균일한 전단응력에 의한 혈관내피세포의 운동성 변화
신현정
†
· 송석현†
EFFECTS OF UNIFORM SHEAR STRESS ON THE MIGRATION OF VASCULAR ENDOTHELIAL CELL
Jennifer H. Shin and Sukhyun Song
Key Words :
Vascular Endothelial Cell(혈관내피세포), Cell Motility(세포 운동성), Fluid Shear Stress(유동전단응력), Substrate Stiffness(바닥강성)Abstract
The migration and proliferation of vascular endothelial cells (VEC), which play an important role in vascular remodeling, are known to be regulated by hemodynamic forces in the blood vessels. When shear stresses of 2, 6, 15 dynes/cm
2are applied on mouse micro-VEC in vitro, cells surprisingly migrate against the flow direction at all conditions. While higher flow rate imposes more resistance against the cells, reducing their migration speed, the horizontal component of the velocity parallel to the flow increases with the flow rate, indicating the higher alignment of cells in the direction parallel to the flow at a higher shear stress. In addition, cells exhibit substrate stiffness and calcium dependent migration behavior, which can be explained by polarized remodeling in the mechanosensitive pathway under shear stress.
1. INTRODUCTION
The vascular endothelium forms a continuous monolayer lining of the luminal surface of the cardiovascular system, providing a structural and communicational interface between the circulating blood and underlying tissue. Vascular endothelial cells (VECs) are continuously exposed to hemodynamic forces such as the tensile stress of blood vessel dilatation and hydraulic pressure and shear stress from blood flow as well as biochemical stimuli from the neighboring cells and blood cells (1).
Atherosclerosis, one of the most common cardiovascular diseases, is known to develop in specific
regions of arterial trees with abnormal blood flow patterns and lower shear stress than normal (1, 2). In addition, recent research shows that shear stress due to abnormal flow induces significant changes in the structure and function of endothelial cells, which include changes in gene and protein expression, cytoskeleton arrangement, proliferation rate, cell migration, and apoptosis (3). Therefore, proper shear stress conditions are essential for VEC to keep their innate characteristics expressed in morphological and physiological behavior;
also, shear stress can be a main key of cardiovascular disease.
In this research, we first develop a parallel plate flow chamber which allows the application of a desired shear stress on the cultured VEC. It is designed based on three- dimensional computational fluid dynamics (3D-CFD) analysis. To apply the desired shear stress on the VEC monolayer, the flow chamber system is designed to minimize undesired pulsation due to peristaltic pumping and to maintain essential cell culture conditions.
†
Mechanical Engineering Bldg. 5205,
Korea Advanced Institute of Science and Technology (KAIST), 373-1 Guseong-dong, Yuseong-Gu, Daejeon, 305-701, Republic of Korea
E-mail : [email protected]
TEL : (042)350-3232 FAX : (042)350-5230
대한기계학회 2008년도 추계학술대회 논문집Moreover, dy by the flow r
The dynam proliferation remodeling healing. In t cell motility have been stu Moreover, s migration be remodeling i stress, have i
2. M
2.1 Comp To analy dynamics, (Fluent Inc.
(Fluent Inc generate the
2.2 Shear Fig. 1 sho setup for the A parallel-p desired she monolayer.
plates and 0.5mm-heig which VEC chamber.
The flow flow circuit (Cole Parme damper, and pump and d media. For t temperature controlled w 95% air) blo Wall shea determined manipulate
ynamic patter rate controller mics response
play an during angi this research, in the mouse udied under v substrate stiff ehavior, which in the mechan investigated.
