강원도 삼척 지역에서 소나무 이차림의 대상 벌채에 따른 초기 식생 재생 반응
1a정세영2⋅조용찬3⋅변봉규4⋅김혜진5⋅배관호6⋅김현섭7⋅김준수8*
Initial responses of vegetation regeneration after strip clear cutting in secondary Korean red pine (Pinus densiflora) forest in Samcheok, Gangwon-do, South Korea1a
Se-Yeong Jeong2, Yong-Chan Cho3, Bong-Kyu Byun4, Hye-Jin Kim5, Kwan-Ho Bae6, Hyun-Seop Kim7, Jun-Soo Kim8*
요 약
대면적 개벌의 대안으로서, 대상 개벌은 생산림지의 생태 조건 보전과 임분 재생을 모두 고려하는 작업법이다.
그러나 우리나라에서는 대상 벌채 기법의 적용 및 효율성에 대한 연구를 찾아보기 어렵다. 본 연구는 강원도 삼척에서 2012년부터 2014년까지 소나무림 대상 벌채 (10m, 20m 및 40m) 처리 전・후의 하층식생 반응 및 소나무 재생과 임분 환경 변화와의 상관성을 방형구법을 적용하여 조사 및 분석하였다. 하층식생 피도 및 종다양성은 대상 벌채 넓이에 상관없이 교목 종 및 광선호 종의 확장에 의해 증가하는 경향이었고, 신갈나무, 맑은대쑥 및 산딸기 등의 벌채 후 잔존 교목 및 광선호 종들의 활발한 증가가 관찰되었다. 대상 벌채 구역의 소나무 정착은 수관층 환경 (광량 및 수관열림 도) 및 토양나출도와는 정의 상관, 그리고 하층식생 및 임상 목질잔해물의 피도와는 음의 상관을 나타내었다.
주요어: 대상 벌채, 임분 재생, 소나무, 식물 다양성, 하층식생
1 접수 2015년 7월 22일, 수정 (1차: 2015년 10월 21일), 게재확정 2015년 10월 22일 Received 22 July 2015; Revised (1st: 21 October 2015); Accepted 22 October 2015
2 경북대학교 대학원 생태환경시스템학과 Dept. of Ecology and Environmental System, Graduate School, Kyungpook National Univ., Sangju 37224, Korea
3 국립수목원 Korea National Aboretum, Pocheon 11186, Korea
4 한남대학교 생명시스템과학과 Dept. of Biological Science and Biotechnology, Hannam Univ., Daejeon 305-811, Korea 5 한국산림생태연구소 Korea Forest Ecology Instititute, Daegu 41478, Korea
6 경북대학교 생태환경시스템학부School of Ecology & Environmental System, Kyungpook National Univ., Sangju 37224, Korea 7 국립산림과학원 산림생산기술연구소 Forest Practice Research Center, Korea Forest Research Institute, Pocheon 11185, Korea 8 자연과숲연구소 Nature and Forest Research Institute, Daegu 41478, Korea
a 이 논문은 국립산림과학원 일반연구과제 “주요 경제수종의 천연갱신 기법 및 수확작업에 관한 연구(과제번호 : SC0400-2012-01)”
에 의하여 연구되었음.
* 교신저자 Corresponding author: Tel: +82-70-4870-4400, Fax: +82-53-959-4551, E-mail: [email protected]
ABSTRACT
As an alternative to large-scale clear cutting silviculture, strip clear cutting (SC) is being considered as a system compatible with ecological conservation and forest regeneration. In South Korea, application and effectiveness of SCC in varying forest types were rarely found. In this study, under the subject of strip clear cutting lands of pinus densiflora forest at Samcheok, Gangwon-do Province, the developmental aspect of low vegetation prior to and after deforestation and the correlation between environmental factor and pine regeneration were analyzed. The cover rate of understory vegetation was appeared to be increased after
Artemisia keiskeana, and Rubus crataegifolius that influence the cover rate showed the inclination of continuous growth. The diversity of species showed increment inclination as well due to introduction and settlement of early transient species. As a result of analyzing the correlation between vegetation and environmental factor and generation of pine tree size, the soil exposure rate, intensity of light, and canopy openness showed positive relationship, and the understory vegetation cover and woody debris cover rate showed negative relationship.
