Taxonomic studies of tribe Epilobieae Endl. (Onagraceae) in Korea based on morphology and seed microstructure
Sangryong Lee
1, Kyeong-In Heo, Sangtae Lee, Manhee Yoo
2, Yongseong Kim, Joon Seon Lee and Seung-Chul Kim*
Department of Biological Sciences, Sungkyunkwan University, Suwon 440-746, Korea
1Botanical Gardens, Tohoku University, Aoba, Sendai 980-0682, Japan
2Korea Environmental Research Inc, Anyang 431-060, Korea
(Received 14 June 2013; Revised 6 August 2013; Accepted 3 September 2013)
외부형태와 종자의 미세구조에 의한 한국산 바늘꽃족(바늘꽃과)의 분류학적 연구
이상룡1·허경인·이상태·유만희2·김용성·이준선·김승철*
성균관대학교 생명과학과, 1일본 토호쿠대학교 식물원, 2한국환경기술개발(주) 부설생태연구소
ABSTRACT:
In this paper, we conducted the taxonomic study of the tribe Epilobieae Endl. and concluded that a total of nine taxa, including one Chamerion (Raf.) Raf. ex Holub and eight Epilobium L., exist in Korea.
Although C. angustifolium subsp. angustifolium has been placed traditionally either in Epilobium or Chamaen- erion, it can be clearly distinguished from the species of Epilobium by having alternate leaves, slightly zygo- morphic flowers, non-clefted petals, and equal length of 8 stamens, supporting the recognition of genus Chamerion. All but one species of Epilobium, E. platystigmatosum, was investigated for the surface of seeds using scanning electron microscope (SEM). The seed sculpture of Korean Epilobium can be classified into three types, i.e., papillose, reticulate, and ridged. E. ciliatum subsp. ciliatum is the only species Epilobium, which has the ridged seed sculpture. E. amurense subsp. cephalostigma can be distinguished from conspecific E. amurense subsp. amurense based on leaf shape, trichome shape and distribution, size and habit. Both E. fastigiatoramo- sum and E. palustre have entire leaf margins, but they can be distinguished based on leaf shape, stigma, and seed sculpture; the former has elongated elliptic leaves, capitate stigma, ridged seed sculpture, whereas the latter one has elongated lanceolate leaves, club-shaped stigma, and reticulate seed sculpture. Finally, we report the first record of E. platystigmatosum in Korea, and further comparative study including conspecific populations from Japan and China can clarify the occurrence of this taxon in Korea.
Keywords:
Epilobium, Chamerion, tribe Epilobieae, morphological characters, seed sculpture
적 요: 한국산 바늘꽃족(Epilobieae Endl.)에 대한 외부형태학적 형질에 근거한 분류학적인 연구를 수행하였다.
그 결과 한국에는 분홍바늘꽃속(Chamerion (Raf.) Raf. ex Holub) 1분류군과 바늘꽃속(Epilobium L.)분류군이 존 재하는 것으로 정리되었다. 분홍바늘꽃(C. angustifolium subsp. angustifolium)은 이전까지 바늘꽃속이나 또는 분 홍바늘꽃속에 속했으나, 잎이 호생하고, 꽃은 다소 좌우상칭이며 꽃잎의 끝이 갈라지지 않고, 수술 8개의 길이 가 거의 비슷한 특징으로 바늘꽃속 식물들과 잘 구분되었다. 가는민바늘꽃(E. platystigmatosum)을 제외한 바늘
*Author for correspondence: [email protected] /[email protected] http://www.pltaxa.or.kr
Copyright © 2013 the Korean Society of Plant Taxonomists
꽃속 종들의 종자 표면을 전자현미경으로 관찰한 결과 종자 표면은 유두상 돌기(papillose), 그물형 돌기 (reticulate), 이랑형 배열의 유두상 돌기(ridged)의 세 가지 형태로 분류되었다. 줄바늘꽃(E. ciliatum subsp.
ciliatum)은 속내에서 유일하게 이랑형(ridged) 배열의 유두상 돌기를 지녀 차이가 있었다. 돌바늘꽃(E. amurense subsp. cephalostigma)은 잎의 모양, 털의 형태와 분포, 식물체의 크기와 습성 등에서 호바늘꽃(E. amurense subsp.
amurense)과 잘 구별되었다. 회령바늘꽃(E. fastigiatoramosum)과 버들바늘꽃(E. palustre)은 잎이 전연인 점에서 비슷하나, 전자는 잎이 긴 타원형이며 주두가 두상이고 종자의 표면무늬가 유두상인 특징을 보이며, 후자는 잎 이 긴 피침형이며 주두가 곤봉형이고 종자의 표면무늬가 그물상인 특징을 가져 잘 구별되었다. 그동안 보고되 지 않았던 가는민바늘꽃(E. platystigmatosum)을 새롭게 포함시켰다.
