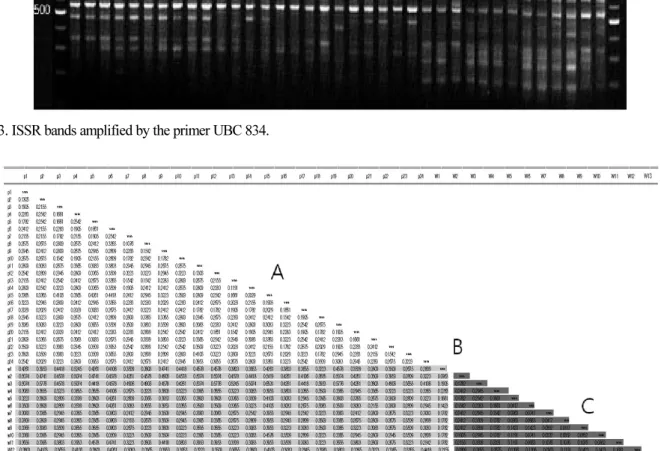
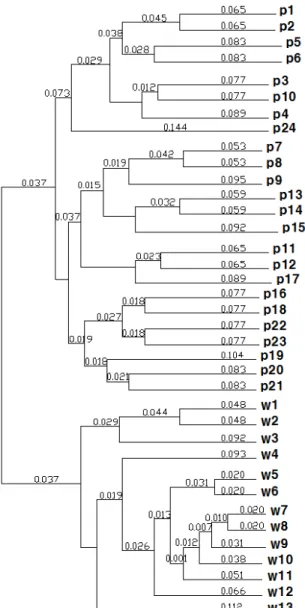
Variant Identification in Platanus occidentalis L. Using SNP and ISSR Markers
Jin Young Lee
1, Mu Seok Han
2and Chang Seob Shin
1*
1
Department of Forest Science, Chungbuk National University, Cheongju 361-763, Korea
2
전체 글
1
2
수치


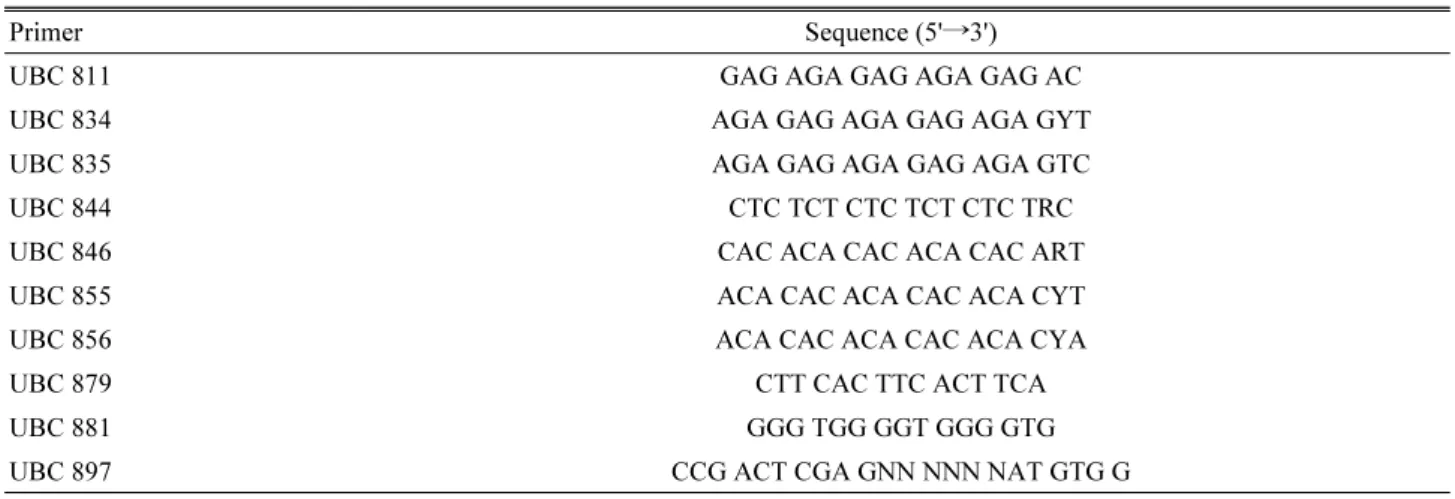
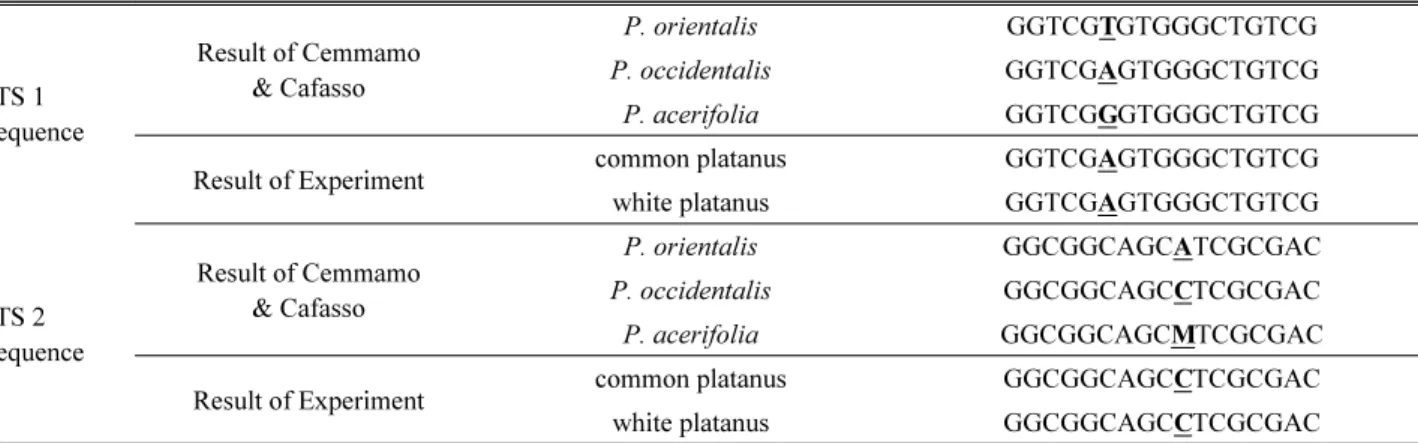
관련 문서