B16F10 흑색종 세포에 대한 Biorenovation 기법을 이용한 파리풀 캘러스 추출물의 Melanin 생성 억제 효과
이경미1, 박태진1, 지은이2, 김석원2, 한동환1, 김승영1*
Melanogenesis Inhibitory Effects of Phryma leptostachya Callus using Biorenovation in B16F10 Melanoma Cells
Kyung-Mi Lee1, Taejin Park1, Eun Yee Jie2, Suk Weon Kim2, Dong-Hwan Han1, and Seung-Young Kim1*
Received: 27 January 2021 / Revised: 25 February 2021 / Accepted: 26 February 2021
© 2021 The Korean Society for Biotechnology and Bioengineering
Abstract: Bio-renovation is the microorganism-assisted con- version of the structure and improvement of biological effica- cies of natural products and synthetic compounds. In this study, we investigated the effect of plant callus on anti-mela- nogenic effect and compared the activity changes of bio-reno- vated callus. Phryma leptostachya bio-renovation products at the concentrations of 100, 200, 400 μg/mL suppressed tyrosi- nase activity and the expression of melanogensis-related pro- tein such as tyrosinase-related protein-1 (TRP-1), tyrosinase- related protein-2 (TRP-2) and microphthalmia-associated transcription factor (MITF) without a loss of cell viability in α-melanocyte-stimulating hormone (α-MSH)-stimulated B16F10 melanoma cells relative to its substrate. Moreover, cell viabil- ities of PLBR increased over the tested concentrations com- pared to its substrate (PL). PLBR at 400 μg/mL inhibited the α-MSH-induced melanogensis effect to the level of non-treated control. These results indicate that the structure of various compounds contained in Phryma leptostachya (PL) were modified through a bio-renovation technique and which con- ferred anti-melanogenic properties to PL.
Keywords: Phryma leptostachya, Bio-renovation, Tyrosinase activity, Whitening agent, MITF
1. INTRODUCTION
멜라닌은 멜라닌 세포에서 분출되는 흑갈색의 페놀류 고분 자 화합물로 멜라닌 분포와 양에 의해 피부색 및 눈동자 색 이 결정되며, 자외선에 의한 피부 세포의 손상을 억제하는 역할을 한다 [1-2]. 그러나 멜라닌의 비정상적인 합성 및 축 적은 노인성 흑자, 기미 및 색소침착 등의 문제를 유발한다 [3]. 자외선에 의해 자극받은 피부의 각질세포 (keratinocyte) 는 뇌하수체, 시상 하부 및 피부 세포에서 멜라닌 생성 자극 호르몬인 α-melanocyte stimulating hormone (α-MSH)신호전 달 물질을 분비하여 염증, 면역반응 등 다양한 생리 작용에 관여한다 [4-6]. 이러한 α-MSH의 활성은 cyclic adenosine monophosphate (cAMP)의 생산과 activation of protein kinase A (PKA)효소 및 tyrosinase의 활성화시켜 멜라닌 생성을 촉 진한다고 보고되었다 [7]. 멜라닌은 표피 기저층의 멜라노사 이트 (melanocyte)내의 소기관인 멜라노좀 (melanosome)에 서 단계적 효소반응에 의해 합성되며 tyrosinase, tyrosinase related protein-1 (TRP-1), tyrosinase related protein-2 (TRP-2) 등의 멜라닌 합성에 필요한 특이적인 효소들을 함유한다 [8- 10]. 이 중 tyrosinase효소는 멜라닌 색소 형성과 분비 기능을 담당하며 [11], melanogenesis의 속도결정단계인 초기반응에 작용하는 효소로서 tyrosine을 L-3,4-dihydroxyphenylalanine (L-DOPA)로 전환하는 tyrosine hydroxylase 활성과 DOPA를 도파퀴논 (DOPA-quinone)으로 산화하는 DOPA oxidase 활 성을 모두 가진 것으로 보고되었다 [12]. 따라서 tyrosine은
1선문대학교 제약생명공학과
1Department of Pharmaceutical Engineering & Biotechnology, Sunmoon University, Asan 31460, Korea
Tel.: +82-41-530-2390, Fax: +82-41-530-2939 E-mail: [email protected]
2한국생명공학연구원 생물자원센터
2Biological Resource Center, Korea Research Institute of Bioscience and Biotechnology, Jeongeup 56212, Korea.