주요어: 바늘꽃속, 분홍바늘꽃속, 바늘꽃족, 외부형태학적 형질, 종자의 표면무늬
바늘꽃과(Onagraceae Juss.)의 바늘꽃족(Epilobieae Endl.) 은 2속 약 172종으로 구성되며(Baum et al., 1994; Levin et al., 2004), 전 세계에 넓게 분포한다(Chen et al., 2007). 한국산 바늘꽃족 식물들은 분홍바늘꽃(Chamerion angustifolium (L.) Holub subsp. angustifolium)을 제외하고는 모두 바늘꽃 속(Epilobium L.)에 포함된다. 분홍바늘꽃은 많은 학자들에 의해서 Epilobium속(Nakai, 1909; Chung, 1957; Song et al., 1974; Oh, 1983; Do and Im, 1988; Lee, 1996a, b; Lee, 1999;
Lee, 2006) 또는 Chamaenerion속(Chung, 1965; Im, 1976; Kim et al., 1988; Im et al., 1998; Iwatsuki et al., 2002) 으로 분류되 어 혼란이 있었다.
한국산 바늘꽃족 식물들은 학자에 따라 분류군에 대한 인식이 매우 다양하여, 적게는 6분류군부터 많게는 16분류 군까지 분포하는 것으로 기록되어 왔다(Nakai, 1909; Chung, 1957, 1965; Song et al., 1974; Im, 1976; Oh, 1983; Do and Im, 1988; Kim et al., 1988; Lee, 1996a, b; Im et al., 1998; Lee, 1999;
Lee, 2006). 분류군 한계에 대한 학자들 간의 견해 차이는 한 국산 바늘꽃족 종들의 분류학적 연구가 미진했기 때문인 것으로 생각된다. 예를 들면, 동일한 학명에 대해서 다른 국명이 사용되는 경우가 있었다. E. amurense Hausskn.는 호 바늘꽃(Chung, 1957, 1965; ; Oh, 1983; Lee, 1996a, b; Lee, 1999; Lee, 2006), 둥근잎바늘꽃(Im, 1976; Do and Im, 1988; Im et al., 1998), 큰바늘꽃(Song et al., 1974)으로 기록되어 혼란 이 있어왔다. 같은 식물에 여러 학명이 사용되는 경우도 있 었다. 넓은잎바늘꽃은 E. nudicarpum(Chung, 1957, 1965;
Song et al., 1974; Im, 1976; Do and Im, 1988; Kim et al., 1988;
Im et al., 1998) 혹은 E. cephalostigma Hausskn. var. nudicarpum Hara(Oh, 1983; Lee, 1996a,b; Lee, 1999; Lee, 2006) 으로 기록 되었으나, Iwatsuki et al.(2002)과 Chen et al.(1992)은 돌바늘 꽃(E. amurense subsp. cephalostigma Hausskn.)과 동일 종으로 취급하였다. 한라바늘꽃은 E. pyrricholophum var.
curvatopilosum Hara(Song et al., 1974; Oh, 1983; Lee,1999; Lee, 2006) 으로 기록되었으나, Iwatsuki et al.(2002)과 Chen et al.(1992) 은 바늘꽃(E. pyrricholophum Fr. & Sav.)과 같은 종으 로 인식하였다.
호바늘꽃과 돌바늘꽃은 그동안 각각 E. amurense Hausskn. 과 E. cephalostigma Hausskn.으로 기록되어 왔다
(Nakai, 1909; Chung, 1957, 1965; Song et al., 1974; Im, 1976;
Oh, 1983; Do and Im, 1988; Kim et al., 1988; Im et al., 1998;
Lee, 1996a, b; Lee, 1999; Lee, 2006). 그러나 Iwatsuki et al.(2002) 과 Chen et al.(1992)은 돌바늘꽃을 호바늘꽃의 아 종(E. amurense subsp. cephalostigma)으로 기재하였다.
E. tenue Komar. 은 장진바늘꽃(Chung, 1965; Lee, 1996a, b;
Lee, 2006), 솜바늘꽃(Chung, 1957), 좀바늘꽃(Im, 1976; Oh, 1983; Lee, 1996a, b; Do and Im, 1988; Im, 1998) 으로 기재되어 왔으나, Chen et al.(1992)은 E. tenue Komar.을 독립된 종으로 취급하지 않고 호바늘꽃(E. amurense subsp. amurense)의 이 명으로 기재하였다. 줄바늘꽃은 Lee(1996a,b), Lee(1999), Lee(2006) 등이 E. glandulosum var. asiaticum Hara으로, Song et al.(1974) 은 E. glandulosum Lehm. var. brevifolium Nakai(멍울 바늘꽃)으로, Im(1976)과 Im et al.(1988)은 E. glandulosum Lehm.(몽울바늘꽃)으로, Oh(1983)은 E. glandulosum var.
