS Microbiol Ecol 53, 87-94.
Sørensen FJ, Christensen D and Jørgensen BB, 1981. Volatile fatty acids and hydrogen as substrates for sulfate reducing bacteria in anaerobic marine sediment. Applied and Envi- ron. Microbiol 42, 5-11.
Taylor J and Parkes J. 1984. Identifying different populations of
sulfate-reducing bacteria within marine sediment systems, using fatty acid biomarkers. J Gen Microbiol 131, 631-642.
Tor JM, Amend JP and Lovley DR. 2003. Metabolism of organ- ic compounds in anaerobic, hydrothermal sulfate reducing marine sediments. Environ Microbiol 5, 583-591.
Valdemarsen T and Kristensen E. 2010. Degradation of organic monomers and short-chain fatty acids in sandy marine sedi- ment by fermentation and sulfate reduction. Geochim Cos- mochim Acta 74, 1593-1605.
Winfrey MR and Ward DM. 1983. Substrates for sulfate reduc- ing and methane production in intertidal sediments. Applied and Environ Microbiol 45, 193-199.
Yu Z, Sengco MR and Anderson DM. 2004. Flocculation and re- moval of the brown tide organism, Aureococcus anophagef-
ferens (Chrysophyceae),using clays. J Applied Phycol 16, 101-
Zeikus JG. 1983. Metabolic commucation between biodegrada-110.tive populations in nature. In : Microbes in their natural en- vironments. Slater JH, Whittenbury R and Wimpenny JWJ, eds. Cambridge Uni Press Cambridge, 423-462.
Shaw DJ, Alperin MJ, Reeburgh WS and Mc-Intosh DJ. 1984.
Biogeochemistry of acetate in anoxic sediment of Skan Bay, Alaska. Geochim Cosmochim Acta 48, 1819-1825.