MATERIAL
putational Flui ze and solve the numerica ., USA) is use c., USA), we e mesh of the
r Stress System ows a schema e shear-induci plate flow ch
ar stress on The flow ch a sandwichin ght flow chann C is plated i
w chamber is t which is co er, Masterflex d reservoir wi damper genera the real-time o
is maintained with a humidi own into the re
ar stress is p by rotation pump speed,
rns of shear str .
es of VEC of important ro ogenesis as the shear stre e micro-vesse various flow c
fness and ca h can be expla nosensitive pat
LS AND ME
id Dynamics A problems rela al method sof
ed. In addition e can create flow chamber
m
atic diagram o ing device for hamber is us the cultured amber consis ng silicone g nel. Two mic is mounted i
connected to omposed of a x LJ-07523-60 ith culture me ate a stable pe
observation b d by water bat
fied gas mixt eservoir.
proportional al speed of a PID (Prop
ress are gener
f cell motility ole in vasc well as wo ess effects on el endothelial
onditions in v alcium depen ained by polar thway under s
ETHODS
Analysis ated to fluid ftware Fluent n, using GAM
a geometry r model.
of the experim r endothelial c sed to apply d endothelial sts of two ac gasket to mak croscope slide
n the bottom
o a re-circula a peristaltic p 0, USA), pulsa
edia. A perist erfusion of gro
y microscope th and pH lev ture (5% CO
2to the flow f the pump.
portional-Integ rated
and cular ound n the cell vitro.
ndent rized shear
flow t 6.0 MBIT
and
ment cells.
the cell rylic ke a es on m of
ating pump ation taltic owth e, the vel is
2
and rate To gral-
De inc 327 and flow per boa
2.3 F (AT end Me dis sod bov ant pen mic slid µg/
inc cal ion
2.4 D tim (ZE ima Ax pos Tra
Fig
rivative) cont cludes a liqui 704-02, USA) d a remote co w rate. For ristaltic pump ard (Sensoray,
3 Cell Cultu For the shear TCC CRL-22 dothelium) is edium (DMEM
solved in dist dium bicarbon vine serum tibiotics (Gi nicillin and s crobial conta des are sterili /ml fibronecti cubation at ro
cium chelatin n in growth me
4 Time-Lap Dynamic resp me-lapse imag EISS Axiove ages were re xiocam HSm,
sition of indi acking Module
g. 1 Schemati
trol scheme is id flow rate ) to measure ontrolled peris the interfa and compute , Model 626, U
ure Condition r stress expe 99, Mus musc
cultured in D M, Gibco 12 tilled and aut nate. It is sup (Gibco 261 ibco 15240- streptomycin aminations. B
ized by autoc in (Sigma F11 oom tempera ng agent, is ad
edia.
se Microscopy ponses of the ge using ZE ert-200M, G ecorded with
Germany) a ividual cells w
e in ImageJ so
ic diagram of
s used. Shear sensor (Cole- the flow rate staltic pump t ce between er, a multifunc USA) is used.
eriments, bEn culus brain ce Dulbecco's M 2800-017, US toclaved wate pplemented w 40-079, USA -096, USA)
to prevent b Before cell se clave and co 141-1MG, US ature. 1.81mM dded to deplet
y
e cells are m EISS inverted Germany). Ph a CCD cam at every 30 m was measured oftware (NIH,
in vitro shear
stress system -Parmer, CZ- e in real time, to manipulate flow meter, ction PCI I/O .
nd.3 cell line erebral cortex odified Eagle SA) which is r with 1.5g/L with 10% fetal A) and 1%
) containing bacterial and eeding, glass oated with 10 A) for 1 hour M of EGTA, te the calcium
monitored by d microscope hase contrast mera (ZEISS minutes. The d by Manual , USA).
stress device.
m - , e , O
e x e s L l
% g d s 0 r , m
y
e
t
S
e
l
3. RESULTS
3.1 Optimization of Parallel Plate Flow Chamber The most important part in this system is the parallel plate flow chamber. To find the optimal design for our cell monolayer plated flow chamber, four possible features, based on inlet and outlet configurations, are considered: whether the inlet and outlet are situated horizontally or vertically with respect to the flow chamber, and whether the inlet and outlet feature a diffuser and converging duct. By three-dimensional computational fluid dynamics analysis, Fluent 6.0, four types of models are evaluated.