asiaticum Hara( 몽골바늘꽃)으로 기재하였다. 그러나 Chen et al.(1992) 과 Iwatsuki et al.(2002)은 줄바늘꽃을 E. ciliatum subsp. ciliatum의 이명으로 기재하였으므로 줄바늘꽃의 정 확한 학명과 국명의 재정립이 필요하다. E. platystigmatosum C. B. Robinson 은 우리나라 어느 도감이나 문헌상 기록이 없 는 종이며, 일본(Iwatsuki et al., 2002)과 중국(Chen et al., 1992, 2007)에 분포한다고 알려진 종이다. 본 종에 대하여 Oh(1958) 는 1958년 9월 경북 비슬산에서 채집한 표본 1개를 돌바늘꽃의 신변종인 가는민바늘꽃으로 기록한 채 성균관 대학교 표본관(SKK)에 보관하였으나, 그 이후에 발표한 한 국산 바늘꽃과 식물에 관한 식물분류 및 지리학적 연구 논 문에는 전혀 언급하지 않았다(Oh, 1983). 따라서 본 식물의 정확한 분류군 실체에 대한 연구가 필요하다.
본 연구에서는 외부형태 형질 및 종자 표면의 미세구조 형질 특성을 파악하여 한국산 바늘꽃족 분류군들의 실체 를 파악하고 한계를 설정하고자 하였으며, 학명과 국명을 재정리하고, 종 검색표를 작성하고자 하였다.
재료 및 방법
본 연구의 재료는 2006년부터 2007년까지 남한 전역과
백두산 지역에서 직접 채집하거나 성균관대학교(SKK)
표본관에 소장된 식물표본을 사용하였으며, 직접 채집한 식물들은 석엽표본으로 제작하여 성균관대학교 식물표 본관(SKK)에 보관하였다. 그 외의 표본들은 국내표본관 (CNNU, KU, KWNU, SNU) 에서 대여하여 재료로 사용하 였다. 학명의 오류, 국명의 오류로 달리 불리었던 개체들 을 가능한 한 모두 확보하여 관찰하려 했으나, 그렇지 못 한 경우에는 그 동안 발표되었던 문헌들을 참고하여 정리 하였다.
외부 형태학적 형질 측정에 사용한 표본은 관찰 표본 중 잎과 꽃, 그리고 열매 모두 완전한 표본만을 선정하여 재 료로 사용하였다. 연구에 사용된 표본 정보를 Appendix 1 에 기재하였다. 석엽표본을 원기재문과 도감 등을 참조하 여 동정하였으며, 이로부터 각 기관별로 형태학적 특징과 주요 식별형질들을 관찰하였다. 형태학적 변이성을 잘 반 영하고 있는 정량형질들을 분류군당 1-20개체씩 측정하 였으며, 측정 결과를 Table 1에 정리하였고, 측정기준을 모 식도로 제시하였다(Fig. 1). 종자의 표면무늬 관찰을 위하 여 건조된 종자를 Au로 이온 증착(JEOL JFC-1100 ion sputter)시켰으며, 주사전자현미경(JEOL JSM-5600)으로 관찰하였다. 외부형태 형질 및 종자 미세구조 연구 결과 를 바탕으로 검색표 및 기재문을 작성하였다. 종자형질 기재에 사용된 용어는 Chen et al.(1992) 을 참조하였고, 외 부형태형질 용어는 Lee(2004)를 따랐다.
결 과
1. 외부형태학적 특징
한국산 Chamerion 속 1분류군과 Epilobium속 8분류군의 영양기관 및 생식기관에 대한 외부 형태학적 형질을 관찰 하고, 아래와 같이 형질별로 유형화 하였다.
1) 식물체의 크기: 전체 높이는 20-120 cm이다(Table 1).
이 중 분홍바늘꽃(Chamerion angustifolium) 이 가장 높으며(70-120 cm), 바늘꽃속내 분류군들의 식물 높 이는 차이가 거의 없었으나, 큰바늘꽃(E. hirsutum)과 버들바늘꽃(E. palustre)의 경우 높이가 100 cm 이상 인 개체들이 관찰된다.
2) 지하부 구조(Underground system): (1) 근경이 발달하지 않고, 줄기 기부에 섬유형의 기는 뿌리가 발달하는 바 늘꽃(E. pyrricholophum)과 버들바늘꽃(E. palustre); (2) 땅속 옆으로 기는 비대한 근경을 갖는 분홍바늘꽃 (Chamerion angustifolium), 돌바늘꽃(E. amurense subsp.
cephalostigma), 큰바늘꽃(E. hirsutum), 줄바늘꽃(E.
ciliatum); (3) 땅속 옆으로 기는 가늘고 짧은 근경을 갖 는 회령바늘꽃(E. fastigiatoramosum), 호바늘꽃(E.
amurense subsp. amurense), 가는민바늘꽃(E. platysti- gmatosum) 으로 구분된다.
3) 줄기(Stem): 대부분이 직립형으로 자란다.