KEY WORDS: Pinus densiflora, PLANT DIVERSITY, REGENERATION, STRIP CLEAR CUTTING, UNDERSTORY VEGETATION
서 론
대상 개벌 (strip clear cutting)은 대면적 개벌에 대한 하 나의 대안으로서 (Mizoue et al., 2002), 생산림지의 생태 조건 보전과 임분 재생을 모두 고려하는 작업법이다 (Ito et al., 2006). 그러나 우리나라에서는 활엽수 또는 침엽수 이차림에 대한 대상 벌채 기법의 활용과 결과에 대한 연구 를 찾아보기 어렵다.
벌채 및 간벌과 같은 인간 활동은 산림생태계의 가장 중 요한 요소인 교목 종의 구조와 구성을 급격하게 변화시키 며, 직․간접적인 과정을 통해 생태계 기능을 변화시킨다. 그 리고 이러한 산림 활동은 대부분의 식물다양성이 존재하는 하층식생의 구조와 종조성에 큰 영향을 미치게 된다 (Halpern and Spies, 1995; Cho et al., 2011).
세계적으로 생물 및 물리 요인의 변화 결과를 고려하여 다양한 연령구조의 임분을 조성하기 위한 산림 벌채 및 재 생 기법에 대한 평가들이 이루어지고 있다 (Griffis et al., 2001; Klenner and Sullivan, 2003; Ito et al., 2006; Cho et al., 2009a; Cho et al., 2009b; Kim, 2014). 또한 산림 개벌의 생물다양성 영향에 대한 활발한 연구들이 보고되고 있다 (Battles et al., 2001; Brashears et al., 2004).
본 연구는 우리나라에서 대상 개벌의 적용 및 그것의 생 태적 영향에 관한 첫 연구이다. 대상 개벌 처리는 개벌 보다 는 낮은 수준의 임분 교란이며, 처리 후 나타나는 식생의 변화 양상은 대상 개벌의 폭, 사면방향, 원 식생 조성 등에 의해 영향을 받게 된다. 한편 임분 처리 후 급격한 식생 재생은 소나무림의 재생에 어떠한 영향을 주는지 알려진 바 없다. 본 연구는 소나무 이차림을 대상으로 대상 개벌을 적용하여 1) 수관열림도 및 광량의 비생물요인의 변화, 2) 하층식생의 종조성 및 다양성 변화, 그리고 3) 소나무 어린 개체의 이입 양상을 분석하였다.
연구방법
연구 대상지는 강원도 삼척시 하장면 갈전리 (N37°23′4 7″, E128°54′00″, 해발고 약 725m)에 위치한 6영급의 소나 무림이다. 2012년 9월 고정조사구를 설치하여 대상 벌채 처리 이전의 식생 구조 및 조성, 그리고 관련 환경 요인을 측정하였다. 이후 2012년 10월 임분 처리가 이루어졌으며, 처리 후 2014년까지 측정된 모든 요인을 반복 측정하였다.
대상 벌채 처리의 규모는 10m(W) × 80m(L) (SC_10m), 20m × 70m (SC_20m), 40m × 120m (SC_40m)이다. 처리 구 내에서 5개 소방형구 (1m × 1m)로 구성된 방형구 (1m
× 5m 및 n = 57)를 일정 간격 (조사구 환경에 따라 2.5m, 5m, 10m 및 20m)으로 설치하였고, 방형구의 수는 SC_10m 에서 15개소, SC_20m에서 15개소 및 SC_40m에서 27개소 로 구성되어 있다. 각 방형구의 상단에서 수관열림도 (Canopy Openness, %) 및 광량 (estimated transmitted light, mol m-1 day-1) 측정을 위한 어안렌즈 영상을 촬영하 였다 (Nikon D90, 렌즈 Sigma 4.5 ㎜; F2.8 EX DC CIRCULAR FISHEYE). 영상은 지상 1 m에서 획득하였고, 분석은 Gap Light Analyzer v2.0 프로그램을 이용하였으며, 촬영은 직사광의 영향을 최소화 할 수 있도록 흐린 날 또는 해 뜨기 전·후 시간대에 수행하였다 (Frazer et al., 1999;
Haugo and Halpern, 2007). 각 소방형구 내에서 비생물요 인 (수관열림도, 광량, 토양나출도, 목질잔해물 및 암석노출 도) 및 생물요인 (출현 식물 종 및 피도)을 측정하였고, 5개 소방형구에서 구한 값의 평균을 하나의 방형구 값으로 하였 다. 식물의 동정은 Lee (2003), 국명 및 학명은 Korea National Arboretum and Korea Society of Taxonomy (2007)를 따랐다.