Research Paper
L-DOPA를 거치고 산화 반응에 의해 도파퀴논으로 전환되며 자동 산화 반응을 통해 도파크롬 (dopa-chrome)을 거쳐 흑갈 색의 멜라닌이 생성되게 되며 이것은 DOPA-chrome tautmerase 로 작용하는 TRP-2에 의해 5,6-dihydroxyindole-2-carboxylic acid (DHICA)으로 전환된다 [13-15]. 최종적으로 DHICA는 TRP-1에 의해 carboxylated indole quinone으로 산화하여 흑 갈색의 eumelanin으로 전환된다 [16-17]. 따라서 멜라닌 색소 를 형성하는 효소인 tyrosinase의 활성 저해를 확인함으로써 소재의 미백효과를 검증할 수 있다.
파리풀 (Phryma leptostachya)은 파리풀과의 여러해살이풀 로서 우리나라 각처의 산과 들에서 자라며 일본, 중국, 히말 라야 산맥, 동시베리아 등지에 분포한다. 뿌리를 찧어 벌레 물린 곳에 붙이거나 즙을 내어 바르면 해독하는 효능이 있어 오래전부터 해독제와 살충제로 사용되며 피부 종기 및 부스 럼 치료 등에 이용되어왔다 [18]. Lignans 류의 haedoxan J, phrymarolin III, phrymarolin IV, leptostachyol acetate, haedoxan A 등의 성분을 함유한다고 알려져 있으며 lignans은 주로 식 물에 존재하는 폴리페놀 (polyphenol) 성분으로 항산화 활성 을 갖으며 [19-20], 파리풀의 뿌리 추출물에 대한 항염, 항산 화, 미백 효능 등이 알려져있다 [21-22].
최근 다양한 기술을 통하여 천연소재의 재활용하는 연구 개발이 이루어지고 있는 가운데, 기능이 없거나 미약한 천연 소재들의 생리활성을 증진시킬 수 있는 생물전환 기술 중 하 나인 biorenovation 기법을 이용하였다 [23]. 본 연구에서는 α-MSH로 자극된 B16F10 melanoma cells에서 파리풀 callus 추출물 (PL)과 biorenovation 기법을 사용하여 생물 전환된 파리풀 callus 추출물 (PLBR)이 멜라닌 합성에 관여하는 tyrosinase, TRP-1, TRP-2 단백질 발현 및 전사인자 MITF 활 성에 미치는 영향을 조사하고 새로운 미백기능성 화장품 소 재로서의 활용 가능성을 증명하고자 한다.
2. MATERIALS AND METHODS
2.1. 파리풀 추출물 제조
본 연구에서 사용된 파리풀 [Phryma leptostachya] callus는 한국생명공학연구원 생물자원센터로부터 제공받았다. 효율 적인 생장조절을 위해서 0.318% (w/v) Schenk & Hildebrandt, 0.1% (w/v) myo-inositol, 3% (w/v) sucrose, 0.4% (w/v) gelrite (Duchefa Biochemie, Haarlem, Netherlands), 5.0 mg/L thiamine HCl, 5.0 mg/L nicotinic acid, 0.5 mg/L pyridoxine HCl, 0.5 mg/L 2,4-dichlorophenoxyacetic acid (2,4-D), 2.0 mg/L para-chlorophenoxyacetic acid (P-CPA), 0.1 mg/L kinetin (Sigma- Aldrich, St. Louis, MO, USA)가 첨가된 배지를 사용하여 6개 월간 한 달 주기로 계대배양 하였으며 24
oC 조건에서 암배양 하였다 [24, 25]. 이를 건조 분쇄한 1 g에 대한 10배수 증류수를 첨가하고 열수 추출하였다. 추출액은 paper filter (ADVANTEC, Tokyo, Japan)로 여과한 후 감압 농축하고 −110
oC에서 동결 건조하여 분말화 하였다. Dimethyl sulfoxide (DMSO, Sigma-
Aldrich, St. Louis, MO, USA)를 용매로 사용하여 100 mM stock을 제조하였고 이후 Dulbecco’s Modified Eagle’s medium (DMEM, Welgene, Gyeongsan, KOR)으로 희석하여 100, 200, and 400 µg/mL 농도로 실험을 진행하였다.
