O.-J. Hwang ・ J.-I. Kim ( )
전남대학교 생명공학과 & 금호생명과학연구소
(Department of Biotechnology and Kumho Life Science Laboratory, Chonnam National University, Gwangju 500-757 Korea) e-mail: [email protected]
생명공학 벤트그라스 개발
황옥진 ・ 김정일
Recent advances in the development of biotech bentgrass
Ok-Jin Hwang ・ Jeong-Il Kim
Received: 24 November 2009 / Accepted: 1 December 2009
ⓒ Korean Society for Plant Biotechnology
Abstract Creeping bentgrass (Agrostis stolonifera L.) is economically important as the principal turfgrass species for golf course greens and fairways in temperate climates around the world. As the utilization area of the turfgrass species increases recently, the demand for new and improved cultivars increases. Thus, substantial progress has been made in applying modern biotechnology to develop genetically engineered (i.e., biotech) creeping bentgrass with new traits that eluded the breeders. This review article addresses the advances made in developing biotech creeping bentgrass, which are categorized in the following topics: (i) genetic transformation of creeping bentgrass, (ii) development of various biotech creeping bentgrasses by genetic engi- neering, and (iii) progresses in the deregulation of herbicide- resistant creeping bentgrass.
Keywords Creeping Bentgrass, Agrostis, Transformation, Biotech crop, genetic engineering, Deregulation
서 론
잔디는 각종 환경에 대한 적응력이 매우 강한 식물로 척박한 토양을 피복할 목적으로 많이 이용되어 왔으나, 요즈음 생활수준이 향상되면서 환경 미화와 레저 등에 대한 관심이 증가됨에 따라 그 효용성은 더욱 높아지고 있다. 특히 경제수준의 향상과 더불어 최근 증가되고 있 는 골프장이나 축구장과 같은 레저 및 스포츠산업을 비
롯한 사회 전반에서 잔디의 수요가 급격히 증가하고 있 으며, 이로 인하여 잔디관련 사업규모가 크게 확대되고 있어 고품질의 잔디 개발을 필요로 하고 있다. 미국의 경 우, 잔디 종자시장은 옥수수 다음의 위치를 점하는 등 매 우 높은 경제성을 지니고 있으며, 잔디는 유지관리 비용 이 많이 드는 작물로서 해마다 약 45억불 이상을 유지관 리 비용으로 소모하고 있다 (Lee 1996; Zilinskas and Wang 2004). 이에 생명공학 기법을 이용한 고부가가치 생명공 학 잔디 품종 개발의 필요성과 기대가 증대되고 있다.
벤트그라스 (Bentgrass)는 일반적으로 가장 품질이 좋은 다년생 한지형 잔디로 알려져 있으며, 옆폭이 매우 짧은 포복형 (stoloniferous) 잔디로 질감이 매우 곱고 3mm 정도 의 짧은 예초에도 잘 견뎌 주로 골프장의 그린이나 테니 스 코트 등에 거의 전적으로 쓰이는 경제적으로 중요한 작물이다 (Warnke 2003). 벤트그라스는 서유럽과 아시아 가 원산지이며, Agrostis 속에 속하고 약 200 여종이 알려 져 있다. 하지만 상업 용도로 개발된 것은 약 5종 정도의 소수에 불과하며, 대표적인 종으로 크리핑 벤트그라스 (Creeping bentgrass; 2n=4x=28)인 Agrostis stolonifera가 있다 (Bonos et al. 2002). 크리핑 벤트그라스는 지표면에 발달 하는 강력한 포복경과 각 마디에서 뿌리와 싹이 새롭게 발생하는 것에서 유래되어 붙여진 이름이다. 그리고 문 헌마다 크리핑 벤트그라스의 종명 사용에 차이가 존재하 는데, 예를 들면 Agrostis palustris Huds., Agrostis stolonifera L., Agrostis stolonifera L. var palustris 등이 있다. 이것은 포복경 형성 정도의 차이에 기인하며, 현재는 Agrostis
stolonifera L.이 가장 보편적으로 사용된다 (Warnke 2003).크리핑 벤트그라스의 상업적 품종으로는 1950년대 초반 부터 “Penncross”가 가장 널리 이용되어 최근까지도 많이 사용되고 있으며, 1990년 전후로 다수의 신품종이 개발 되기 시작하여 “Pennlinks” “Crenshaw” “L-93” “Penn-A-4”
등의 품종이 개발되어 사용되고 있다. 예로써, “Crenshaw”
DOI:10.5010/JPB.2009.36.4.327
Review
328 J Plant Biotechnol (2009) 36:327–335
품종은 “Penncross”보다 더 우수한 여름철 밀도와 진녹색 을 보이며 여름에 강한 잔디로 개발된 것이다 (Engelke et al. 1995).
벤트그라스는 우수한 질감과 색, 그리고 짧은 예초에 잘 견디고 골프장 그린용 잔디로서 우수한 퍼팅 품질을 갖고 있는 데 반하여, 왕성한 생장을 지님으로써 높은 시 비량과 관개를 매일 필요로 할 뿐만 아니라, 고온과 건조 조건 그리고 병충해에도 약하여 늘 세심한 관리와 주의 가 요구되는 초종이다 (Warnke 2003). 이에 이렇게 불리 한 형질들은 전통 육종 방법에 의해 많이 개량되었으나, 아직 병 저항성, 내건성, 내서성 등과 같은 형질들의 개 선은 여전히 필요하다. 따라서 최근에는 전통 육종의 한 계를 극복하기 위하여 생명공학 기법을 이용하여 우수한 품종을 개발하는 연구가 활발히 진행되고 있다 (Chai and Sticklen 1998; Wang et al. 2001; Zilinskas and Wang 2004;
Wang and Ge 2006a). 한 예로써, 벤트그라스는 잡초와의 경쟁에서 약하기에 생명공학 기법을 이용하여 잡초 관리 에 용이한 제초제 저항성 벤트그라스들이 개발되었다 (Hartman et al. 2004; Asano et al. 1997; Chai et al. 2003; Luo et al. 2004; Kim et al. 2007). 본 논문에서는 생명공학 벤트 그라스 개발 연구 현황과 동향을 이해하기 위하여, 현재 까지 확립된 벤트그라스 형질전환 시스템과 개발된 생명 공학 벤트그라스 종류, 그리고 상업화 추진 현황 등에 대 하여 정리해 보고자 한다.