대상 벌채 처리 후 환경, 식생 변화 및 식물다양성 변화는 측정된 요인의 평균을 구한 후 비교하였다. 평균 비교는 one-way analysis of variance (ANOVA) 분석을 이용하였
Figure 1. Overview of study area (left:2012.8, center:2013.5, right:2014).
Figure 2. Total understory cover of strip clear cutting area.
다. 식물다양성은 풍부도 (richness) 및 Shannon Index (H’, Magurran, 2004)를 적용하였다. 각 식물 종의 중요치는 각 방형구 피도 총합에 대한 상대값에 100을 곱하여 계산하였다.
결과 및 고찰
1. 하층식생 피도 변화
SC_10m의 하층식생 전체종 피도는 2012년 24.4±2.2 % 에서 벌채 후 1년차인 2013년 19.8±1.8 %로 4.6% 감소한 이후 2년차인 2014년 71.7±6.3%로 51.9%가 증가하며 벌 채 이전 대비 2.9배 늘어난 것으로 분석되었다. SC_20m의 하층식생 전체종 피도는 2012년 15.3±6.7%에서 2013년 26.1±9.3%로 10.9%가 증가하였고, 2014년 56.7±4.1%로 30.5%가 증가하며 벌채 이전 대비 약 3.7배의 피도가 늘어 난 것으로 분석되었다. SC_40m의 하층식생 전체종의 피도 는 2012년 15.3±4.7%에서 2013년 26.7±8.0%로 나타나 11.4 %가 증가했고, 2014년 82.2±7.8%로 55.5% 증가하며 벌채 이전과 비교하여 약 5.4배가 늘어난 것으로 분석되었 다. 벌채 직후(2013년)에 비해 2년차(2014년)의 전체종 피
도가 급격히 증가한 것으로 나타났는데, 이러한 결과는 기 존에 생육하던 종들이 외부의 교란에 의해 즉각적으로 반응 하지는 않는다는 것을 보여준다(Figure 2).
2014년의 종별 피도를 분석한 결과 SC_10m의 경우 신 갈나무(Quercus mongolica)가 11.9±3.2%로 가장 높게 나타났 고, 고사리(Pteridium aquilinum var. latiusculum) 6.10±1.21%, 맑은대쑥(Artemisia keiskeana) 5.8±0.7%, 산딸기(Rubus crataegifolius) 5.8±1.2%, 큰기름새(Spodipogon sibiricus) 4.8±1.0% 등의 순으로 나타났다. SC_20m는 신갈나무의 피 도가 20.6±3.2% 가장 높은 값을 보이고, 고사리 8.3±1.8%, 가는잎그늘사초(Carex humilis var. nana) 4.2±1.2%, 싸리 (Lespedeza bicolor) 2.7±0.5%, 이고들빼기(Crepidiastrum denticulatum) 2.5±0.7% 등의 순으로 나타났다. SC_40m에 서는 신갈나무가 19.4±4.3%로 가장 높게 나타났고, 조록싸 리(Lespedeza maximowiczii) 14.3±3.1%, 굴참나무(Quercus variabilis) 7.1±3.6%, 고사리 7.0±1.4%, 가는잎그늘사초 5.9±1.5% 등의 순으로 높은 피도를 가지는 것으로 분석되 었다. 처리구별 종의 피도 변화 결과에서 벌채 폭에 관계없 이 신갈나무의 피도가 가장 높게 나타나는 것으로 분석되었 는데, 이것은 중층 및 하층에서 생육하던 신갈나무가 벌채 이후 맹아지의 발달로 인해 높은 피도로 번성한 결과로 판 단된다. 이러한 참나무의 피도 증가는 벌채지의 임분재생에 중요한 역할을 하는 것으로 생각되나 소나무림의 갱신을 위해서는 지속적인 관리가 이루어져야 할 것으로 판단된다.
한편 조록싸리, 산딸기 등과 같은 관목성 종 및 고사리, 가는 잎그늘사초 등의 초본류와 같은 광선호성 종이 벌채 이후 전체종의 피도 증가에 영향을 주는 것으로 나타났다.