2.2. 미생물 배양 및 biorenovation 반응
본 연구에 사용된 미생물은 Bacillus amyloliquefaciens (BA, KCTC 43033)균주로 미생물센터에서 분양받았다. Nutrient broth (0.3% beef extract, 0.5% peptone (Thermo Fisher Scientific, Hants, UK))에서 37
oC, 200 rpm으로 18시간 동안 배양하고 4,500 rpm으로 15 min 동안 centrifuge 하여 미생물 pellet을 수득한 후 PG buffer (50 mM Phosphate buffer, 2% Glycerin) 로 3회 세척하였다. PG buffer에 미생물 pellet을 현탁하여 대 조군 (BA)으로 설정하였고 여기에 파리풀 건조 시료 100 mg 을 혼합하여 30
oC에서 72h동안 생물전환 하였다. 이후 원심 분리하여 얻은 상등액을 감압 농축하고 –110
oC에서 동결건 조하여 분말화 한 후 실험에 사용하였다.
2.3. Biorenovation 전환물의 HPLC 분석
HPLC분석을 위해서 Shimadzu SpectroMonitor 3200 digital UV/Vis detector와 Shim-pack GIS C18 Column (5 μm ODS, 250
× 4.6 mm id) 을 사용하였다. 이동상 용매는 0.1% trifluoroacetic acid (TFA)를 함유한 water와 acetonitrile을 사용하였다. 시료 10 μl를 주입하여 40
oC 온도에서 gradient 조건의 1.0 mL/min 유속과 254 nm의 파장으로 40분 동안 분석하였다.
2.4. 세포배양
본 연구에서 사용된 murine B16F10 melanoma cell은 ATCC 사 (American Type Cell Culture, USA)에서 분양받아 DMEM 배양액에 10% fetal bovine serum (FBS, Welgene, Gyeongsan, KOR) 와 100 units/mL penicillin-streptomycin (P/S, Welgene, Gyeongsan, KOR) 을 첨가한 배지를 사용하여 37
oC, 5% CO
2조건에서 배양하였다.
2.5. 세포 생존율 측정
시료 처리 시 B16F10 세포의 생존율을 확인하기 위해서 24 well plate 에 B16F10 cell을 1.0 × 10
4cells/well 로 분주하고 37
oC, 5% CO
2조건의 incubator에서 24시간 전 배양하였다.
이후 100, 200, 400 µg/mL 농도의 PL, PLBR, BA를 각각 처 리하고 α-MSH (200 nM, Sigma-Aldrich, St. Louis, MO, USA)를 동시 처리하여 72시간 배양하였다. 2 mg/ml 3-(4,5- dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT, (Sigma-Aldrich, St. Louis, MO, USA)) 시약을 첨가하여 2시간 동안 반응시키고 염색된 formazan 결정을 DMSO로 용해시켜 96 well plate로 옮겨준 후 ELISA reader (multiwell microplate reader)를 사용하여 570 nm에서 흡광도를 측정하였다.
2.6. Melanin contents 측정
6 well plate에 B16F10 cell을 4.0 × 10
4cells/well로 분주하고,
37
oC, 5% CO
2조건의 incubator에서 24시간 전 배양한 뒤 시 료와 α-MSH (200 nM)을 동시처리하고 96시간 배양하였다.
Phosphate buffered saline (PBS buffer, Sigma-Aldrich, St.
Louis, MO, USA) 로 1회 세척한 후에 0.5 × trypsin-EDTA (Gibco, Grand Island, NY, USA)으로 세포를 수거하였다.
13000 rpm에서 3 min동안 centrifuge하여 얻은 pellet에 1%
protease inhibitor (Sigma-Aldrich, St. Louis, MO, USA)를 함 유한 radioimmunoprecipitation assay buffer (Sigma-Aldrich, St. Louis, MO, USA) 를 100 μL씩 첨가하고 원심분리하였다.
수득한 pellet에 10% DMSO를 함유한 1 N NaOH를 500 μl 넣 고 90
oC의 heating block에서 1시간 동안 용해시켰다. 이후 ELISA reader 를 사용하여 405 nm에서 흡광도를 측정하였다.