본 론
벤트그라스 형질전환 시스템
크리핑 벤트그라스 (Agrostis stolonifera L.)의 안정적인 형질전환은 1993년에 microprojectile bombardment 방법으 로 처음 보고되었다 (Zhong et al. 1993). 이후 또다른 그룹 들도 동일한 방법으로 형질전환을 보고하였으며 (Hartman et al. 1994; Xiao and Ha 1997), 또한 protoplast를 분리하여 polyethylene glycol 또는 electroporation을 이용하여 유전자 를 직접 주입시키는 방법의 형질전환 방법도 보고되었다 (Lee et al. 1996; Asano et al. 1997; 1998). 이들 대부분은 제초제 저항성 유전자인 bar를 도입시키기 위해 이루어 졌다. 하지만 이들 방법들은 도입 유전자 수가 많은 점, 후세대에서 도입된 유전자가 발현되지 않는 gene silencing 문제와 낮은 성공 확률 등으로 최근에는 많이 사용되지 않고 있으며, 이러한 문제점들이 적은 Agrobacterium 매개 형질전환 방법이 주로 이용되고 있다 (Dai et al. 2001).
크리핑 벤트그라스의 Agrobacterium 매개 형질전환은 2000년에 최초로 보고되었다 (Yu et al. 2000). 이때에는 형질전환 과정에서 reporter gene으로 GFP (Green fluorescent
protein) 유전자가 이용되었고, 단 두 개의 독립 라인이 확 인되어 보고되었다. 이후 Agrobacterium 매개 형질전환 효 율을 증대시키기 위하여 여러 연구 그룹에서 보고가 잇 따랐는데, 대체적인 형질전환 과정은 비슷하나 사용하는 벤트그라스 종이나 selection marker 또는 조직배양 조건 등에서 약간씩의 차이가 있었다 (Chai et al. 2003; 2004;
Luo et al. 2004; Han et al. 2005; Aswath et al. 2005; Fu et al. 2005a; 2005b; 2007; Kim et al. 2007; Han et al. 2009). 예 를 들면, 크리핑 벤트그라스가 아니라 콜로니얼 벤트그 라스를 대상으로 한 보고가 있었으며 (Chai et al. 2003;
2004), selection marker 유전자로 bar가 아니라 hygromycin 저항 성 유전자를 사용한 보고도 있었다 (Chai 2004; Han et al.
2005). 또한 제초제 저항성이나 hygromycin 저항성을 사용 하지 않고, GFP와 manA (phosphomannose isomerase) 유전 자를 사용하는 보고도 있었다 (Fu et al. 2005a). 나머지 보 고들은 대부분 bar 유전자를 이용하여 형질전환을 수행 하였고, 일부 embryogenic callus 유도 조건이나 Agrobacterium 형질전환 과정에서 효율을 증진시키기 위한 방법의 차이 들이 있었다. 또한 최근에는 성숙종자에서 유래된 em- bryogenic callus가 아니라 stolon을 이용한 Agrobacterium 매개 형질전환 방법도 보고되었다 (Wang and Ge 2006b;
Xu et al. 2009).
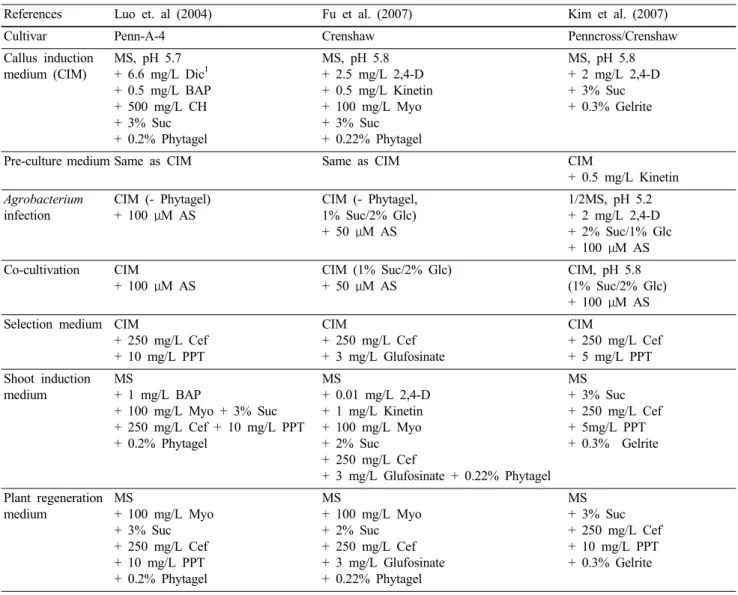
현재까지 보고된 크리핑 벤트그라스의 Agrobacterium 매개 형질전환 과정을 종합하여 최적화된 과정을 살펴보 면 Fig. 1과 같다. 간단히 살펴보면, 소독된 성숙 종자로 부터 호르몬을 첨가한 callus 유도 배지에서 embryogenic callus를 유도하고 (callus induction), 계대 배양을 통하여 좋은 embryogenic callus를 선발한다 (embryogenic callus subculture). 그리고 Agrobacterium 접종 1주일 전에 0.5 mg/L kinetin이 포함된 배지에서 배양한 후 (pre-culture),
Agrobacterium 용액에 10분 정도 접종하여 약 3일간 공동 배양시킨다 (co-cultivation). 이후 bialaphos 또는 PPT (phosphinotricin)가 함유된 배지에서 약 4-10주일간 배양하 면서 살아있는 callus들만 골라낸 후 (selection), 호르몬이 없는 shooting 유도 배지로 옮겨 재분화를 유도한다 (shoot induction & root induction). 이후 토양으로 옮겨 심어 약 2 주일간 순화시키고, 제초제 저항성 실험을 통하여 형질 전환체를 선발하게 된다. 보고된 논문마다 단계별 약간 의 기간 차이가 있긴 하지만, 대략 callus induction은 8 주~9주, transformation 후 co-culture는 약 3일, selection은 4~10주, shoot induction은 4-6주, root induction은 약 4주 정 도로 형질전환체를 얻는 데는 약 6-7개월 정도 걸리는 것 으로 파악되었다. 만약 충분한 embryogenic callus가 확보 된 상태라면 callus induction 기간이 생략되므로 형질전환 체를 약 4-5개월 내에 얻을 수 있다.