2. 상대중요치 변화
SC_10m의 2014년 상대중요치는 신갈나무(10.4), 맑은 대쑥(7.0), 산딸기(6.9), 고사리(6.5), 큰기름새(6.3), 가는잎 그늘사초(5.7) 등의 순으로 나타났다. SC_20m의 2014년 상대중요치는 신갈나무(22.7), 고사리(11.0), 가는잎그늘사 초(7.9), 싸리(6.3), 큰기름새(6.1), 세잎양지꽃(5.7), 맑은대
Quercus mongolica 2.5 ±0.7 3.7 ±1.2 11.9 ±3.2 4.1 ±1.0 12.3 ±3.2 20.6 ±3.6 1.2 ±0.3 6.4 ±1.2 19.4 ±4.3 Lespedeza maximowiczii 2.1 ±1.2 0.5 ±0.3 3.4 ±1.4 1.8 ±1.2 1.7 ±0.9 2.1 ±1.2 5.0 ±1.4 6.3 ±1.8 14.3 ±3.1
Quercus variabilis 0.2 ±0.1 0.2 ±0.2 0.7 ±0.5 1.3 ±0.9 1.7 ±1.7 0.1 ±0.1 3.1 ±1.3 7.1 ±3.6
Pteridium aquilinum var. latiusculum 1.0 ±0.3 2.3 ±0.5 6.1 ±1.2 0.8 ±0.3 4.4 ±1.6 8.3 ±1.8 0.9 ±0.3 2.1 ±0.5 7.0 ±1.4 Carex humilis var. nana 0.9 ±0.2 0.7 ±0.1 4.0 ±0.9 0.9 ±0.2 1.0 ±0.3 4.2 ±1.2 1.0 ±0.2 1.3 ±0.3 5.9 ±1.5 Artemisia keiskeana 1.6 ±0.3 1.7 ±0.2 5.8 ±0.7 1.3 ±0.3 0.7 ±0.2 2.2 ±0.6 1.2 ±0.2 1.5 ±0.2 5.0 ±0.8 Spodipogon sibiricus 2.3 ±0.6 1.2 ±0.3 4.8 ±1.0 2.4 ±0.6 1.2 ±0.3 2.5 ±0.4 1.9 ±0.3 1.6 ±0.3 4.5 ±1.1 Lespedeza bicolor <0.1 2.0 ±0.7 3.3 ±0.6 0.2 ±0.1 1.0 ±0.4 2.7 ±0.5 0.2 ±0.1 0.7 ±0.2 3.5 ±1.0 Rubus crataegifolius 0.1 ±0.1 0.4 ±0.1 5.8 ±1.2 0.2 ±0.2 0.1 ±0.1 1.2 ±0.5 0.3 ±0.1 0.4 ±0.2 3.0 ±1.2
Fraxinus sieboldiana 1.9 ±0.9 0.5 ±0.2 1.2 ±0.5 <0.1 0.3 ±0.1 1.1 ±0.6 1.2 ±0.5
Patrinia villosa 0.4 ±0.1 2.3 ±0.4 0.1 ±0.1 1.2 ±0.5 0.1 ±0.0 1.2 ±0.4
Crepidiastrum denticulatum 2.1 ±0.7 2.5 ±0.7 1.0 ±0.3
Corylus heterophylla 1.2 ±0.5 0.2 ±0.1 1.2 ±0.7 1.0 ±0.8 0.2 ±0.1 1.2 ±0.7 0.7 ±0.2 0.1 ±0.1 0.9 ±0.4 Potentilla freyniana <0.1 0.8 ±0.1 2.4 ±0.4 <0.1 0.6 ±0.1 1.7 ±0.3 0.0 ±0.0 0.3 ±0.1 0.8 ±0.2
Rubus phoenicolasius 0.4 ±0.2 0.1 ±0.0 0.8 ±0.5
Quercus dentata <0.1 0.5 ±0.3 0.3 ±0.1 0.1 ±0.1 0.8 ±0.8
Bidens tripartita 0.7 ±0.7
Symplocos chinensis f. pilosa 7.1 ±2.1 1.3 ±0.4 4.2 ±1.5 0.1 ±0.0 0.1 ±0.1 0.2 ±0.2 0.1 ±0.0 0.7 ±0.7