2.7. Tyrosinase activity 측정
6 well plate에 B16F10 cell을 4.0 × 10
4cells/well로 분주하고, 37
oC, 5% CO
2조건의 incubator에서 24시간 전 배양 한 후 시 료와 α-MSH (200 nM)을 동시에 처리하고 72시간 배양하였 다. 이후 상기의 melanin 실험과 동일한 방법으로 lysis하고 4
oC, 13000 rpm에서 30분 동안 원심분리하여 상층액을 사용 하였다. BCA kit (Thermo Scientific, USA)를 이용하여 단백 질 정량하였고, 100 mM sodium phosphate buffer (pH 6.8)와 2 mg/ml L-DOPA (Sigma-Aldrich, St. Louis, MO, USA) 를 첨 가하여 37
oC에서 2시간 반응시켰다. 이후 ELISA reader를 사 용하여 490 nm에서 흡광도를 측정하였다.
2.8. Western Blot analysis
6 well plate 에 B16F10 cell을 4.0 × 10
4cells/well 로 분주하고, 37
oC, 5% CO
2조건의 incubator에서 72시간 전 배양 하였다.
이후 시료와 α-MSH (200 nM)을 동시처리하고 24시간 후 세 포를 수거하였다. 수득한 세포에 세포 용해액 [1 × RIPA buffer, 100 mM phenylmethylsulfonyl fluoride (PMSF), 200 mM Na
3VO
4및 protease inhibitor] 100 μL를 첨가하고 4
oC, 13000 rpm에서 30분 동안 원심분리하여 상층액을 얻었다. BCA kit (Thermo Scientific, USA)를 사용하여 단백질 정량하였고 2 ×
Laemmli sample buffer (65.8 mM Tris-HCl, pH 6.8, 2.1%
SDS, 26.3% (w/v) glycerol, 0.01% bromophenol blue)와 동량 혼합하여 10% SDS-PAGE gel에 전기영동한 후 poly-vinylidene difluoride (PVDF) membrane (Bio-rad, USA) 에 전이시켰다.
상온에서 1시간 동안 5% skim milk를 함유한 TBST (10% 10
× TBS, 1% tween 20)에 blocking시킨 다음 TBST를 사용하여 10분 간격으로 4회 세척하였고 1차 항체인 TRP-1, TRP-2, tyrosinase (1:1,000, Santa cruz Biotechnology, USA), MITF (1:500, Santa cruz Biotechnology, USA) 를 처리하여 4
oC 에서 overnight 하였다. 반응이 끝난 후 TBST로 4회 세척한 후 2차 항체인 horseradish peroxidase (HRP)가 결합된 anti-mouse IgG 을 (1 : 1,000) 사용하여 상온에서 1시간 반 동안 반응시 켰다. 다시 TBST로 4회 세척한 후 ECL kit (Bio-Rad, USA)를 사용하여 imaging densitometer (model GS-700, Bio-rad, USA)을 통해 측정하였다. 측정 후 β-actin 대비 MITF, TRP1, TRP2, Tyrosinase 단백질 발현량에 대한 면적값을 imageJ program (NIH, Bethesda, MD, USA) 을 이용하여 수치화한 뒤 그래프로 나타내었다.
2.9. 통계처리
본 연구의 모든 실험은 3회 반복하여 진행하였다. 결과 값은 평균값 ± 표준편차로 나타내었으며 통계적 분석은 분산분석 (analysis of variance, ANOVA)후 student’s t-test로 다중비교 를 실시하여 각 처리군 간의 유의성 (p < 0.05, 0.01, 0.001)을 검증하였다.
3. RESULTS AND DISCUSSION
3.1. Biorenovation 전환물의 HPLC 분석
생물 전환에 이용한 미생물 Bacillus amyloliquefaciens (BA) 을 대조군으로 하여 파리풀 callus 추출물 (PL)과 생물 전환 된 파리풀 callus 추출물 (PLBR)을 HPLC 분석을 통해 비교 하였다. 분석 결과 PLBR의 19-20 min에서는 PL에서 보이지
Fig. 1. HPLC analysis of Phryma leptostachya callus extract (PL), Phryma leptostachya callus biorenovated Extract (PLBR) and
Bacillus amyloliquefaciens (BA). The chromatogram showed PL, PLBR, and BA.
않았던 신규 peak가 발견되었다 (Fig. 1). 이는 미생물을 이용 한 생물전환 과정에서 PL과의 생화학적인 반응에 의하여 성 분 변화가 유도되었을 가능성을 시사한다. 이후 성분 변화에 따른 생리학적 활성 변화를 알아보고자 B16F10 melanoma cell에서 실험을 진행하였다.