현재까지 보고된 크리핑 벤트그라스의 대표적인 Agro-
bacterium 매개 형질전환 조건들을 비교하면 Table 1과 같Fig. 1 Agrobacterium-mediated transformation procedures for creeping bentgrass. Embryogenic callus were induced from mature seeds on callus induction medium, and selected visually and pre-cultured on callus induction medium supplemented with 0.5 mg/L kinetin for 1 week prior to Agrobacterium-mediated transformation. The embryogenic calli were then immersed in Agrobacterium suspensions and incubated for 10 min with gentle shaking, followed by co-cultivation in the dark at 25°C for 3 d. After the co-cultivation, the infected calli were transferred onto a medium supplemented with 5 mg/L PPT and 250 mg/L cefotaxime, and cultured under darkness for 4-10 weeks to select PPT-resistant calli. For plant regeneration, PPT-resistant green shoots were transferred to plastic vessels containing a hormone-free MS medium with 10 mg/L PPT and 250 mg/L cefotaxime under appropriate lighting, and plantlets with well-developed roots were placed on soil and grown under greenhouse conditions
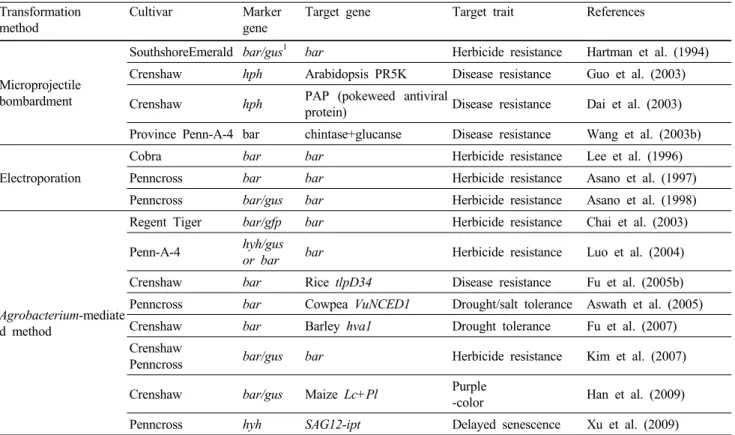
다. 전반적인 형질전환 과정들은 대부분의 연구 그룹에 서 비슷하게 진행하고 있으며, 다만 callus induction medium (CIM)이나 pre-culture 조건, 그리고 Agrobacterium 접종 및 공동배양 조건에서 차이를 보이고 있다. 현재까지 보고 된 내용들을 토대로 형질전환 과정에서 중요하다고 여 겨지는 과정들을 살펴보면, 벤트그라스에서 embryogenic callus를 얻기 위한 호르몬 조성은 2,4-D 단독처리 (Kim et al. 2007; Han et al. 2009), 2,4-D와 kinetin의 혼용처리 (Yu et al. 2000; Fu et al. 2005a; 2005b; 2007), dicamba와 BAP의 혼용 처리 (Luo et al. 2004) 등을 이용하고 있으며, 각각 다른 호르몬 조성에 의해 유도된 embryogenic callus의 비 율과 재분화율은 큰 차이를 나타내지 않는 것으로 보인 다 (Kim et al. 2007). 따라서 벤트그라스 종자로부터 em- bryogenic callus 유도 조건은 크게 중요한 것으로 여겨지 지 않으며, 다만 embryogenic callus 비율에 차이를 주는 것
으로 여겨진다. 하지만 Agrobacterium 접종 전의 pre-culture 단계는 형질전환에 중요한 영향을 미치는 것으로 여겨진 다. Luo 등 (2004)은 Agrobacterium 접종 하루 전에 100 μM acetosyringone을 전처리시키는 과정을 거치고, 또한 Kim 등 (2007)은 접종 1주일 전에 0.5 mg/L kinetin를 처리하면 형질전환 효율이 약 3배가 증대됨이 보고되었다. 그리고
Agrobacterium 접종 및 공동 배양 조건도 중요한 것으로여겨지는데, Luo 등 (2004)은 각 캘러스에 Agrobacterium 배양액을 10 μL 바로 떨어뜨리면서 접종하였고, 다른 연 구 그룹들도 탄소원 비율을 바꾼다거나 MS 배지를 half- strength로 하거나 pH를 바꾸는 등 Agrobacterium에 의한 유전자 도입 조건을 최적화 하였다 (Fu et al. 2005a; 2005b;
Kim et al. 2007; Han et al. 2009). 마지막으로 공동배양 후
유전자가 도입된 callus를 selection하는 과정이 중요한 것
으로 파악되었다. 이 과정은 형질전환된 callus에서 shooting
330 J Plant Biotechnol (2009) 36:327–335
Table 1 Comparison of Agrobacterium-mediated transformation conditions for creeping bentgrass
References Luo et. al (2004) Fu et al. (2007) Kim et al. (2007)
Cultivar Penn-A-4 Crenshaw Penncross/Crenshaw
Callus induction medium (CIM)
MS, pH 5.7 + 6.6 mg/L Dic
1+ 0.5 mg/L BAP + 500 mg/L CH + 3% Suc + 0.2% Phytagel
MS, pH 5.8 + 2.5 mg/L 2,4-D + 0.5 mg/L Kinetin + 100 mg/L Myo + 3% Suc + 0.22% Phytagel
MS, pH 5.8 + 2 mg/L 2,4-D + 3% Suc + 0.3% Gelrite
Pre-culture medium Same as CIM Same as CIM CIM
+ 0.5 mg/L Kinetin Agrobacterium
infection
CIM (- Phytagel) + 100 μM AS
CIM (- Phytagel, 1% Suc/2% Glc) + 50 μM AS
1/2MS, pH 5.2 + 2 mg/L 2,4-D + 2% Suc/1% Glc + 100 μM AS Co-cultivation CIM
+ 100 μM AS
CIM (1% Suc/2% Glc) + 50 μM AS
CIM, pH 5.8 (1% Suc/2% Glc) + 100 μM AS Selection medium CIM
+ 250 mg/L Cef + 10 mg/L PPT
CIM
+ 250 mg/L Cef + 3 mg/L Glufosinate
CIM
+ 250 mg/L Cef + 5 mg/L PPT Shoot induction
medium
MS
+ 1 mg/L BAP
+ 100 mg/L Myo + 3% Suc + 250 mg/L Cef + 10 mg/L PPT + 0.2% Phytagel
MS
+ 0.01 mg/L 2,4-D + 1 mg/L Kinetin + 100 mg/L Myo + 2% Suc + 250 mg/L Cef
+ 3 mg/L Glufosinate + 0.22% Phytagel MS + 3% Suc + 250 mg/L Cef + 5mg/L PPT + 0.3% Gelrite
Plant regeneration medium
MS
+ 100 mg/L Myo + 3% Suc + 250 mg/L Cef + 10 mg/L PPT + 0.2% Phytagel
MS
+ 100 mg/L Myo + 2% Suc + 250 mg/L Cef + 3 mg/L Glufosinate + 0.22% Phytagel
MS + 3% Suc + 250 mg/L Cef + 10 mg/L PPT + 0.3% Gelrite
1
Dic, Dicamba (3,6-dichloro-o-anisic acid); BAP, 6-benzylaminopurine; 2,4-D, 2,4-dichlorophenoxyacetic acid; CH, Casein hydrolysate;
Myo, Myo-Inocitol; Suc, Sucrose; Glc, Glucose; AS, Acetosyringone; Cef, Cefotaxime; PPT, Phosphinothricin; bialaphos, L- phosphinothricinyl-L-alanyl-L-alanine
이나 재분화를 유도하기 전에 제초제나 항생제가 포함된 배지에서 유전자가 도입된 callus들만 선발하는 것으로, 암 조건에서 다양한 시간대에서 적절하게 선발된 callus 들만 이후의 shooting이나 재분화를 진행시켰을 때 높은 형질전환 효율을 보였다. 따라서 이러한 조건들이 최적 화되면 크리핑 벤트그라스의 형질전환은 비교적 잘 이루 어지는 것으로 여겨진다.