Aster scaber 0.1 ±0.0 0.2 ±0.1 0.9 ±0.4 0.2 ±0.1 <0.1 0.1 ±0.1 0.1 ±0.0 <0.1 0.7 ±0.3
Lysimachia clethroides <0.1 <0.1 0.9 ±0.2 <0.1 0.3 ±0.2 0.1 ±0.1 0.7 ±0.3
Calamagrostis arundinacea 0.2 ±0.1 0.1 ±0.1 0.1 ±0.1 <0.1 0.1 ±0.1 0.2 ±0.1 0.1 ±0.0 0.6 ±0.3 Atractylodes ovata 0.3 ±0.1 0.2 ±0.1 0.9 ±0.3 0.2 ±0.1 <0.1 0.3 ±0.2 0.2 ±0.1 0.1 ±0.0 0.4 ±0.1
Artemisia stolonifera 0.1 ±0.1 <0.1 0.2 ±0.2 0.1 ±0.0 <0.1 0.4 ±0.2
Fraxinus rhynchophylla 0.9 ±0.7 1.0 ±0.7 2.0 ±1.1 <0.1 <0.1 0.3 ±0.3
Ampelopsis brevipedunculata 0.1 ±0.1 <0.1 <0.1 <0.1 <0.1 <0.1 0.3 ±0.3
Pinus densiflora 0.1 ±0.1 0.1 ±0.0 <0.1 0.2 ±0.1 <0.1 0.2 ±0.1
Rhus tricocarpa 0.1 ±0.0 0.1 ±0.1 0.8 ±0.4 0.1 ±0.1 0.2 ±0.1 1.2 ±0.8 0.1 ±0.1 0.3 ±0.2 0.2 ±0.2
Dioscorea nipponica <0.1 <0.1 0.1 ±0.1 <0.1 0.1 ±0.0 0.1 ±0.1
Commelina communis <0.1 0.5 ±0.2 0.2 ±0.1 0.1 ±0.0 0.1 ±0.1
Astilbe rubra 0.1 ±0.1 0.2 ±0.1 1.3 ±0.6 <0.1 0.1 ±0.1
Kalopanax septemlobus <0.1 0.2 ±0.2 <0.1 0.1 ±0.1
other species 1.5 ±0.0 1.9 ±0.0 4.8 ±0.0 1.3 ±0.0 0.6 ±0.00 1.9 ±0.1 1.2 ±0.0 0.7 ±0.0 0.6 ±0.0 Total 24.4 ±9.2 19.8 ±7.2 71.7 ±22.1 15.3 ±6.7 26.1 ±9.3 56.7 ±16.5 15.3 ±4.7 26.7 ±8.0 82.2 25.4
쑥(5.2), 이고들빼기(5.2) 등의 순이었다. SC_40m의 2014 년 상대중요치는 신갈나무(15.3), 조록싸리(12.1), 가는잎그 늘사초(7.7), 고사리(7.4), 큰기름새(7.0), 맑은대쑥(7.0), 굴 참나무(5.3), 싸리(4.9), 산딸기(3.9), 세잎양지꽃 등의 순이 었다. 조사구별 상대중요치를 분석한 결과 잠재 우점종인 신갈나무의 맹아발생으로 인한 번성, 광선호성 종들의 피도 증가로 인해 상대중요치가 높아진 것으로 분석되었다.
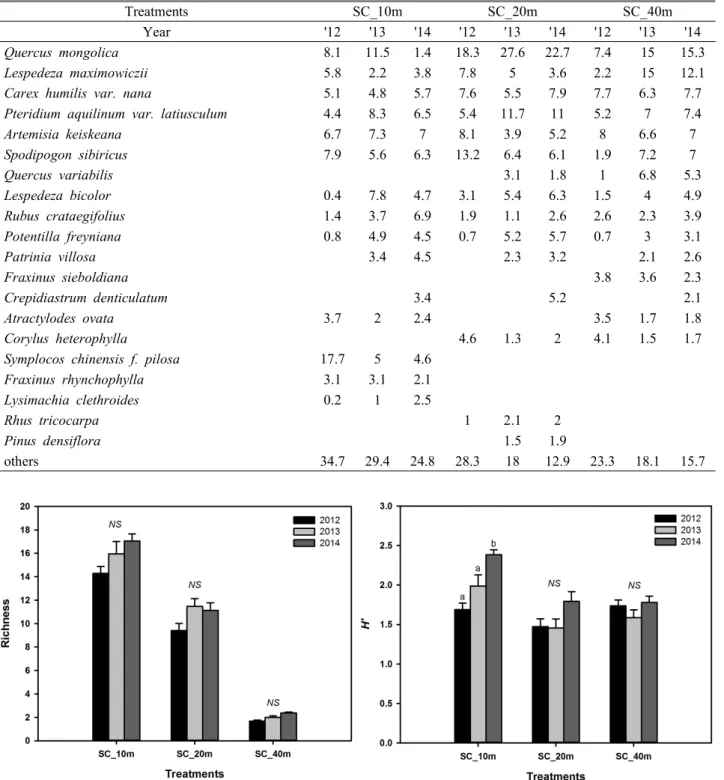
3. 종다양성 변화
소나무림 개벌 후 하층식생의 종다양성 변화를 파악하기 위해 종풍부도 및 종다양도를 분석한 결과는 Figure 6과 같다. 벌채 후 초기 종풍부도는 벌채 후 유의한 차이를 보이
지 않았지만 지속적으로 증가하는 경향을 보이는 것으로 분석되었다. 종다양성에서도 벌채 이후 증가하는 경향을 보 였으나 SC_10m에서만 유의한 결과를 보이고 있었다. 종풍 부도 및 종다양도의 지속적인 증가 경향은 숲가장자리와 인 접한 대상지의 특성으로 인해 벌채 후 두릅나무, 닭의장풀 등 천이 초기 종의 이입과 정착으로 인한 결과로 판단된다.