3.2. 세포 생존율 측정
미토콘드리아의 reductase에 의하여 MTT 시약은 보라색의 formazan 으로 전환되며 formazan의 양은 생존 가능한 세포 의 수와 비례한다 [26]. B16F10 cell에 PL과 PLBR 그리고 생 물전환 반응에 사용한 균 (BA)을 100, 200, 400 µg/mL의 농 도로 처리하여 세포 생존율을 확인한 결과 측정 농도에서 PL, PLBR 및 BA 모두 85.1 ± 4.3% 이상의 생존율을 보였다 (Fig. 2). 따라서 추후 실험에서는 추출물 처리 군에 대해 세 포의 성장 저해 및 사멸을 보이지 않는 400 µg/mL을 최대 농 도로 설정하여 실험을 진행하였다.
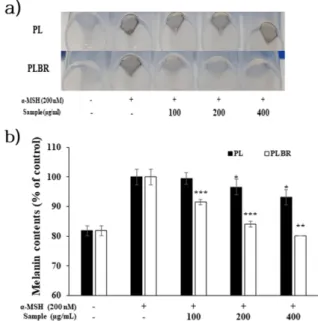
3.3. Melanin contents 함량 측정
α-MSH (200 nM)로 자극된 B16F10 cell에 PL 및 PLBR 추출 물을 농도별로(100, 200, 400 µg/mL) 처리하여 멜라닌 합성 에 대한 저해 활성을 조사하였다. 실험결과 α-MSH 자극에 의해 세포 내 멜라닌이 축적되었으며 PL 처리군은 이를 억 제하지 못하여 pellet 색이 검게 변한 것이 관찰되었다. 반면 PLBR 처리군에서는 농도 의존적으로 멜라닌 축적을 저해하 여 최고농도인 400 µg/mL에서는 α-MSH 무 처리군과 유사 한 pellet 색을 나타내었다 (Fig. 3(a)). 또한 흡광도 측정 결과 PL 처리군 보다 PLBR 처리군에서 더 우수한 멜라닌 합성 저 해 활성이 확인되었다 (Fig. 3(b)). 이는 생물전환 과정에서 미 백활성을 가진 유효물질이 생성되어 멜라닌 합성을 저해한 것 으로 사료된다. 따라서 세포 밖으로 분비되는 melanogenesis에 관여하는 하위인자들의 발현을 알아보기 위하여 추가적인 실험을 진행하였다.
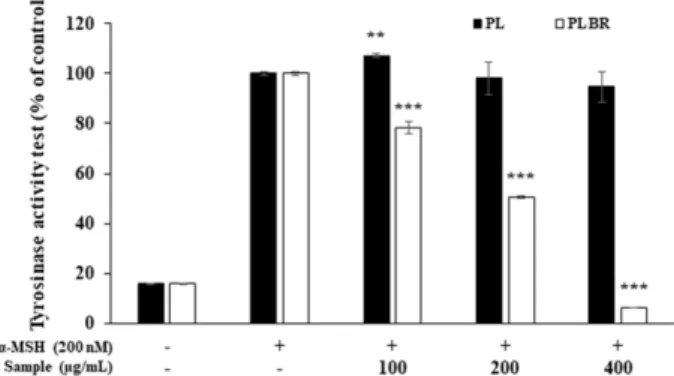
3.4. Tyrosinase activity 측정
Tyrosinase는 멜라닌 생성 초기반응에 관여하는 효소로서 L- DOPA와의 반응에 의하여 dopa-chrome을 형성하고 이를 흡 광도로 측정함으로써 dopa oxidase 활성 및 tyrosinase 저해 활성을 평가한다 [27]. α-MSH 자극에 의해서 tyrosinase activity 발현을 유도하고 PL 및 PLBR을 처리한 결과 α-MSH 단독 처리군과 비교하여 PL은 최대농도인 400 μg/mL에서 tyrosinase 효소 활성이 5.3 ± 6.0% 억제된 것에 비해 PLBR은 같은 농도에서 93.9 ± 0.1%의 tyrosinase activity 저해를 나타 내며 농도 의존적으로 우수한 활성을 보였다 (Fig. 4). 이는 biorenovation 생물전환체에 의해 tyrosinase 저해 활성이 크 게 향상된 것으로 사료되며, 이러한 tyrosinase 저해는 멜라 닌 합성이 억제된 주요 원인으로 판단된다.