보고된 형질전환 효율을 비교해 보면, 현재까지 가장 높은 효율이 약 45%로 되어 있는데 (Luo et al. 2004), 이것 은 접종된 callus 당 PPT-resistant event를 기준으로 한 것 으로서 아마도 많은 PPT-resistant 식물체들이 실제로는 중복될 것으로 여겨진다. 보다 정확하게 측정된 것으로 는 callus 당 PPT-resistant shoot을 기준으로 하면 약 18-37.
2%, 최종적으로 온실에서 얻게 된 PPT-resistant shoot을 기준으로 하면 약 13.5-25.6%, 그리고 Southern blot 분석을
통하여 independent line들을 기준으로 하면 약 5.33-9.50%
로 보고되었다 (Kim et al. 2007). 실제 벤트그라스에서 shooting을 유도하면 multiple shoot이 유도되는 경향이 있 고, 같은 캘러스에서 유도된 대부분의 식물체는 Southern blot 분석에서 대체로 같은 결과를 보여주었다. 따라서 PPT-resistant event를 기준으로 한 형질전환 효율의 경우 한 캘러스에서 여러 개의 식물체를 선발하게 되면 실제 와는 다르게 높은 수치로 나올 수 있다. 그러므로 Southern blot 분석을 통한 independent line들을 기준으로 형질전환 효율을 측정하는 것이 적절하다고 판단되며, 이러한 기 준에 비추어 보면 크리핑 벤트그라스의 형질전환 효율은 현재의 최적화된 형질전환 과정을 거쳤을 때 약 5-10%
정도의 효율로 진행되는 것으로 판단할 수 있다. 결론적으
로 2000년도에 최초로 크리핑 벤트그라스의 Agrobacterium
매개 형질전환 성공과 두 개의 형질전환체가 보고된 이
후, 현재까지 형질전환 조건의 최적화가 진행되어 약 5-10% 정도의 형질전환 효율의 표준화된 벤트그라스 형 질전환 프로토콜이 완성된 것으로 판단된다 (Figure 1 및 Table 1 참조).
다양한 생명공학 벤트그라스 개발
벤트그라스가 주로 이용되는 골프장의 그린 같은 경우 는 많은 인원이 사용하기 때문에 잔디가 상처를 입고 생 육에 지장을 초래한다. 또한 우리나라의 여름처럼 고온 다습한 기후 조건이나 잔디 상태를 좋게 유지하기 위한 비료의 과다 사용으로 인해 벤트그라스는 병해충에 대한 저항성이 매우 낮다. 이에 잔디를 잘 관리하기 위해서 매 일 관수와 잦은 농약 방제를 하고 있는 실정이며, 이로 인해 많은 유지관리 비용이 발행하며 특히 최근 골프장 의 농약에 의한 환경오염이 사회적으로 문제되어 농약의 종류와 사용횟수도 제한되고 실정이다 (Wang et al.
2003a; Fu et al. 2005b; Wang and Ge 2006a). 따라서 이러한 약점을 극복하기 위하여 생명공학 잔디 개발이 필요하며, 2000년도에 Australia의 Melbourne에서 개최된 “Millennium Turfgrass Conference”에서 발표된 내용 중에 전통 육종 방 법에 의해 만들어지기 매우 힘든 잔디의 중요한 형질들 로 제초제 저항성 (herbicide resistance), 병해충 저항성 (disease and insect resistance), 내염성 및 내건성 (salt and drought tolerance), 효율적 영양분 사용 (nutrient use) 등이 발표되었다. 이러한 내용을 토대로 생명공학 잔디의 목 표 형질들을 크게 나누어 보면, 제 1세대로 여겨지는 제 초제 저항성 잔디, 제 2세대로 잔디의 유지관리 비용을 절감할 수 있는 병해충 저항성 및 내염성과 내건성 같은 스트레스 저항성 잔디, 제 3세대로 고부가가치 잔디 등으 로 구분할 수 있다. 따라서 이러한 형질을 도입시킨 생명 공학 잔디 개발을 위하여 많은 연구들이 진행되었다.
현재까지 보고된 형질전환 벤트그라스 잔디들을 Table 2 에 요약하였다. 여기에는 형질전환이나 효율 증진 등에 대한 보고들은 포함하지 않았다 (Zhong et al. 2003; Xiao and Ha 1997; Yu et al. 2000; Chai et al. 2004; Han et al.
2005; Fu et al. 2005a). 형질전환에 이용된 품종으로는 1950년대에 개발되어 현재까지도 널리 이용되는 Penncross 품종이 가장 많았고, 1990년대 이후 개발된 품종으로 Crenshaw 품종이나 Penn-A-4 품종 등이 사용되었다. 또한 도입하고자 하는 형질들의 경우, 제초제 저항성이 가장 많았으며, 다음으로 병 저항성과 가뭄 저항성 및 내염성, 그리고 최근에는 노화지연이나 anthocyanin이 축적된 보 라색 잔디 등이 있었다. 제초제 저항성 잔디가 많이 개발 된 이유는 초기에 형질전환 과정에서 selectable marker로 제초제 저항성 유전자 (bar)가 주로 이용되었기 때문에 형질전환 조건 확립과 동시에 얻을 수 있었기 때문이었다.
그리고 벤트그라스의 가장 큰 약점 중의 하나가 잡초와 의 경쟁에서 약하다는 것이며, 이로 인해 Annual bluegrass 등이 골프장에서 벤트그라스보다 우점이 가능하여 이러 한 잡초들을 방제하기 위해 오랜 시간 화학적 방법과 재 배적 방법에 관한 수많은 연구가 진행되었으나 모든 잡 초 종류에 대한 효과적인 방제 방법은 아직 없는 실정이 다. 따라서 이들의 효과적인 방제와 농약 사용 등을 줄이 기 위해 비선택성 제초제인 Roundup
®또는 BASTA
®등에 저항성을 가지는 제초제 저항성 생명공학 잔디를 우선적 으로 개발되었다 (Hartmann et al. 2004; Lee et al. 2006;
Asano et al. 1997; 1998; Chai et al. 2003; Luo et al. 2004;
Kim et al. 2007; Guo et al. 2009). 이 잔디들은 특정 제초제 만으로 다른 잡초를 동시에 효과적으로 방제가 가능하기 때문에 농약사용을 줄일 뿐 아니라 잔디 관리를 수월하 게 할 수 있다.
