인삼 체세포 배 발아를 위한 GA 3 의 최적 조건
김영창*·박홍우*·김옥태†·방경환·현동윤·차선우·김동휘 충북 음성군 농촌진흥청, 국립원예특작과학원, 인삼특작부
The Effects of Optimal Germination of Somatic Embryos Induced from Mature Cotyledon Explants of Panax ginseng C. A. Meyer by Gibberellic Acid
Young Chang Kim
*, Hong Woo Park
*, Ok Tae Kim
†, Kyong Hwan Bang, Dong Yun Hyun, Seon Woo Cha, and Dong Hwi Kim
Department of Herbal Crop Research, National Institute of Horticultural and Herbal Science, Rural Development Administration, Eumseong 369-873, Korea.
ABSTRACT : Somatic embryos on growth regulator-free medium can be produced directly from cotyledon explants of
Panax ginseng C. A. Meyer. When the embryo developmental stage was torpedo and cotyledon, the germination rate of embryos was quite high on MS medium supplemented with gibberellic acid (GA
3). However, the percentage of plantlet for- mation at the cotyledon stage was higher than that at the torpedo stage. This result demonstrates that the embryo at the cot- yledon stage was the most appropriate for increasing germination by GA
3. Embryos cultured on medium including four levels of GA
3concentrations (3, 5, 10, or 20 ㎎ / ℓ ) showed all quite high germination rates (87-91%). When the well-devel- oped embryos were continuously cultured on media including high concentrations of GA
3from 10 to 20 ㎎ / ℓ , the percent- age of formation of normal plantlets was lower than that seen under low concentrations from 3 to 5 ㎎ / ℓ . This treatment of high concentrations resulted in shoots with abnormal shape. The optimal GA
3treatment provides a basis for the efficient method obtaining healthy plantlets derived from ginseng somatic embryos.
Key Words : Direct Somatic Embryogenesis, Panax ginseng , Gibberellic acid, Germination
INTRODUCTION
Panax ginseng C. A. Meyer, commonly known as Korean or Asian ginseng, is a perennial herb native to Korea, China, and Southeast Asia, and has been utilized for thousands of years as an herbal remedy in East Asia. Its roots are highly prized for its purported medicinal properties (Proctor and Bailey, 1987).
Ginseng has a long production cycle, as ginseng seeds are generally produced after a 3-year period of cultivation (Zhou and Brown, 2006) and must be stratified for an additional 3 months prior to germination. Problems in the long reproductive and production cycles of ginseng breeding are generally associated with interspecific cross-breeding. The inflorescence is an umbel, and the plant is a perennial
which required approximately 4-6 growing years before the yield could be assessed. The breeding objectives of ginseng might include reduced seed set time, increased yield, higher quality roots, and disease resistance (Proctor and Bailey, 1987). Breeding work with perennial crops, such as ginseng, is rather time-consuming for farmers, as the long period makes it too late for the seeds to propagate, and thus ginseng is only propagated by the form of seeds. It was taken a time over 40 years to develop eight varieties of ginseng in Korea. However, it is generally presumed that ginseng should be farm-cultivated, due to its late propagation rate. Thus, it is necessary to develop a tissue culture technique that circumvents the long-term problems of conventional breeding by using millions of in vitro cultured plants, and by repeating the experiment all year round in
†
Corresponding author: (Phone) +82-43-871-5587 (E-mail) [email protected] Received 2009 June 8 / Revised 2009 July 21 / Accepted 2009 July 29
*Kim YC and Park HW contributed equally to this work.
established using zygotic embryo explants (Choi and Soh, 1996; Choi and Soh, 1997; Choi et al ., 1998a; Choi et al ., 1998b; Kim et al ., 2006).
These protocols entail the production of somatic embryos, their germination, and the acclimation of regenerated plantlets into the soil. When the generated somatic embryos were germinated with gibberellic acid (GA
3), they promoted embryo germination and stem elongation (Sun and Gubler, 2004), however, negative effects, including the abnormal elongation of embryos, can be spontaneously generated, despite the fact that a number of direct somatic embryos capable of becoming normal plants were derived from the explants. Finally, the acclimation rate was very low when regenerated plantlets transplant the soil. Although GA
3has been previously utilized to germinate somatic embryos derived from zygotic embryo explants, there has been no report concerning the optimal embryo developmental stage and its concentration in tissue cultures of Korean ginseng using cotyledon explants.
In the present report, we describe the somatic embryos derived from mature zygotic embryos of Panax ginseng . In addition, the effect of GA
3on the developmental embryonic stage and the optimal GA
3concentration were assessed in an effort to achieve more efficient plant regeneration.