3.5. 멜라닌 합성 관련인자 발현 측정
α-MSH 자극으로 유도된 B16F10 melanoma cell에 PL 및 PLBR 처리 시 melanin 합성 하위기전인 TRP-1, TRP-2, tyrosinase 단백질 발현량과 전사인자 MITF 활성에 미치는 영향을 조사하였다. 그 결과 α-MSH 자극에 의하여 무 처리 군에 비해 melanin합성 관여인자의 발현이 증가된 것을 확인 하였으며 PL 처리 시 멜라닌 합성 관여인자들의 단백질 발 현량이 일관된 저해를 보이지 않지만 PLBR 처리 시 농도 의 존적으로 단백질 발현을 저해하였고 200 μg/mL 이상의 농도 에서는 TRP-1, TRP-2, tyrosinase 및 MITF 발현량이 α-MSH Fig. 2. Cell viability of PL, PLBR and BA on B16F10 melanoma
cell by the MTT assay. B16F10 cells (1 × 10
4cells) were started in medium for 24 h. After the cells were treated α-MSH (200 nM) with various concentrations of PL and PLBR extract (100, 200, and 400 μg/mL) for 72 h. Result is expressed as percentages compared to the respective values obtained for the control. The values represent mean ± SD. * p < 0.05.
Fig. 3. Inhibitory effects of PL and PLBR extract on cellular
melanin synthesis in B16F10 cells. B16F10 cells (4 × 10
4cells)
were started in medium for 24 h. After the cells were stimulated
with α-MSH (200 nM) and treated with 100, 200, and 400 μg/mL
of PL and PLBR extract for 96 hours. The picture showed
accumulation of intracellular melanin (a), and melanin content
was measured by absorbance at 405 nm (b). The values represent
mean ± SD. * p < 0.05, ** p < 0.01, *** p < 0.001.
무 처리군과 비슷한 수준으로 확인되어 PLBR 추출물이 멜 라닌 합성 관여인자를 효과적으로 저해한다는 것을 확인하 였다 (Fig. 5). 이 결과는 biorenovation 기법으로 생물 전환된 PLBR 내의 유효성분이 tyrosinase avtivity 저해 뿐만 아니라 melanogensis 관련 인자들에도 직간접적인 영향을 미친 것으 로 판단된다.
4. CONCLUSION
α-MSH로 자극된 B16F10 melanoma cell에 PL과 PLBR을 처 리하여 세포 생존율 및 미백 활성을 조사하였다. HPLC 분석 을 통해 기존 소재 (PL)와 비교하여 생물 전환된 추출물 (PLBR) 에서 신규 피크가 생성된 것을 확인하였고 이후 세포 사멸을 보이지 않는 농도 (100, 200, 400 μg/mL) 내에서 멜라 닌 생성 기작에 관여하는 효소 및 단백질의 발현량을 확인하 Fig. 4. Inhibitory effects of PL and PLBR extract on tyrosinase
activity in B16F10 cells. B16F10 cells (4 × 10
4cells) were started in medium for 24 h. After the cells were stimulated with α-MSH (200 nM) and treated with 100, 200, and 400 μg/mL of PL and PLBR extract for 72 hours. Tyrosinase activity was measured by absorbance at 490 nm. The values represent mean ± SD. * p < 0.05, ** p < 0.01, *** p < 0.001.
Fig. 5. Western blot analysis of the expression of MITF, TRP-1, TRP-2, tyrosinase in B16F10 melanoma cells. Protein expression
levels of TRP-1 (a), TRP-2 (b), tyrosinase (c), MITF (d) in α-MSH simulated B16F10 melanoma Cells. After B16F10 cells (4 × 10
4cells) were started in medium for 72 h, the cells were stimulated with α-MSH (200 nM) and treated with 100, 200, and 400 μg/mL of PL
and PLBR extract for 24 hours. Western blot analysis of cell lysates was performed using indicated antibodies. Proteins were visualized
using an ECL detection system and β-actin expression was used as a control for normalization of target proteins. The values represent
mean ± SD. * p < 0.05, ** p < 0.01, *** p < 0.001.