4. 식생 및 환경인자와 소나무 치수 발생과의 상관관 계 분석
소나무 치묘 발생량에 영향을 미치는 인자를 분석하기 위해 하층식생 피도(Unverstory cover), 토양노출도(Bared soil), 암석노출도(Rock), 목질잔해물피도(Woody debris),
Figure 3. Changes of Species diversity(left : richness, right : H’)
Treatments SC_10m SC_20m SC_40m
Year '12 '13 '14 '12 '13 '14 '12 '13 '14
Quercus mongolica 8.1 11.5 1.4 18.3 27.6 22.7 7.4 15 15.3
Lespedeza maximowiczii 5.8 2.2 3.8 7.8 5 3.6 2.2 15 12.1
Carex humilis var. nana 5.1 4.8 5.7 7.6 5.5 7.9 7.7 6.3 7.7
Pteridium aquilinum var. latiusculum 4.4 8.3 6.5 5.4 11.7 11 5.2 7 7.4
Artemisia keiskeana 6.7 7.3 7 8.1 3.9 5.2 8 6.6 7
Spodipogon sibiricus 7.9 5.6 6.3 13.2 6.4 6.1 1.9 7.2 7
Quercus variabilis 3.1 1.8 1 6.8 5.3
Lespedeza bicolor 0.4 7.8 4.7 3.1 5.4 6.3 1.5 4 4.9
Rubus crataegifolius 1.4 3.7 6.9 1.9 1.1 2.6 2.6 2.3 3.9
Potentilla freyniana 0.8 4.9 4.5 0.7 5.2 5.7 0.7 3 3.1
Patrinia villosa 3.4 4.5 2.3 3.2 2.1 2.6
Fraxinus sieboldiana 3.8 3.6 2.3
Crepidiastrum denticulatum 3.4 5.2 2.1
Atractylodes ovata 3.7 2 2.4 3.5 1.7 1.8
Corylus heterophylla 4.6 1.3 2 4.1 1.5 1.7
Symplocos chinensis f. pilosa 17.7 5 4.6
Fraxinus rhynchophylla 3.1 3.1 2.1
Lysimachia clethroides 0.2 1 2.5
Rhus tricocarpa 1 2.1 2
Pinus densiflora 1.5 1.9
others 34.7 29.4 24.8 28.3 18 12.9 23.3 18.1 15.7
Table 2. The changes of relative importance value(RIV) of species
수관열림도(Canopy openness), 광량(Transmitted lights), 종풍부도(Richness), 종다양도(Species diversity)를 매개변 수로 이용하여 중회귀분석을 수행하였다. 그 결과 소나무치 묘 발생량에 영향을 주는 인자는 토양노출도, 광량, 수관열
림도, 목질잔해물피도, 하층식생피도 순으로 나타났다. 이 러한 결과로 미루어 볼 때, 하층식생 및 목질잔해물의 피도 가 낮고 광량과 토양노출도, 수관열림도가 높아질수록 치수 발생량이 높아지는 것으로 생각된다.