MATERIALS AND METHODS
1. Plant Material and culture condition
Stratified P. ginseng seeds were acquired from the Depart- ment of Herbal Crop research of the RDA (Korea). All media were adjusted to a pH 5.8 using 1N HCl or 1N NaOH prior to the addition of agar, and were subsequently autoclaved for 15 min at 121 ℃ . The culture room was maintained at 23 ± 2 ℃ and all cultures were conducted under a 16-h photoperiod with irradiance (24 µ㏖ /s/ ㎡ from white-fluorescent tubes) provided by cool fluorescent tubes.
embryos were generated as previously described by Kim et al. (2006).
3. Germination of somatic embryos
For germination, somatic embryos at four different developmental stages (globular, heart, torpedo, and cotyledon) were cultured on germination medium (30 ㎖ of MS medium supplemented with 5 ㎎ / ℓ GA
3, 3% sucrose, 0.8%
agar in 98 × 15 ㎜ Petri dishes). These embryos from the globular to cotyledonary stage were isolated carefully from explants under a microscope (ICS-305B, Alphasystec). Fifteen embryos at each particular stage were inoculated on germination medium. After 5 weeks of cultivation, the number of germinated embryos produced per petri dish was recorded. Five of the germinated somatic embryos were transferred onto free-GA
31/2 MS medium supplemented with 3% sucrose and 0.35% gelrite in a bottle (7 × 12 ㎝ ).
A normal plant is defined as a shoot exhibiting good shape and a root without a callus. After 5 weeks of culture, the percentages of normal plant formation were recorded. These regenerated plantlets were subcultured on the same medium every 4 weeks until they were 7 ㎝ tall.
In order to assess the optimal GA
3concentration, the
well-developed somatic embryos of a cotyledonary stage
between one and two months of cultivation were transferred
onto the germination medium. The effect of GA
3concen-
tration on germination was assessed by placing the mature
somatic embryos on MS basal medium containing GA
30,
3, 5, 10, or 20 ㎎ / ℓ . Fifteen well-developed somatic embryos
per petri dish were inoculated. The germination rate and the
percentage of normal plantlet formation were recorded as
described by the above method. In all experiments, there
were three replicates per treatment; the experiment was
repeated three times. Statistical calculations were conducted
via analysis of variance (ANOVA), and the results were
assessed via Duncan’s multiple range test (DMRT).
4. Scanning Electron Microscope
The samples were fixed in 1% glutaraldehyde (phosphate buffer pH6.8) for 4 h, then dehydrated with ethyl alcohol.
The specimens were mounted on aluminum stubs, sputter- coated with gold in a Polaron SC 7610 sputter coater (FISION Instruments), and viewed in a LEO Stereoscan 440 SEM (ZEISS) at an accelerating voltage of 15 ㎸ . The images were recorded on TIF graphic files.
RESULTS AND DISCUSSION
Direct somatic embryos were obtained from the lateral edges and adaxial surfaces of the zygotic embryo explants on MS basal medium without plant growth regulators, as was previously described by Kim et al. (2006) (Fig. 1a).
When the somatic embryos reached the cotyledonary stage, they stopped growing or grew very slowly. Although the somatic embryos were generated from the explants, they must be treated with GA
3to break their dormancy as zygotic embryos (Choi et al ., 1999). As is shown in Fig.
1b and 1c, shoot elongation was achieved, and the shoot spontaneously rooted after GA
3treatment. After 8 weeks of treatment with continuous subculture, the regenerated plantlets
that rose to a height of over 7 ㎝ could be transferred into the soil to increase survival against the ex vitro environment (Fig. 1d). The survival rate of healthy plantlets into the soil is relatively high (unpublished). Although the rate of somatic embryo production was relatively high, growth problems occurred during the plant regeneration procedure.