였다. 그 결과 PL 처리군은 melanin 합성에 대한 저해 활성이 미미한 반면 PLBR 처리군은 tyrosinase 발현을 억제하여 L- DOPA 합성을 저해하였다. 또한 MITF 전사인자의 발현을 억제함으로써 TRP-1,2 효소의 발현이 조절되었고 최종적으 로 eumelanin 합성량이 농도 의존적으로 저해되었다. 특히 200-400 μg/mL 농도에서 melanogenesis 관여인자의 발현은 무처리군 수준으로 나타났다. 이 결과는 PL의 기존 화합물 들이 미생물 내에 존재하는 효소에 의해 구조변화가 유도됨 으로써 멜라닌 생성 효소 및 단백질의 발현을 저해하는 활성 성분이 합성된 것으로 사료되며 추후 PLBR의 유효성분 분 석에 관한 연구가 수행되어야 할 것으로 판단된다. 본 연구 에서는 biorenovation 기법을 이용한 생물전환을 통해서 구 조적으로 복잡한 천연물의 생리학적 활성을 증진하였고 새 로운 미백 소재를 탐색하였으며 기능성 화장품 소재로서 PLBR의 활용 가능성을 제시한다.
Acknowledgements
본 연구는 보건복지부의 재원으로 한국보건산업진흥원의 보건의료기술연구개발사업 지원에 의하여 이루어진 것임 ( 과제고유번호 : HP20C0231).
REFERENCES
1. Quevedo, W. C., T. B. Fitzpatrick, M. A. Pathak, and K. Jimbow (1975) Role of light in human skin color variation. America Jour- nal of Physical Anthropology. 43: 393-408.
2. Hill, H. Z., W. Li, P. Xin, and D. L. Mitchell (1997) Melanin: a two edged sword?. Pigment Cell Re. 10: 158-161.
3. Fernandez, A (2015) Dermatology update: The dawn of targeted treatment. Cleve. Clin. J. Med. 82: 309-320.
4. Leiter, U., T. Eigentler, and C. Garbe (2014) Epidemiology of skin cancer. Adv. Exp. Med. Biol. 810: 120-140.
5. Jung, S. H., M. J. Ku, H. J. Moon, B. C. Yu, M. J. Jeon, and Y. H.
Lee (2009) Inhibitory Effects of Fucoidan on Melanin Synthesis and Tyrosinase Activity. Korean Society Of Life Science. 19: 75- 80.
6. Davidson, R. L (1972) Regulation of Melanin Synthesis in Mam- malian Cells: Effect of Gene Dosage on the Expression of Differ- entiation. Proc. Natl. Acad. 69: 951-955.
7. Kreiner, P. W., C. J. Gold, J. J. Keirns, W. A. Brock, and M. W.
Bitensky (1973) Hormonal control of melanocytes: MSH-sensi- tive adenyl cyclase in the Cloudman melanoma. Yale J. Biol. Med.
46: 583-591.
8. Slominski, M. R., M. A. Zmijewski, and A. T. Slominski (2015) The role of melanin pigment in melanoma. Exp Dermatol, 24:
258-259.
9. Bertolotto, C., R. Buscà, P. Abbe, K. Bille, E. Aberdam, J. P.
Ortonne, and R. Ballotti (1998) Different cis-acting elements are involved in the regulation of TRP1 and TRP2 promoter activities
by cyclic AMP: pivotal role of M boxes (GTCATGTGCT) and of microphthalmia. Mol. Cell Biol. 18: 694-702.
10. Newton, R. A., A. L. Cook, D. W. Roberts, J. Helen Leonard, and R. A. Sturm (2007) Post-Transcriptional Regulation of Melanin Biosynthetic Enzymes by cAMP and Resveratrol in Human Mela- nocytes. Journal of Investigative Dermatology. 127: 2216-2227.
11. Jeon, S. H., K. H. Kim, J. U. Koh. and K. H. Kong (2005) Inhibi- tory effects on L-dopa oxidation of tyrosinase by skin-whitening agents. Bull Korean Chem Soc. 26: 1135-1137.
12. Song, H. S., H. J. Moon, B. E. Park, B. S. Choi, D. J. Lee, J. Y.
Lee, C. J. Kim, and S. S. Sim (2007). Anti-oxidant activity and whitening activity of bamboo extracts. Yakhak Hoeji. 51: 500-507.