4) 잎(Leaf)
(1) 잎의 외형(Leaf shapes): 1) 장타원형 또는 타원상 피 침형: 분홍바늘꽃; 2) 좁은 장타원형 또는 피침형: 큰 바늘꽃; 3) 난형 또는 난상 피침형: 바늘꽃; 4) 난형 또 는 피침형: 호바늘꽃; 5) 장타원상 피침형, 좁은 난형 또는 좁은 마름모형: 돌바늘꽃; 6) 좁은 피침형: 버들 바늘꽃; 7) 장타원상 피침형 또는 좁고 긴 타원형: 회 령바늘꽃; 8) 선형 또는 좁은 피침형: 가늘바늘꽃; 9) 피침형 또는 좁은 난형: 줄바늘꽃으로 나타난다. 잎 의 외형은 종에 따라 다소 차이가 있을뿐 유형화하 기 힘들다.
(2) 엽두(Leaf apex): 1) 예두: 분홍바늘꽃, 큰바늘꽃, 버 들바늘꽃, 줄바늘꽃; 2) 예두, 아둔두 또는 둔두: 바 늘꽃; 3) 예두 또는 아둔두: 호바늘꽃; 4) 예두 또는 점첨두: 돌바늘꽃; 5) 예두 또는 둔두: 회령바늘꽃, 가는민바늘꽃으로 엽두형질은 바늘꽃과 분류군간 에 다소 구분된다.
(3) 엽연(Leaf margin): 거치가 있는 분류군(바늘꽃, 돌 바늘꽃, 호바늘꽃, 큰바늘꽃, 줄바늘꽃, 가는 민바 늘꽃)과 거치가 없는 분류군(분홍바늘꽃, 버들바 늘꽃, 회령바늘꽃)으로 구분된다.
(4) 엽저(Leaf base): 1) 둔저: 분홍바늘꽃; 2) 아설저: 큰 바늘꽃; 3) 둔저, 원저 또는 아심장저: 바늘꽃; 4) 원 저 또는 넓은 설저: 호바늘꽃; 5) 설저: 돌바늘꽃; 6) 아원저: 버들바늘꽃, 회령바늘꽃; 7) 유저: 가는민 바늘꽃; 8) 원저 또는 아심장저: 줄바늘꽃으로 나타 난다.
(5) 잎의 길이와 너비(Leaf length and width): 분홍바늘꽃 (8.5-13.6 cm)과 큰바늘꽃(4.4-10.3 cm) 잎의 길이는 나머지 분류군들(1.0-7.5 cm)에 비하여 길다(Table 1).
다만 바늘꽃속의 큰바늘꽃의 잎 길이의 변이 폭이
Fig. 1. Morphological characteristics of the Korean tribe EpilobieaeEndl. A. Inflorescence and Leaf; B. Corolla; C. Calyx; D. Fruit (1- length of leaf; 2-width of leaf; 3-length of petiole; 4-length of petal; 5- length of sepal; 6-width of sepal; 7-length of capsule).
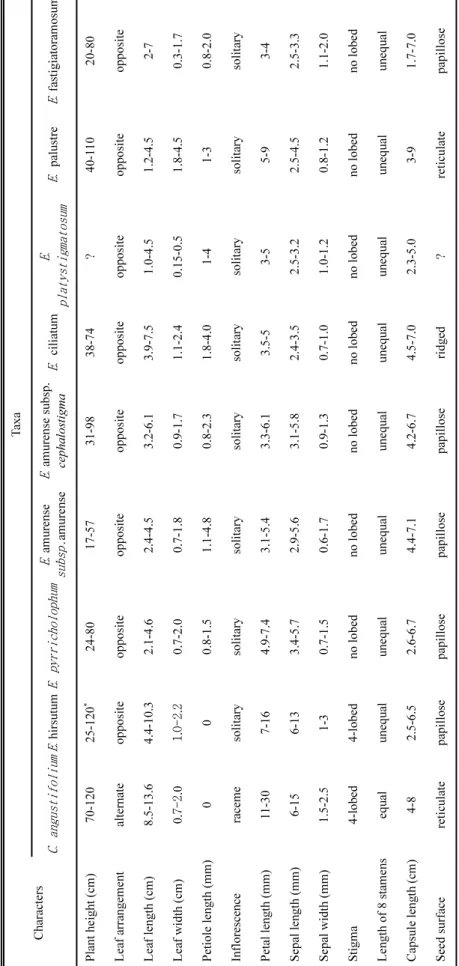
Table 1. Comparisons of morphological and seed characters of the Korean Chamerion (Raf.) Raf. ex Holub and Epilobium L. (Minimum-Maximum; ?: missing data). Characters
Taxa C. angustifoliumE. hirsutumE. pyrricholophumE. amurense subsp.amurenseE. amurense subsp. cephalostigmaE. ciliatumE. platystigmatosumE. palustreE. fastigiatoramosum Plant height (cm)70-12025-120* 24-8017-5731-9838-74?40-11020-80 Leaf arrangementalternateoppositeoppositeoppositeoppositeoppositeoppositeoppositeopposite Leaf length (cm)8.5-13.64.4-10.32.1-4.62.4-4.53.2-6.13.9-7.51.0-4.51.2-4.52-7 Leaf width (cm)0.7-2.01.0-2.20.7-2.00.7-1.80.9-1.71.1-2.40.15-0.51.8-4.50.3-1.7 Petiole length (mm) 000.8-1.51.1-4.80.8-2.31.8-4.01-41-30.8-2.0 Inflorescenceracemesolitarysolitarysolitarysolitarysolitarysolitarysolitarysolitary Petal length (mm)11-307-164.9-7.43.1-5.43.3-6.13.5-53-55-93-4 Sepal length (mm)6-156-133.4-5.72.9-5.63.1-5.82.4-3.52.5-3.22.5-4.52.5-3.3 Sepal width (mm)1.5-2.51-30.7-1.50.6-1.70.9-1.30.7-1.01.0-1.20.8-1.21.1-2.0 Stigma4-lobed4-lobedno lobedno lobedno lobedno lobedno lobedno lobedno lobed Length of 8 stamensequalunequalunequalunequalunequalunequalunequalunequalunequal Capsule length (cm) 4-82.5-6.52.6-6.74.4-7.14.2-6.74.5-7.02.3-5.03-91.7-7.0 Seed surface reticulatepapillosepapillosepapillosepapilloseridged?reticulatepapillose *: obtained from Flora of China (Chen et al., 2007).