골프장에서는 벤트그라스는 높은 밀도로 재배하며 수 시로 짧은 예초를 하게 되는데, 이는 Rhizoctonia solani에 의한 갈색 마름병 (brown patch), Sclerotinia homeocarpa에 의한 동전 마름병 (dollar spot), Pythium spp.에 의한 Pythium blight병 등 광범위한 곰팡이 병원균에 대해 감수 성을 증가시킨다 (Belanger et al. 2004; Bonos et al. 2006).
따라서 골프장에서 이 병해를 막기 위해 많은 양의 살균 제를 살포하게 되는데, 이는 높은 유지관리 비용 문제뿐 만 아니라 환경적인 문제를 일으키게 된다. 이에 벤트그 라스의 병 저항성 (disease resistance)을 증가시키고 살균 제의 사용을 줄여 잔디의 유지관리 비용을 낮출 수 있는 품종의 개발이 진행되었다. 특히 골프장에서 크게 문제 되는 Rhizoctonia solani에 의한 갈색 마름병과 Sclerotinia
homoeocarpa에 의한 동전 마름병에 대한 저항성을 지니는 잔디를 개발하는 연구가 많이 진행되었다. 현재까지 보고된 것에 의하면, PR (pathogenesis-related) 단백질과 관 련된 유전자들, 예를 들어 rice의 PR-5 유사 유전자 (tlpD34)나 PR-5와 결합하는 kinase인 PR5K 유전자들을 이 용하여 동전 마름병에 저항성을 갖는 잔디가 개발되었다 (Guo et al. 2003; Fu et al. 2005b). 또한 미국자리공 (pokeweed) 식물에서 분리한 ribosome-inactivating protein 인 pokeweed antiviral protein (PAP)를 도입시켜 동전 마름 병에 저항성을 보이는 잔디가 개발되기도 하였다 (Dai et al. 2003). 하지만 이들 생명공학 잔디들에서는 갈색 마름 병에 뚜렷한 저항성을 발견할 수는 없었다 (Fu et al.
2005b). 다만 chitinase와 glucanase를 동시에 도입시킨 잔 디의 경우, 동전 마름병과 갈색 마름병 모두에 저항성을 보임이 관찰되었다 (Wang et al. 2003b). 본 연구진의 경 우, 고추에서 분리한 esterase 유전자인 PepEST1 (Kim et al.
2001)를 도입시킨 생명공학 벤트그라스에서 아주 우수한 갈색 마름병 저항성을 확인할 수 있었다 (unpublished).
가뭄에 매우 취약한 Creeping bentgrass의 단점을 보완
332 J Plant Biotechnol (2009) 36:327–335
Table 2 Developed biotech creeping bentgrass Transformation
method
Cultivar Marker
gene
Target gene Target trait References
Microprojectile bombardment
SouthshoreEmerald bar/gus
1bar Herbicide resistance Hartman et al. (1994) Crenshaw hph Arabidopsis PR5K Disease resistance Guo et al. (2003) Crenshaw hph PAP (pokeweed antiviral
protein) Disease resistance Dai et al. (2003) Province Penn-A-4 bar chintase+glucanse Disease resistance Wang et al. (2003b)
Electroporation
Cobra bar bar Herbicide resistance Lee et al. (1996)
Penncross bar bar Herbicide resistance Asano et al. (1997)
Penncross bar/gus bar Herbicide resistance Asano et al. (1998)
Agrobacterium-mediate d method
Regent Tiger bar/gfp bar Herbicide resistance Chai et al. (2003)
Penn-A-4 hyh/gus
or bar bar Herbicide resistance Luo et al. (2004) Crenshaw bar Rice tlpD34 Disease resistance Fu et al. (2005b) Penncross bar Cowpea VuNCED1 Drought/salt tolerance Aswath et al. (2005) Crenshaw bar Barley hva1 Drought tolerance Fu et al. (2007) Crenshaw
Penncross bar/gus bar Herbicide resistance Kim et al. (2007)
Crenshaw bar/gus Maize Lc+Pl Purple
-color Han et al. (2009)
Penncross hyh SAG12-ipt Delayed senescence Xu et al. (2009)
1
bar, bialaphos resistance gene; gus, β-glucuronidase; hph, hygromycin phosphotransferase; gfp, green fluorescent protein
하기 위해 가뭄 저항성 잔디도 개발되었다. 이를 위하여 식물의 스트레스 호르몬으로 알려진 ABA의 생합성에 관 여하는 것으로 알려진 9-cis-epoxycarotenoid dioxygenase 유 전자 (VuNCED1)를 cow pea에서 분리하여 벤트그라스로 도입시켜 분석한 결과, ABA 합성이 증대되어 가뭄 저항 성과 내염성이 증대되었음이 보고되었다 (Aswath et al.
2005). 또한 종자 성숙기간에 축적되고 다양한 stress 조건 에서 강하게 유도되면서 식물에 가뭄 저항성을 줄 수 있 는 것으로 알려진 LEA (late-embryogenesis-abundant) 단백 질도 이용되었는데, 보리 (Hordeum vulgare)의 LEA3인
hva1 유전자를 ABA inducible promoter를 이용하여 벤트그라스로 도입시킨 결과 가뭄 저항성이 크게 증대됨이 관 찰되었다 (Fu et al. 2007). 본 연구진도 ABA 신호전달에 중요한 ABF3 유전자 (Kang et al. 2002)를 벤트그라스로 도입시켜 가뭄 저항성이 뛰어난 생명공학 벤트그라스 잔 디를 개발할 수 있었다 (unpublished).
최근에는 벤트그라스 형질전환 시스템이 안정화되면 서 제초제 저항성, 병 저항성이나 스트레스 저항성 이외 에 고부가가치 형질을 도입시키는 연구도 진행되고 있 다. 예를 들어, 벤트그라스 잔디의 녹색기간을 연장시키 기 위하여 노화를 지연시키도록 하는 연구가 본 연구진 을 포함하여 진행되었다. 이때 식물 호르몬인 cytokinin 생합성에 관여하는 유전자인 IPT (isopentenyl transferase)
를 senescence-specific promoter인 SAG12에 붙여 이용하였 다 (Gan and Amasino 1995). 이 SAG12-IPT construct를 벤트 그라스로 도입시켜 분석한 결과, 노화지연뿐 아니라
SAG12-IPT를 도입시킨 한지형 잔디인 벤트그라스에서는고온에서도 잘 자라는 특징을 보여주었다 (Xu et al. 2009).
그리고 안토시아닌 생합성을 조절하는 전사인자 유전자 들인 Lc와 Pl을 maize로부터 분리하여 동시에 벤트그라 스로 도입시켜, 안토시아닌이 축적되어 보라색을 띄는 잔디가 개발되기도 하였다 (Han et al. 2009). 최근에는 벤 트그라스의 상업화 과정에서 크게 문제가 된 유전자 이 동 (gene flow) 문제를 제어하기 위한 웅성불임 잔디가 개 발되고 있는 데, 본 연구진의 경우 pollen-specific promoter 인 TA29 promoter에 RIP (ribosome-inactivating protein) 유전 자를 붙여 벤트그라스로 도입시켜 분석 중에 있는 데, 이 잔디는 sterile pollen을 만들게 되어 유전자 이동성 문제를 해결할 수 있는 대책을 제시할 수 있을 것으로 기대된다.