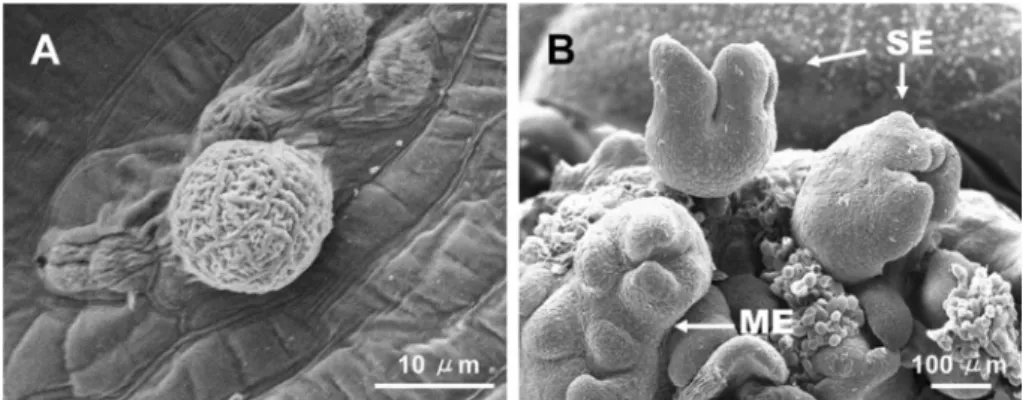
In order to obtain more normal plants via the regeneration procedure, a somatic embryo should be induced from a single cell, as is shown in Fig. 2a. The number of fused embryos (Fig. 2b) should also be diminished, as they also produced only multiple shoots, from which good quality of roots could not be acquired (Choi et al ., 1998b). Several studies have been conducted to obtain normal plantlets more efficiently. The plasmolysis and osmotic stress induced by high sucrose concentrations enhanced single embryo formation (Choi and Soh 1997; Choi et al ., 1999). Langhansová et al . (2004) previously reported that abscisic acid and polye- thylene glycol caused a slight improvement in the root meristem organization of ginseng somatic embryos.
After 2-5 weeks on growth regulator-free MS medium, four different developmental stages (globular, heart, torpedo and cotyledon) of somatic embryos were transferred onto MS basal medium containing GA
3, and were germinated
Fig. 1.
Direct somatic embryogenesis from mature zygotic embryos. (A) Direct somatic embryos from a zygotic embryo cultured on
growth regulator-free MS medium. (B) Embryo germination after 2 weeks by GA
3of 5
㎎/
ℓ. (C) Rooted plantlets from elongated
shoots. (D) Regenerated plantlets with a taproot before the acclimation.
over 3-5 weeks. According to the developmental stages of the embryos, different germination rates were observed (Table 1). After free-GA
3treatment, the embryos of the globular stage stopped growing as well as germinating.
When the embryo developmental stage was the heart, the level of germination and percentage of plantlet conversion was low; abnormal shoots were also observed at this stage after exposure to GA
3. When the embryo developmental stage was torpedo and cotyledon, the germination rate of embryos by GA
3was quite high. However, the percentage of formation into normal plantlets at the cotyledon stage was higher than that at the torpedo. Our results demonstrate that well-developed embryos at the cotyledon stage were the most suitable for increasing germination by GA
3.
In an effort to assess the optimal GA
3concentration, well-developed embryos were maintained for 5 weeks on media including different concentrations. No growth of somatic embryos cultured on the control medium without GA
3, was observed after 5 weeks of cultivation, whereas all
of the germination rates of embryos cultured on the medium including individual concentrations of GA
3were high (87-95%) (Table 2). Our results were consistent with those of Choi et al . (1999). Therefore, our results indicate that the germination effect of the different concentrations used did not differ significantly. When the well-developed embryos were continuously cultured on media with high concentrations of GA
3from 10 to 20 ㎎ / ℓ , the percentage of formation into normal plantlets was lower than that seen with low concentrations from 3 to 5 ㎎ / ℓ . Abnormally- shaped shoots grew at a high concentration. As is shown in Fig. 3, the surface of a shoot at a 5 ㎎ / ℓ concentration of GA
3did not differ from that of the wild-type, while after 4 weeks callus were formed in GA
3of 20 ㎎ / ℓ . These results showed that GA
3treatment at high concentrations negatively affects embryo germination and thus obtaining a good plantlet. In the case of American ginseng ( Panax quinquefolius L.), a relatively high concentration (10-20
Fig. 2.
Scanning electron microscope photographs of somatic embryos. (A) A single cell shown to globular stage of embryo. (B) Mature somatic embryos cultured on growth regulator-free MS medium after 5 weeks. SE-single embryo, ME-multiple embryo fused with other embryo.
Table 1.
Effect of GA
3and somatic embryo stage on germination and formation of normal plantlets.
Embryo stages Germination rate of embryos (%)
aFormation to normal plantlets in 5 weeks (%)
Globular 0c 0d
Heart 32.6±5.0b 12.8±2.0c
Torpedo 92.5±8.0a 67.4±7.0b
Cotyledon 95.5±10.1a 83.2±8.5a
a
Particular embryos on MS medium supplemented with 5
㎎/
ℓGA
3for 5 weeks.
Each value represents mean ± SE of 45 embryos in three repeated experiments.
Means followed by the same letters are not significantly different at P = 0.05 of DMRT
Table 2.
Influence of different concentration of GA
3on germination and formation of normal plantlets.