크기 때문에 나머지 바늘꽃속 분류군들과 연속변 이를 보이기도 한다. 잎의 너비는 대부분의 분류군 들이 0.7-2.4 cm 정도이나, 가는 민바늘꽃은 0.15-0.5 cm로서 가장 폭이 좁고, 버들바늘꽃은 1.8-4.5 cm로 서 가장 폭이 넓다. 한편 회령바늘꽃의 경우 잎의 너 비가 0.3-1.7 cm로서 변이 폭이 크게 나타난다.
(6) 엽병(Petiole): 엽병이 거의 없는 분류군(분홍바늘꽃, 큰바늘꽃, 바늘꽃)과 짧은 엽병(0.8-4.8 mm)을 가지 는 분류군(호바늘꽃, 돌바늘꽃, 버들바늘꽃, 회령 바늘꽃, 줄바늘꽃, 가는민바늘꽃)으로 구분된다.
5) 화서(Inflorescence): 총상화서를 가지는 분홍바늘꽃과 엽액에 꽃이 한개씩만 달리는 분류군으로 구분된다.
6) 꽃(Flower)
(1) 화판(Petal): 화판끝이 갈라지지 않는 분홍바늘꽃;
두 갈래로 갈라지는 바늘꽃속 분류군으로 나뉜다.
바늘꽃속 큰바늘꽃의 화판 길이는 7-16 mm로서 나 머지 바늘꽃속 분류군들(3-9 mm)에 비하여 길다.
(2) 악편(Sepal): 모든 분류군들의 악편은 4개이며 선형이 고, 뒤로 젖혀지지 않는다(Fig. 1C). 악편의 크기(길이/
너비)는 분홍바늘꽃(6-15/1.5-2.5 mm)과 큰바늘꽃(6- 13/1-3 mm)이 나머지 분류군들(2.4-5.8/0.6-2.0 mm)에 비하여 다소 큰 것으로 나타난다.
(3) 주두(Stigma): 4갈래로 갈라지는 분홍바늘꽃, 큰바
늘꽃; 곤봉 모양인 바늘꽃과 버들바늘꽃; 두상인 호바늘꽃, 돌바늘꽃, 회령바늘꽃, 가는민바늘꽃, 줄바늘꽃으로 나뉜다.
(4) 수술(Stamen):수술대 8개의 길이가 일정한 분홍바 늘꽃; 수술대 4개는 길고, 4개는 짧은 분류군으로 구분된다.
7) 열매(Fruit): 분류군 모두 삭과로 결실기가 되면 4개의 선을 따라 포배열개한다(Fig. 1D). 삭과의 길이는 2.3- 9 cm로서 분류군 내의 변이 폭이 크다. 따라서 분류 군 간에 연속변이로 나타난다.
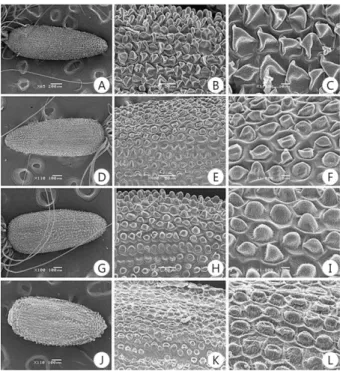
8) 종자(Seed): 분류군의 모든 종자에는 솜털이 존재한 다. 종자의 표면무늬를 전자현미경으로 관찰한 결과 에 의하면(Figs. 2 & 3), (1) 유두상 돌기형(papillose):
바늘꽃, 돌바늘꽃, 호바늘꽃, 큰바늘꽃, 회령바늘꽃;
(2) 그물형(reticulate): 버들바늘꽃, 분홍바늘꽃; (3) 이 랑형(ridged): 줄바늘꽃으로 구분된다.