결론적으로 전통육종 방법으로는 개발하기 힘든 벤트
그라스 품종을 생명공학 기법을 이용한 방법으로 개발하
기 위한 연구가 활발히 진행되고 있으며, 현재까지 개발
된 생명공학 벤트그라스 잔디로 제초제 저항성 잔디, 가
뭄 저항성 잔디, 병 저항성 잔디, 그리고 노화지연이나
색을 지니는 고부가가치 잔디 등이 있다. 그리고 앞으로
도 웅성불임 잔디나 여러 형질을 동시에 지니는 복합형
질잔디들이 계속 개발되어, 지속적으로 늘어나는 잔디 품종에 대한 수요를 충족하게 될 것으로 여겨진다.
생명공학 벤트그라스의 상업화 추진 현황
벤트그라스 형질전환이 최초로 보고된 1993년 이후 다양 한 생명공학 벤트그라스들이 개발되어지고 있다 (Table 2 참조). 특히 제초제 저항성 생명공학 벤트그라스는 여러 가지 장점으로 인해 상업화하려는 시도가 지속적으로 전 개되고 있다. 현재 제초제 저항성 벤트그라스 잔디의 상업 화를 추진하는 회사는 미국에 있는 “The Scotts Company”
이다. 이 잔디는 Mansanto Company와 공동으로 개발된 것으로, Microprojectile Bombardment 방법으로 Roundup
®브랜드 제초제의 활성 성분인 glyphosate에 저항성을 지 니도록 bacterial EPSPS (5-enolpyruvyl-3- phosphoshikimate synthase) 유전자를 도입시킨 Roundup Ready 벤트그라스 잔디로서 최근에 미국 특허로 등록되었다 (Guo et al.
2009; US patent No. 7,569,747). 이 생명공학 잔디는 1990 년대 후반에 포장시험을 포함한 많은 연구가 진행되었 고, 2003년에 특허 출원을 하였으나 2009년 8월에서야 등 록이 되었다. 실제 생명공학 잔디 품종에 대한 특허는 최 근에서야 등록이 이루어졌으며, 현재까지 3건의 벤트그 라스 품종이 미국 특허로 등록된 것으로 파악되었다. 이 렇게 개발되어 탈규제 (deregulation)를 추진 중인 Roundup Ready Creeping Bentgrass 잔디는 Event “ASR-368”로 명명 되었으며, ATCC (American Type Culture Collection)에 accession No. PTA-4816으로 기탁되어 있다.
Event ASR-368 크리핑 벤트그라스에 대한 환경위해성 평가 등은 2000년부터 미국 Oregon, New Jersey, Ohio state 등 여러 곳에서 광범위하게 수행되었으며, 2003년 4월에 USDA 산하 APHIS (Animal and Plant Health Inspection Service)에 deregulation 심사를 요청하였다 (Nelson and Stone 2003). USDA/APHIS의 예비 환경위해성 평가 결과, 생명공학 크리핑 벤트그라스는 표현형에서는 일반 잔디 와 거의 동등하게 관찰되었으며, 생명공학 잔디 내에 EPSPS 유전자가 한 개가 도입되어 있고 멘델 법칙을 따 른다는 것, 그리고 일반 잔디와 비슷하게 병해충 감수성 또는 내성을 지니는 것으로 평가되었다 (USDA/APHIS 2004). 하지만 이 잔디들로부터의 유전자 이동성 (gene flow)이 최초 보고보다 훨씬 더 멀리 갈 수 있음이 관찰되 어, transgene escape 가능성 문제 제기 때문에 deregulation 이 이루어지지는 못하였다. 처음 보고에서는 미국에서 벤트그라스를 가장 많이 생산하는 지역인 Oregon 주의 Willamette Valley에서 수행된 intra-specific 및 inter-specific gene flow 실험 결과, 교잡율은 약 0.2%로 나타났고 유전
자 이동이 약 1.6 km 이내에서 관찰되었다 (Wipff and Fricker 2001). 하지만 미국 EPA 연구실에서 실험을 수행 한 결과, 대부분의 유전자 이동이 바람이 부는 쪽으로 약 2 km 내에서 일어났으나, 최대 유전자 이동이 21 km 지 역에서도 관측되는 것이 확인되었다 (Watrud et al. 2004).
이는 다른 잔디 종과 비교했을 때 (Bae et al. 2008), 벤트 그라스 잔디가 매우 작은 화분 (pollen)과 종자 (seed)를 지 니는 것에 기인하는 것으로 여겨진다. 벤트그라스 화분 은 지름이 25.4 μm, 평균 무게가 52 ng이며 종자는 그램 당 약 13,500개의 종자가 포함될 정도로 작기에 유전자 이동성이 큰 것으로 파악된다 (Mallory-Smith and Zapiola 2008). 뿐만 아니라 nonagronomic habitat에서도 Roundup Ready Creeping Bentgrass가 발견됨도 보고되었다 (Reichman et al. 2006; Zapiola et al. 2008). 따라서 이러한 환경 문제 가 제기됨에 따라 Roundup Ready Creeping Bentgrass에 대 한 허가는 철저한 환경영향평가를 수행한 이후로 연기되 었다.
The Scotts Company는 APHIS에 Roundup Ready Creeping Bentgrass의 deregulation 심사를 요청한 이후, 회사 홈 페이지 (www.scottsproseed.com) 내의 future product로 올려 놓고 이 잔디를 홍보하였으나, 국제기술평가센터 (ICTA, International Center for Technology Assessment)가 USDA를 대상으로 이 잔디가 다른 식물 종의 생존을 위협할 수 있 기에 deregulation 신청을 기각해야 한다는 민원을 제출하 였고, 2007년 2월 미국 법원이 이를 받아들임으로써 현재 는 Roundup Ready Creeping Bentgrass를 홈페이지에서 삭 제하고 후속 연구를 진행 중인 것으로 알려졌다. 특히 gene flow 문제를 근본적으로 일으키지 않는 웅성불임의 벤트그라스를 개발하는 연구도 수행하고 있는 것으로 알 려졌다. 결론적으로 제초제 저항성 생명공학 벤트그라 스는 잔디 재배 및 관리 측면에서 매우 큰 장점을 지닌 매우 좋은 생명공학 제품으로 평가되지만, 상업화를 위 한 deregulation 과정은 순탄치 않을 것으로 전망된다. 이 는 생명공학 벤트그라스가 다른 상업화된 생명공학 작물 들 (콩, 옥수수, 면화 등)과 달리 처음으로 농지가 아닌 교외 환경에 심는 다년생 식물이라는 점과, 바람에 의 한 가루받이 (wind-pollination)가 일어나고 이종교배 (out- crossing)가 매우 잘 일어나는 식물이라는 점에서 기존 생 명공학 농작물과는 다르게 다루어지고 있다. 또한 이미 상업화된 생명공학 작물들은 카놀라를 제외하면 야생종 천적이 없고 경작지를 벋어나면 살아남기 어렵지만 잔디 는 최소 12가지 이상의 야생종과 교차수정을 할 수 있고 야생에서도 생존이 가능한 점도 문제로 지적되고 있다.