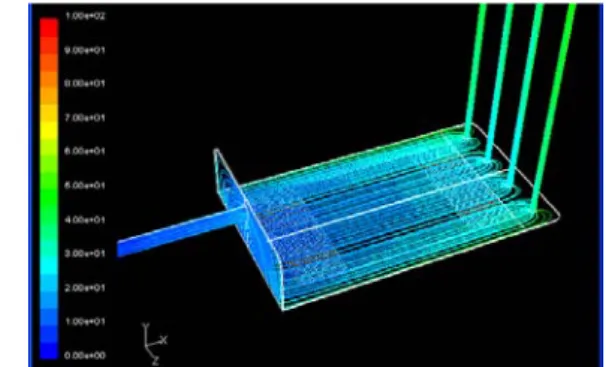
Simulation of shear stress distribution in a moderate flow rate (Q=60ml/min) condition are performed for validation of models. According to symmetry, half of the flow chamber is taken as the computational fluid dynamics simulation. When the flow rate is increased, shear stress on the central region is much higher than on the side region in the horizontal inlet model compared to the vertical inlet model. This non-uniform shear stress field can be explained by observing the path line in the flow chamber. Path lines are colored from blue to red as time passes. In the horizontal inlet model, when the flow rate is increased, the water inlet behaves like a water jet and the water is not well dispersed in contrast to dispersion in the vertical inlet model. Moreover, the presence of a converging duct is helpful for a more uniform path line near the outlet. Therefore, we decided on the model which features a vertical inlet and outlet with converging ducts to ensure a uniform shear stress field inside the flow chamber. In addition, the flow chamber is changed to vertical multiple outlets to prevent vortex flow in converging ducts and is bent into an L-shape to extend the effective length of the chamber and minimize the inlet transient region effect. After detail modifications, the optimized flow chamber design is developed as shown in Fig. 2.
In a computational fluid dynamic simulation, wall shear stress is fully proportional to flow rate as the equation of τ (dyne/cm
2) = 0.0520 × Q (ml/min). The coefficient in this equation depends on the geometry of the flow chamber as shown in Eq. (1), which is derived by the parallel plate flow model (4). The uniform shear stress area of this system is substantially larger than
other systems (5-8), 75×50mm
2at minimum. Thus, we can guarantee a large and uniform shear stress region for conventional molecular biology work.
2
6
μ
τ μ
=
= =
w
y h
du Q
dy bh Eq. (1)
( τ : average wall shear stress, μ : viscosity,
wu : velocity in parallel plate function of height, Q : volume flow rate, b : width of chamber, h : height of chamber)
3.2 Shear stress effects on cell motility
Using the shear stress system, the shear stress effects on VEC migration have been studied as a function of time under various flow conditions. The migration velocity in parallel (V
X) and perpendicular (V
Y) to the direction of flow are calculated from the time-lapse images. Surprisingly, most cells tend to migrate against the flow direction when shear stress is applied for 16 hours. This directional migration of cells becomes remarkable as the magnitude of shear stress increases.
Moreover, while the average velocity in the direction parallel to the flow increases with the flow, the average speed decreases as the applied shear stress increases (Fig.
3). It means that higher flow rate applied more resistance against cell migration speed, but the direction of cell motility is more aligned to the flow direction. Trajectory of cells shows that the cell migration is strongly oriented parallel to the flow direction as the applied shear stress increases. Cells tend to migrate in random direction initially and orient themselves in the direction against the flow as time passes.
Fig. 2 Path line result of the final model simulation of
flow chamber
Fig. 3 Migr
3.3 Calciu To regulat and rear of calcium che speed both w However, in preferential d condition of condition, ce by shear str movement, d condition 3.4 Substr
Most cell against the fl for 16 hours.
remarkable a shown in F substrate bec more aligned tendency for direction as t
In this stud shear stress o
Fig. 4 Migr
ration speed(l the direction um Depletion te the activatio f cell, calcium elating agent.
with calcium a n the absen direction of ce f 15dyne/cm
2.