2. 분류군의 기재
한국산 바늘꽃족의 검색표
1. 줄기의 잎은 호생한다. 꽃은 좌우대칭이며, 수술대 8 개의 길이는 일정하다. 꽃잎은 선단이 갈라지지 않고, 주두는 깊게 4갈래 진다··· Chamerion 분홍바늘꽃속 1. 줄기의 잎은 대생한다. 꽃은 방사상칭이고, 수술대 4 개는 길고, 4개는 짧다. 꽃잎은 선단이 갈라지고, 주 두는 갈라지지 않거나 드물게 4갈래 진다
··· Epilobium 바늘꽃속 1) Chamerion (Raf.) Raf. ex Holub, Folia Geobot.
Phytotax. 7: 85 (1972).
Epilobium subg. Chamerion Rafinesque, Amer. Monthly Mag. & Crit. Rev. 2: 266 (1818). Chamaenerion S. F. Gray, Natur. Arr. Brit. Pl. 559 (1821).
국명: 분홍바늘꽃속
다년초이며 식물체가 곧추 서고, 근경이 옆으로 퍼진다.
줄기는 가지가 갈라지지 않거나 많이 갈라지고, 털이 거 의 없으며 선모도 없다. 잎은 호생하며 드물게 대생한다.
줄기 아랫부분의 잎은 엽병이 없으나 위쪽의 잎은 대개 엽병이 있다. 탁엽과 소포가 없다. 화서는 총상화서이며, 꽃은 다소 좌우대칭이고, 분홍색 또는 홍자색이며 드물게 흰색을 띠기도 한다. 꽃받침과 꽃은 4장이다. 수술은 8개 이며 약은 노란색이다. 자방은 하위이며 암술머리가 깊게 4개로 갈라진다. 열매는 긴 삭과이고 포배열개 한다.
(1) Chamerion angustifolium (L.) Holub, Folia Geobot.
Phytotax. 7: 86 (1972).
Fig. 2. Seed morphology by SEM of the Korean tribe Epilobieae Endl.
A-C: E. pyrricholophum, D-F: E. amurense subsp. cephlostigma, G-I:
E. amurense subsp. amurense, J-L: E. hirstum. A-L: Papillose type.
Magnifications: Bar of A: 200 um, D, G, J: 100 um, B, E, H, K: 50 um, C, F, I, L: 10 um.
Epilobium angustifolium L., Sp. Pl.: 347 (1753).
Chamaenerion angustifolium (L.) Scop., Fl. Carniol. ed. 2, 1: 271 (1772).
Epilobium spicatum Lam., Fl. Franc 3: 482 (1778).
국명: 분홍바늘꽃(Chung, 1957, 1965; Song et al., 1974;
Im, 1976; Do and Im, 1998; Kim et al., 1988; Im et al., 1998;
Lee, 1996a,b; Lee, 1999; Lee, 2006)
분홍바늘꽃의 높이는 70-120 cm에 달하고, 근경이 옆으 로 길게 벋으며 군집을 형성한다. 식물체에는 전체적으로 털이 없다. 잎은 거의 호생하고, 장타원형 또는 타원상 피 침형이며 길이 8.5-13.6 cm, 너비 7-20 mm이고, 예두이고, 둔저이며, 엽연에는 거치가 없다. 엽병이 거의 없다. 총상 화서는 7-8월에 원줄기 끝에 생기며 길이 9-19 cm, 너비는 3-11 mm이다. 꽃잎은 홍자색이며, 4장이고, 역심장형이거 나 긴 마름모형이며 선단이 갈라지지 않는다. 꽃받침잎은 4 장이고 선형이며, 길이는 6-15 mm, 너비는 1.5-2.5 mm이 다. 수술은 8개이고, 길이가 거의 비슷하며 수술대 길이는 7-14 mm이고, 약의 길이는 2-2.5 mm이다. 암술머리는 깊 게 4개로 갈라진다. 삭과는 길이 4-8 cm이다. 종자는 장타 원상 도란형이며 표면에는 그물형의 돌기가 존재하고, 갈 색 솜털이 붙어 있다(Fig. 3: G-I).
분포: 강원도 이북, 백두산.
분류학적 검토: Chamerion속은 형태학적 형질 이외에도 핵유전자와 염록체유전자를 이용한 분자계통학적 연구 에서 Epilobium속의 자매군으로서 독립된 분계조를 형성 하며 잘 구분되어 독립적인 속으로 인정되었다(Levin et al., 2003, 2004).