Medel coefficients t p-value
1 B Std. Error Beta
(Constant) -16882.605 2950.327 6078.071 -5.722 .000 Understory cover -20.476 6.577 1043.858 -3.113 .000 Bared soils 6078.071 594.388 559.383 10.226 .000 Woody debris -241.992 82.622 -241.992 -2.929 .007 Canopy openness 559.383 102.888 -20.476 5.437 .004 Transmitted lights 1043.858 112.418 -16882. 9.286 .000 a. Dependent variable : pine seedlings
Pine seedligs = -16,882 + 6,078×토양노출도 + 1,043×
광량 + 559×수관열림도 – 20×하층식생피도 – 242×목질 잔해물피도(R-square : 0.974)
감사의 글
본 연구는 국립산림과학원 일반연구과제 “주요 경제수종 의 천연갱신 기법 및 수확작업에 관한 연구 (과제번호 : SC0400-2012-01)”의 지원에 의해 수행되었습니다.
REFRENCES
Battles, J.J., Shlisky, A.J., Barrestt, A.J., Heald, R.C., Allen-Diaz, B.H. (2001) The effects of forest management on plant species diversity in a Seirran conifer forest. Fores Ecology and Management 146: 211-222.
Brashears, M.B., Fajvan, M.A., Schuler, T.M. (2004) As assess- ment of canopy stratification and tree species diversity follow- ing clearcutting in central Appalachaian hardwoods. Forest Science 50: 54-64.
Cho YC, Kim JS, Lee CS, Cho HJ, Bae KH.(2009a) Initial Responses of Quercus serrata Seedlings and Forest Understory to Experimental Gap Treatments. Journal of Ecology and Field Biology 32: 87-96. (in Korean)
Cho YC, Kim JS, Lee JH, Lee HH, Ma HS, Lee CS, Cho HJ, Bae KH.
(2009b) Early responses of planted Quercus serrata seedlings and understory vegetation to artificial gap treatments in black locust plantation. Journal of Korean Forest Society 98: 94-105.
(in Korean)
Cho, Y.C., Pee, J.H., Kim, K.S., Koo, B.Y., Cho, H.J., Lee, C.S.
(2011) Ecological responses of natural and planted forests to thinning in southeastern Korea: a chronosequence study.
Journal of Ecology and Environment 34: 347-355.
Frazer, G.W., Canham, C.D. and Lertzman, K.P. (1999) Gap light
analyzer (GLA), version 2.0: imaging software to extract can- opy structure and gap light transmission indices from true-col- our fisheye photographs, users manual and program documentation. Simon Frazer University, Burnaby, BC, and the Institute of Ecosystem Studies, Millbrook, NY. pp. 40.
Griffis, K.L., Crawford, J.A., Wagner, M.R., Moir, W.H. (2001) Understory response to management treatments in northern Arizona ponderosa pine forests. Forest Ecology and Management 146: 239-245.
Halpern CB, Spies TA. 1995. Plant species diversity in natural and managed forests of the pacific Northwest. Ecological Applications 5: 913-934.
Haugo RD, Halpern CB. (2007) Vegetation responses to conifer en- croachment in a dry, montane meadow: a chronosequence approach. Canadian Journal of Botany 85: 285-298.
Ito S, Ishigami S, Mizoue N, Buckley GP. (2006) Maintaining plant species composition and diversity of understory vegetation un- der strip-clearcutting forestry in conifer plantations in Kyushu, southern Japan. Forest Ecology and Management 231:
234-241.
Klenner, W., Sullivan, T.P. (2003) Partial and clearcut harvesting of high-elevation spruce-fir forests: implication for small mam- mal communities. Canadian Journal of Forest Research 33:
2283-2296.
Korea Nature Aboretum, The Korean Society of Plant Taxonomists.
(2007) Korean Plant Names Index. Korea Nature Aboretum.
pp.534. (in Korean)
Magurran, A.E. (2004) Measuring Biological Diversity. Blackwell, New York.
Mizoue, N., Ito, S., I., T. (2002) Growth characteristicsof understory trees of sugi (Cryptomeria japonica) and hinoki (Chamaecyparis obtusa) in the alternate strip-clearcutting stand in Morotsuka, Miyazaki prefecture, Southern Japan. J. Jpn. For. Soc. 84, 151.158 (in Japanese ).
Lee, T.B. (2003) Cloured Flora of Korean (Ⅱ). Hyangmunsa, Seoul, 910pp. (in Korean)