따라서 향후 이러한 문제점들을 극복할 수 있는 새로운
생명공학 연구가 진행될 것으로 추정된다.
334 J Plant Biotechnol (2009) 36:327–335
결론과 전망
최근 생활수준 향상에 따라 잔디 요구량과 다양한 잔 디 품종에 대한 수요가 지속적으로 늘어나고 있다. 하지 만 전통육종 방법을 통한 품종 개량이나 개발은 한계가 있고, 어렵고, 시간과 비용이 많이 드는 단점이 있어 최 근에는 생명공학 기법을 이용한 잔디 품종 개발 연구가 많이 진행되고 있다. 실제로 골프장에 전적으로 쓰이는 여러 크리핑 벤트그라스 품종에 대한 형질전환 시스템이 개발되었고, 이를 이용하여 제초제 저항성 잔디, 가뭄 또 는 병 저항성 잔디, 그리고 노화지연 잔디나 색깔 잔디 등 기존의 전통육종 방법으로는 얻을 수 없었던 새로운 형질을 지닌 생명공학 잔디들이 개발되었다. 향후에도 새로운 유전자 연구 진행과 더불어 더 많은 생명공학 잔 디가 개발될 것으로 여겨지며, 미래에는 다양한 형질을 복합적으로 지니는 생명공학 잔디도 개발될 것으로 예측 된다. 하지만 개발된 생명공학 벤트그라스의 상업화를 위한 탈규제 과정을 위하여 앞으로 더 많은 연구가 필요 할 것으로 판단된다.
새로운 Millenium이 시작된 2000년도에 뉴욕타임즈가 선정한 2010년의 첨단상품들에 “깎을 필요 없는 잔디”가 포함되어 있었다 (www.nytimes.com/library/magazine/home/
20000611mag-lawn.html). 이것은 미국 Salk Institute의 Joanne Chory 박사가 발견한 BAS-1이라는 유전자를 담배로 도입 시켰을 때 1.5 m이상 자라던 담배가 약 25 cm로 되는 것 을 관찰하였고, 이 유전자를 벤트그라스 잔디로 도입시 켜 키가 작아 깎을 필요 없는 잔디를 개발할 수 있다는 기대감으로 선정된 것으로 추정된다. 이러한 예에서 보 듯이 미래 사회에 필요하고 사람들의 기대에 부응하는 생명공학 잔디 개발은 앞으로도 중요한 연구가 될 것으 로 판단된다.
사 사
본 논문과 관련된 연구는 농촌진흥청 바이오그린21사 업 (과제번호: 20080401034014)의 지원에 의해 이루어졌 으며 이에 감사드립니다.
인용문헌
Asano Y, Ito Y, Fukami M, Morifuji M, Fujiie A (1997) Pro- duction of herbicide resistant transgenic creeping bent plants.
Intl Turfgrass Soc Res J 8:261-267
Asano Y, Ito Y, Fukami M, Sugiura K, Fujiie A (1998) Production of herbicide-resistant transgenic creeping bentgrass plants obtained by electroporation using an altered buffer. Plant Cell
Rep 17:963-967
Aswath CR, Kim SH, Mo SY, Kim DW (2005) Transgenic plants of creeping bent grass harboring the stress inducible gene, 9-cis-epoxycarotenoid dioxygenase, are highly tolerant to drought and NaCl stress. Plant Growth Regul 47:129-139 Bae TW, Vanjildorj E, Nishiguchi S, Song SY, Yang SS, Song IJ,
Chandrasekhar T, Kang TW, Kim JI, Koh YJ, Park SY, Lee J, Lee YE, Ryu KH, Riu KZ, Song PS, Lee HY (2008) Environmental risk assessment of genetically engineered herbicide-tolerant Zoysia japonica. J Environ Qual 37:207-218 Belanger FC, Bonos S, Meyer WA (2004). Dollar spot resistant
hybrids between creeping bentgrass and colonial bentgrass.
Crop Sci 44:581-586
Bonos SA, Plumley KA, Meyer WA (2002) Ploidy determination in Agrostis using flow cytometry and morphological traits.
Crop Sci 42:192-196
Bonos SA, Clarke BB, Meyer WA (2006) Breeding for disease resistance in the major cool-season turfgrasses. Annu Rev Phytopathol 44:213-234
Chai B, Sticklen MB (1998) Applications of biotechnology in turfgrass genetic improvement. Crop Sci 38:1320-1338 Chai ML, Wang BL, Kim JY, Lee JM, Kim DH (2003)
Agrobacterium-mediated transformation of herbicide
resistance in creeping bentgrass and colonial bentgrass. J Zhejiang Univ Sci 4:346-351Chai ML, Senthil KK, Kim DH (2004) Transgenic plants of colonial bentgrass from embryogenic callus via Agrobacterium-mediated transformation. Plant Cell Tiss Org Cult 77:165-171 Dai S, Zheng P, Marmey P, Zhang S, Tian W, Chen S, Beachy RN,
Fauquet C (2001) Comparative analysis of transgenic rice plants obtained by Agrobacterium-mediated transformation and particle bombardment. Mol Breed 7:25-33
Dai WD, Bonos S, Guo Z, Meyer WA, Dai ZY, Belanger FC (2003). Expression of pokeweed antiviral proteins in creeping bentgrass. Plant Cell Rep 21:497-502
Engelke MC, Lehman VG, Kneebone WR, Colbaugh PF, Reinert JA, Knoop WE (1995) Registration of ‘Crenshaw’ Creeping Bentgrass. Crop Sci 35:590
Fu D, Xiao Y, Muthukrishnan S, Liang GH (2005a) In vivo performance of a dual genetic marker, manA-gfp, in transgenic bentgrass. Genome 48:722-730
Fu D, Tisserat NA, Xiao Y, Settle D, Muthukrishnan S, Liang GH (2005b) Overexpression of rice TLPD34 enhances dollar-spot resistance in transgenic bentgrass. Plant Sci 168:671-680 Fu D, Huang B, Xiao Y, Muthukrishnan S, Liang GH (2007)
Overexpression of barley hva1 gene in creeping bentgrass for improving drought resistance. Plant Cell Rep 26:467-477 Gan S, Amasino RM (1995) Inhibition of leaf senescence by
autoregulated production of cytokinin. Science 270:1986-1988 Guo Z, Bonos S, Meyer WA, Day PR, Belanger FC (2003)
Transgenic creeping bentgrass with delayed dollar spot symptoms. Mol Breed 11:95-101
Guo SX, Harriman RW, Lee L, Nelson EK (2009) Bentgrass event ASR-368 and compositions and methods for detection thereof. US Patent No. 7,569,747 B2.