2) Epilobium L., Sp. Pl. 1: 347-348 (1753) 국명: 바늘꽃속
다년생 초본이며, 로제트형의 근생엽이 존재하고 옆으 로 기는 근경이 발달하거나 그렇지 않다. 줄기에는 털이 없는 것에서부터 많은 것까지 있으며, 털이 있는 경우에 는 엽병 기부에서부터 줄기 아래쪽까지 존재한다. 꽃이 달린 가지 아랫쪽 줄기의 잎은 대생하며, 엽병이 있거나 없다. 탁엽과 소포가 없다. 화서는 엽액에 꽃이 1개씩만 달 리는 단정화서이거나 또는 총상화서를 이룬다. 꽃은 분홍 색-적자색 또는 흰색이고 방사대칭이며, 화통이 있다. 꽃 잎은 4장이고, 도심장형 또는 긴 마름모형이고, 꽃잎 선단 이 두 갈래로 살짝 갈라진다. 수술은 8개인데 4개는 길고 4 개는 짧으며, 약은 노란색이다. 암술대는 곧추서며, 주두 는 큰바늘꽃(E. hirsutum)을 제외하고는 갈라지지 않는다.
열매는 4개로 포배열개하는 삭과이며, 길이가 1.7-9 cm로 서 길고 가늘다. 종자 표면에는 유두상 돌기, 그물형 돌기, 이랑형 배열의 유두상 돌기가 존재하며, 종자 기부에는 흰색, 갈색, 적갈색, 황갈색의 솜털이 많이 난다.
한국산 바늘꽃속의 종 검색표
1. 주두는 4개로 깊게 갈라진다·· (1) E. hirsutum 큰바늘꽃 1. 주두는 갈라지지 않는다
2. 잎에는 분명한 거치가 있다
3. 잎 한쪽면의 거치수가 8개 이상이고, 잎은 좁은 난 형, 피침형-장타원상 피침형이다
4. 주두는 곤봉상이다··· (2) E. pyrricholophum 바늘꽃 4. 주두는 두상이다
5 종자의 표면에는 유두상의 돌기가 발달한다 6. 줄기에는 두 줄의 털이 촘촘하게 분포하며, 악
편에도 털이 많고, 잎은 난형이다
··· (3) E. amurense subsp. amurense 호바늘꽃 6. 줄기와 엽병이 연결되는 마디부분과 악편에는 털이 드물게 분포하고, 잎은 장타원형, 피침형, 또는 좁은 난형이다
·· (4) E. amurense subsp. cephalostigma 돌바늘꽃 5. 종자의 표면은 이랑형이다··(5) E. ciliatum 줄바늘꽃 3. 잎 한쪽면의 거치수는 3-8개이며, 선형 또는 좁은 피 침형이다··· (6) E. platystigmatosum 가는민바늘꽃 2. 잎은 전연이거나 불분명한 거치가 있다
Fig. 3. Seed morphology by SEM of the Korean tribe Epilobieae Endl. A-C: E. fastigiatoramosum, D-F: E. palustre, G-I: C.
angustifolium subsp. angustifolium, J-L: E. ciliatum subsp. ciliatum.
A-C: Papillose type; D-I: Reticulate type; J-L: Ridged type.
Magnifications: same as in Fig. 2. Bar of A,D: 200 um, G, J: 100um, B, E, H, K: 50 um, C, F, I, L: 10um.
7. 잎은 긴 피침형이며, 너비는 1.8-4.5 mm이다
··· (7) E. palustre 버들바늘꽃 7. 잎은 긴 타원형이며, 너비는 3-17 mm이다
··· (8) E. fastigiatoramosum 회령바늘꽃
(1) Epilobium hirsutum Linnaeus, Sp. Pl. 1: 347 (1753) Chamaenerion hirsutum (Linnaeus) Scopoli, Fl. Carniol. ed.
2, 1: 270 (1772).
E. villosum Thunberg, Prodromus 75 (1794).
E. hirsutum var. villosum (Thunberg) H. Hara, J. Jap. Bot.
18: 178 (1942).
E. tomentosum Ventenat, Descr. Pl. Nouv. Jard. Cels. t. 90 (1802).
E. hirsutum var. tomentosum (Ventenat) Boissier, Fl. Orient.
2: 746 (1872).
E. velutinum Nevski, Trudy Bot. Inst. Akad. Nauk SSSR, Ser. 1. Fl. Sist. Vyss. Rast. 4: 312 (1937).
E. hirsutum var. sericeum Bentham ex C. B. Clarke in J. D.
Hooker, Fl. Brit. India 2: 584 (1879).
E. hirsutum var. laetum Wallich ex C. B. Clarke in J. D.
Hooker, Fl. Brit. India 2: 584 (1879).
E. nassirelmulcii Stapf, Denkschr. Kaiserl. Akad. Wiss. Wien Math.-Naturw. Kl. 51: 325 (1886).
국명: 큰바늘꽃(Chung, 1957, 1965; Song et al., 1974; Im, 1976; Do and Im, 1988; Kim et al., 1988; Im et al., 1998; Lee, 1996a,b; Lee, 1999; Lee, 2006)
다년초로서 높이가 1 m 또는 그 이상에 달하며, 원줄기 는 강하고 곧추 자란다. 가지가 많이 갈라지며, 길게 퍼진 털과 선모가 밀생하며, 근경이 옆으로 길게 벋는다. 잎은 대생하고, 좁은 장타원형 또는 피침형이고, 예두이며 아설 저이고, 잎의 기부가 좁아지면서 다소 원줄기를 감싼다.