ells lose the ress. Cell po depends on c
rate Stiffness ls on soft su low direction . This directio as the stiffne Fig. 4, migra come soft, and d to the flow r cells to mig the substrate s
4. DIS
dy, we have e on the motility
ration speed(l
eft) and the av parallel to the Effects on Ce on difference m ion is dep . Cells migr and without c ce of calciu ell migration u . Under calci
directional m olarity, but n calcium ion u
Effects on Ce ubstrate also
under 15 dyn onal migration ess of substra ation speed d the direction direction. Th grate anti-par stiffness decre
SCUSSION
examined the e y of vascular e
eft) and the av
verage velocit e flow(right) ell Motility
between the f pleted by EG rate almost s calcium condi um, there is under shear s um ion deple movement indu not the speed under shear s
ell Motility tend to mig ne/cm
2shear s
n of cells beco ate decreases
increases as n of cell motili
here is a grow rallel to the eases.
N
effects of unif endothelial ce
verage velocit ty in
front GTA, same ition.
s no tress etion uced d of
tress
grate tress omes . As
the ity is wing flow
form lls.
ty in
F regu and the bioc Thi effe sign forc foca spec rem shea cyto CD Mic the form inhi exp Mor acti pers also cros acti may sign In exp bec deg calc acti deg exp dep The pola
From the expe ulated by mec d substrate stif transformat chemical resp s can be div ects on cell m naling, increa ce, and active
al adhesion d cific directio modeling in t ar stress (9). B oskeleton such
C42 and R crotubule elon cell activates mation (10).
ibit Rho acti ression of Ra reover, posi vation and in sistent directio o evidence th sstalk with R vates Rac, an y contribute t naling (13).
n the experi ected that ce ause calcium gradation. Iss
cium ion can n-myosin con grade focal ad eriment, cell ends on calc erefore, calciu
arity in signali
substrat erimental resu chanical factor ffness. Theref tion of me onse, should b vided into fou membrane, po ase of cell ad
e contraction disassembly. T on can be the mechano Balance of sig h as Rho fami Rac can pro ngation due to Rac expressio
Simultaneou ivity locally, ac and Rho d itive feedba ntegrin activat onal migration hat focal adh Rho GTPase- nd inhibits R to polarized
iment of ca ells may not m ion is re hiki et al.
enhanced the ntraction and dhesions loca polarity but n cium ion und um ion has a
ing pathway r
te condition ults, cell migr rs such as flui fore, mechano echanical for
be studied mo ur steps: pur olarized mech dhesion to re
and protease The cell mig
explained b -sensitive pa gnaling pathw ily small GTP ovide good o shear stress a
on to promote usly, activate and results during cell m ck loop b tion in lamell n (12). In add hesion kinase -mediated sig Rho simultane activation of
lcium ion d t migrate as elated to foc found that s function of p d may activat ally (14). How
not the speed der shear stre another funct related to cell
ration can be id shear stress otransduction, rces into a ore intensively e mechanical hano-chemical esist external e activity for gration in the by polarized athway under way related to Pase of RhoA, explanations.
at the front of e lamellipodia ed Rac may in polarized migration (11).
etween Rac lipodia makes dition, there is e (FAK) can gnaling. FAK eously, which f cell motility
depletion, we time passes cal adhesion shear-induced p160ROCK in te calpain to wever, in our of movement ess condition.
tion to make motility.
e s , a y.
l l l r e d r o , . f a y d . c s s n K h y
e
s
n
d
n
o
r
t
.
e
5. CONCLUSION
By the experimental results as shown above, the shear stress system is successfully developed to maintain physiological conditions such as pH, temperature and shear stress and to apply steady and uniform shear stress to established cell monolayer. Moreover we observed that mechanical stimulus mediated by flow can affect the migration behaviors of vascular endothelial cell. Higher flow rate applied more resistance against cell migration speed, but the direction of cell motility is more aligned to the flow direction. In addition, cell polarity, but not the speed of movement, depends on calcium ion under shear stress condition. Moreover, cells tend to migrate anti- parallel to the flow direction as the substrate stiffness decreases. These unusual phenomena can be explained by polarized remodeling in the mechano-sensitive pathway under shear stress. The exact mechanisms for these interesting behaviors are still under investigation with the signaling pathway related to cytoskeleton.
REFERENCE