13. Przemyslaw, M. P., and G. Maja (2006) Melanin synthesis in microorganisms-biotechnological and medical aspects. Acta bio- chimica polonica. 53: 429-443.
14. Fitzpatrick, T. B., S. W. Becker, A. B. Lerner, and H. Montgomery (1950) Tyrosinase in Human Skin: Demonstration of Its Presence and of Its Role in Human Melanin Formation. Science. 112: 223- 225.
15. Edelstein, L. M., N. Cariglia, M. R. Okun, R. P. Patel, and D.
Smucker (1975) Inability of Murine Melanoma Melanosomal
“Tyrosinase” (L-Dopa Oxidase) to Oxidize Tyrosine to Melanin in Polyacrylamide Gel Systems. Journal of Investigative Dermatol- ogy. 64: 364-370.
16. Kovacs, D., E. Flori, V. Maresca, M. Ottaviani, N. Aspite, M.
Dell’Anna, L. Panzella, A. Napolitano, M. Picardo, and M. d’
Ischia (2012) The Eumelanin Intermediate 5,6-Dihydroxyindole2- Carboxylic Acid Is a Messenger in the Cross-Talk among Epider- mal Cells. Journal of Investigative Dermatology. 132: 1196-1205.
17. WILCZEK, A., H. KONDOH, and Y. MISHIMA (1996) Compo- sition of Mammalian Eumelanins: Analyses of DHICA-Derived Units in Pigments From Hair and Melanoma Cells. Pigment Cell Research. 9: 63-67.
18. Park, I. K., S. C. Shin, C. S. Kim, H. J. Lee, W. S. Choi, and Y. J.
Ahn (2005) Larvicidal Activity of Lignans Identified in Phryma leptostachya Var. asiatica Roots against Three Mosquito Species. J.
Agric. Food Chem. 53: 969-972.
19. Chen, C., H. Zhu, D. Zhao, and J. Deng (2012) Lignans from Phryma leptostachya L. Helvetica Chimica Acta. 95: 333-338.
20. Kitts, D. D., Y. V. Yuan, A. N. Wijewickreme, and L. U. Thomp- son (1999) Antioxidant activity of the flaxseed lignan secoisolari- ciresinol diglycoside and its mammalian lignan metabolites enterodiol and enterolactone. Molecular and Cellular Biochemis- try. 202: 91-100.
21. Yoo1, S. M., and J. H. Lee (2019) Antioxidative Effect of Phry- maleptostachyavar. Asiatica HARA Extract on the Neurotoxicity of Aluminum Sulfate, Environmental Pollutant. Korean J Clin Lab Sci. 51: 235-244.
22. Jung, H. J., Y. W. Cho, H. W. Lim, H. j. Choi, D. J. Ji, and C. J.
Lim (2013) Anti-Inflammatory, Antioxidant, Anti-Angiogenic and Skin Whitening Activities of Phryma leptostachya var. asiatica Hara Extract. Biomol Ther. 21: 72-78.
23. Park, T. j., J. H. Sim, H. H. Hong, D. H. Han, and S. Y. Kim (2020) Anti-inflammatory Effect of Colocasia esculenta Bioreno- vate Extract in LPS-stimulated RAW 264.7 Cells. KSBB Journal.
35: 162-168.
24. Kang, C. H., B. S. Han, S. G. Han, S. H. Kown, and Y. Ju. Song (2011) Selection of Suitable Varieties of Carnation (Dianthus caryophyllus L.) and Optimization of Culture Conditions for Effi- cient Tissue Culture. Korean Journal of Plant Resources. 24: 121- 129.
25. Hwang, I. T., J. J. Lee, J. Y. Lee, S. W. Paik, and Y. H. Kim (2015) Production of Baicalin, Baicalein, and Wogonin on Plant Tissue Culture of Scutellaria baicalensis. Korean Journal of Plant
Resources. 28: 526-532.
26. Van Meerloo, J., G. J. L. Kaspers, and J. Cloos (2011). Cell Sensi- tivity Assays: The MTT Assay. Cancer Cell Culture. 731: 237- 245.
27. Jin, Y. Z., S. Y. Ahn, E. S. Hong, G. H. Li, E. K. Kim, and K. H.
Row (2005) Extraction of Whitening Agents from Natural Plants and Whitening Effect. J. Korean Ind. Eng Chem. 16: 348-353.