Concentration (
㎎/
ℓ)
of GA3 Germination rate
of embryos (%)
aFormation to normal plantlets in 5 weeks (%)
0 0b 0c
3 91.1±5.3a 85.6±3.8a
5 95.4±7.0a 87.2±7.9a
10 89.3±6.6a 61.2±4.2b
20 87.7±11.3a 55.7±5.1b
a
Mature somatic embryos on MS medium supplemented with GA
3for 5 weeks.
Each value represents mean ± SE of 45 embryos in three repeated experiments.
Means followed by the same letters are not significantly different at
P = 0.05 of DMRT
㎎ / ℓ ) of GA
3resulted in the optimal germination and production of normal plantlets (Zhou and Brown, 2006).
In conclusion, our results showed that the optimal concentration of GA
3in germination medium was 3-5 ㎎ / ℓ , and cotyledon was the most stage in ginseng somatic embryo. The majority of regenerated plants have thus far failed to survive over three years in the field. Thus, it is crucial to produce normal and healthy plantlets via the most efficient protocols with regard to acclimatization and regeneration on the soil. The results we observed with the optimal GA
3treatment provide the basis for a method obtaining efficiently healthy plantlets derived from ginseng somatic embryos, and it may contribute to the development of an efficient acclimatization system into soil.
LITERATURE CITED
Arya S, Arya ID and Eriksson T. (1993). Rapid multiplication of adventitious somatic embryos of
Panax ginseng. Plant Cell, Tissue and Organ Culture. 34:57-162.
Butenko RG, Brushwitzky IV and Slepyan LI. (1968) Organogenesis and somatic embryogenesis in the tissue culture of
Panax ginsengC. A. Meyer. Botanicheski Zhurnal. 7:906- Chang WC and Hsing YI. 913. (1980). In vitro flowering of embryoids derived from mature root callus of ginseng (
Panax ginseng). Nature. 284:341-342.
Choi KT, Kim MW and Ghin HS. (1981). Root and shoot formation in explant and callus derived from root and cotyledon of ginseng (
Panax ginsengC. A. Meyer). Korean Journal of Ginseng Science. 5:35-40.
Choi YE and Soh WY. (1996). The role of excision on somatic embryogenesis from mature ginseng zygotic embryos.
Phytomorphology. 46:151-159.
Choi YE and Soh WY. (1997). Enhanced somatic single embryo formation by plasmolyzing pretreatment from cultured ginseng
cotyledons. Plant Science. 130: 197-206.
Choi YE, Yang DC and Choi KT. (1998a). Induction of somatic embryos by macrosalt stress from mature zygotic embryos of
Panax ginseng
. Plant Cell, Tissue and Organ Culture. 52:177- Choi YE, Yang DC, Park JC, Soh WY and Choi KT. 181. (1998b).
Regenerative ability of somatic single and multlple embryos from cotyledon of Korean ginseng on hormone-free medium.
Plant Cell Reports. 17:544-551.
Choi YE, Yang DC, Yoon ES and Choi KT. (1999). High- efficiency plant production via direct somatic single embryogenesis from preplasmolysed cotyledons of
Panax ginsengand possible dormancy of somatic embryos. Plant Cell Reports. 18:493-499.
Kim OT, Kim TS, In DS, Bang KH, Kim YC, Choi YE, Cha SW and Seong NS. (2006). Optimization of direct somatic embryogenesis from mature zygotic embryos of
Panax ginsengC. A. Meyer. Journal of Plant Biology. 49:348-352.
Kishira H, Takada M and Shoyama Y. (1992). Micropropagation of
Panax ginsengC. A. Meyer by somatic embryos. Acta Horticulturae. 319:197-202.
Langhansová L, Konrádová H and Van k T. (2004).
Polyethylene glycol and abscisic acid improve maturation and regeneration of
Panax ginsengsomatic embryos. Plant Cell Reports. 22:725-730.
Murashige T and Skoog F. (1962). A revised medium for rapid growth and bioassays with tobacco tissue. Physiologia Plantarum. 15:473-497.
Proctor JTA and Bailey WG. (1987). Ginseng: Industry, Botany, and Culture. Horticultural Reviews. 9:187-236.
Slepyan LI, Brushwitzky IV and Burenko RB. (1967).
Panaxginseng
C. A. Meyer as an object for introduction into tissue culture. Problems Pharmacology. 21:198-203.
Sun TP and Gubler F. (2004). Molecular mechanism of gibberellin signaling in plants. Annual Review of Plant Biology.
55:197-223.
Zhou S and Brown DCW. (2006). High efficiency plant production of North American ginseng via somatic embryogenesis from cotyledon explants. Plant Cell Reports.
25:166-173.
io
e