Han N, Chen D, Bian HW, Deng MJ, Zhu MY (2005) Production of transgenic creeping bentgrass Agrostis stolonifera var.
palustris plants by Agrobacterium tumefaciens-mediated transformation using hygromycin selection. Plant Cell Tiss Org Cult 81:131-138
Han YJ, Kim YM, Lee JY, Kim SJ, Cho KC, Chandrasekhar T, Song PS, Woo YM, Kim JI (2009) Production of purple-colored creeping bentgrass using maize transcription factor genes Pl and Lc through Agrobacterium-mediated transformation.
Plant Cell Rep 28:397-406
Hartman CL, Lee L, Day RR, Tumer NE (1994) Herbicide resistant turfgrass (Agrostis palustris Huds.) by biolistic transformation . Bio/Technology 12:919-923
Kang JY, Choi HI, Im MY, Kim SY (2002) Arabidopsis basic leucine zipper proteins that mediate stress-responsive abscisic acid signaling. Plant Cell 14:343-357
Kim SJ, Lee JY, Kim YM, Yang SS, Hwang OJ, Hong NJ, Kim KM, Lee HY, Song PS, Kim JI (2007) Agrobacterium–mediated high efficiency transformation of creeping bentgrass with herbicide resistance. J Plant Biol 50:577-585
Kim YS, Lee HH, Ko MK, Song CE, Bae CY, Lee YH, Oh BJ (2001) Inhibition of fungal appressorium formation by pepper (Capsicum annuum) esterase. Mol Plant Microbe Interact 14:80-85
Lee L (1996) Turfgrass biotechnology. Plant Sci 115:1-8 Lee L, Laramore CL, Day PR, Tumor NE (1996) Transformation
and regeneration of creeping bentgrass (Agrostis palustris Huds.) protoplasts. Crop Sci 36:401-406
Luo H, Hu Q, Nelson K, Longo C, Kausch AP, Chandlee JM, Wipff JK, Fricker CR (2004) Agrobacterium tumefaciens-mediated creeping bentgrass (Agrostis stolonifera L.) transformation using phosphinothricin selection results in a high frequency of single-copy transgene integration. Plant Cell Rep 22:645-652 Mallory-Smith C, Zapiola M (2008) Gene flow from glyphosate-
resistant crops. Pest Manag Sci 64:428-440
Nelson E, Stone T (2003) Petition for determination of non-regulated status: Roundup Ready Creeping Bent grass Event ASR368.
Petition #01-TR-054U [www.aphis.usda.gov/brs/not_reg.html]
Reichman JR, Watrud LS, Lee EH, Burdick CA, Bollman MA, Storm MJ, King GA, Mallory-Smith C (2006) Establishment of transgenic herbicide-resistant creeping bentgrass (Agrostis
stolonifera L.) in nonagronomic habitats. Mol Ecol 15:4243-
4255USDA/APHIS (2004) APHIS Preliminary Risk Assessment on the Petition for a Determination of Nonregulated Status for Creeping Bentgrass (Agrostis stolonifera) Genetically Engineered (Event ASR368) for Tolerance to the Herbicide Glyphosate submitted by Monsanto Company and the Scotts Company.
[http://www.aphis.usda.gov/ brs/not_reg.html]
Wang Y, Browning M, Ruemmele BA, Chandlee JM, Kausch AP
(2003a) Glufosinate reduces fungal diseases in transgenic glufosinate-resistant bentgrasses (Agrostis spp.). Weed Sci 51:130-137
Wang Y, Kausch AP, Chandlee JM, Luo H, Ruemmele BA, Browning M, Jackson N, Goldsmith MR (2003b) Co-transfer and expression of chitinase, glucanase, and bar genes in creeping bentgrass for conferring fungal disease resistance.
Plant Sci 165:497-506
Wang Z, Hopkins A, Mian R (2001) Forage and turf grass biotechnology. Crit Rev Plant Sci 20:573-619
Wang ZY, Ge Y (2006a) Recent advances in genetic trans- formation of forage and turf grasses. In Vitro Cell Dev Biol Plant 42:1-18
Wang ZY, Ge Y (2006b) Rapid and efficient production of transgenic bermudagrass and creeping bentgrass bypassing the callus formation phase. Funct Plant Biol 32:769-776 Warnke S (2003) Creeping bentgrass (Agrostis stolonifera L.), In:
Casler MD, Duncan RR, (eds), Turfgrass Biology, Genetics, and Breeding. John Wiley & Sons, Inc., NJ, USA, pp 175-185 Watrud LS, Lee EH, Fairbrother A, Burdick C, Reichman JR,
Bollman M, Storm M, King G, Van de Water PK (2004) Evidence for landscape-level, pollen-mediated gene flow from genetically modified creeping bentgrass with CP4 EPSPS as a marker. Proc Natl Acad Sci USA 101:14533-14538
Wipff JK, Fricker C (2001) Gene flow from transgenic creeping bentgrass (Agrostis stolonifera L.) in the williamette valley, Oregon. Intl Truf Soc Res J 9:224-242
Xiao L, Ha SB (1997) Efficient selection and regeneration of creeping bentgrass transformants following particle bom- bardment. Plant Cell Rep 16:874-878
Xu Y, Tian J, Gianfagna T, Huang B (2009) Effects of SAG12-ipt expression on cytokinin production, growth and senescence of creeping bentgrass (Agrostis stolonifera L.) under heat stress. Plant Growth Regul 57:281-291
Yu TT, Skinner DZ, Liang GH, Trick HN, Huang B, Muthukrishnan S (2000) Agrobacterium-mediated transformation of creeping bentgrass using GFP as a reporter gene. Hereditas 133:229-233 Zapiola ML, Campbell CK, Butler MD, Mallory-Smith CA (2008)
Escape and estabilishment of gransgenic glyphosate-resistant creeping bentgrass Agrostis stolonifera in Oregon, USA: a 4-year study. J Appl Ecol 45:486-494
Zhong H, Boyland MG, Srinivasan C, Sticklen MB (1993) Transgenic plants of turfgrass (Agrostis palustris Huds.) from microprojectile bombardment of embryogenic callus. Plant Cell Rep 13:1-6
Zilinskas BA, Wang X (2004) Genetic transformation of turfgrass, In: Liang GH, Skinner DZ, (eds), Genetically Modified Crops: Their Development, Uses, and Risks. Food Product Press, an imprint of the HaworthPress, Inc., New York, pp 309-350