잎의 길이는 4.4-10.3 cm, 너비는 10-22 mm이며, 양면에 긴 털이 있고, 엽연에는 날카로운 거치가 있으며 잎 한쪽면 거치의 수는 20-59개로 촘촘하게 분포한다. 엽병은 거의 없다. 화서는 엽액에서 꽃이 1개씩만 달리는 단정화서를 갖는다. 꽃은 7-8월에 피며, 짙은 보라색이다. 꽃받침은 길 이 6-13 mm이며, 너비는 1-3 mm이다. 꽃잎은 넓은 도란형 이며 길이 7-16 mm, 너비는 9-12 mm 이며, 꽃잎 끝이 2개 로 갈라진다. 수술은 8개이며, 긴 수술대 길이는 7.2- 8.1 mm, 짧은 수술대 길이는 2.5-3.9 mm이고, 약의 길이는 1-3 mm, 너비는 0.6-1.2 mm이다. 주두는 깊게 4개로 갈라 지며, 주두의 길이는 1.1-3.8 mm이고, 암술대 길이는 5-9 mm 이다. 삭과는 길이 2.5-6.5 cm이다. 종자는 도란형이고, 종자 표면에는 유두상 돌기가 밀생하며, 기부에는 어두운 흰색의 솜털이 존재한다(Fig. 2: J-L).
분포: 백두산 주변의 북부지방, 울릉도, 강원 삼척
분류학적 검토: 큰바늘꽃은 식물 전체에 거센 털과 잎에 예리한 거치가 있으며, 주두가 네 개로 나누어지는 특징으 로 속내 다른 종들과 쉽게 구분된다. 남한에서는 울릉도 및 강원 삼척지역에 분포한다. 남한에서의 분포지역이 지 극히 한정되어 있는 종으로서 집단 유전학적 연구에 기반 한 유전적 다양성 분석 및 추가적인 분포지역 탐색을 통하 여 본 종의 보존에 대한 평가가 필요하다고 판단된다.
(2) Epilobium pyrricholophum Franchet & P. A. L.
Savatier, Enum. Pl. Jap. 2: 370 (1879). (Fig. 4).
E. tetragonum var. japonica Mique, Ann. Mus. Bot.
Lugduno-Batavum 3: 94 (1867).
E. japonicum (Miquel) Haussknecht, Oesterr. Bot. Zeitschr.
29: 56 (1879).
E. pyrricholophum var. japonicum (Miquel) H. Hara, J. Jap.
Bot. 18: 236 (1942).
E. japonicum â. glanduloso-pubescens Haussknecht, Oesterr.
Bot. Zeitschr. 29: 56 (1879).
E. oligodontum Haussknecht, Oesterr. Bot. Zeitschr. 29: 58 (1879).
E. rouyeanum H. Lèveillè, Bull. Acad. Int. Gèogr. Bot. 9:
210 (1900).
E. hakkodense H. Lèveillè, Bull. Acad. Int. Gèogr. Bot. 10:
34 (1901).
E. quadrangulum H. Lèveillè, Bull. Soc. Agric. Sarthe 40:
Fig. 4. Habit of Epilobium pyrricholophum.
72 (1905).
E. makinoense H. Lèveillè, Bull. Soc. Agric. Sarthe 40: 73 (1905).
E. chrysocoma H. Lèveillè, Bull. Herb. Boiss., sèr. 2, 7: 589 (1907).
E. arcuatum H. Lèveillè, Bull. Herb. Boiss., sèr. 2, 7: 589 (1907).
E. prostratum H. Lèveillè, Bull. Soc. France 54: 520, 522 (1907) non Epilobium prostratum Warburg (1893).
E. kiusianum Nakai, Bot. Mag. (Tokyo) 22: 84 (1908).
E. pyrricholophum var. anuoleucholophum H. Lèveillè, Repert. Spec. Nov. Regni Veg. 6: 330 (1909).
E. nakaianum H. Lèveillè, Icon. Gen. Epil. 166, t. 140 (1910).
E. axillare Franchet ex Koidzumi, Fl. Symb. Orient.-Asiat.
85 (1930).
E. myokoense Koidzumi, Fl. Symb. Orient.-Asiat. 86 (1930).
E. pyrricholophum var. japonicum (Miq.) H. Hara, J. Jap.
Bot. 18: 236 (1942).
E. pyrricholophum var. curvatopilosum H. Hara, J. Jap. Bot.
18: 237 (1942).
E. pyrricholophum f. kiusianum (Nakai) Nakai, J. Jap. Bot.
25: 150 (1950)
국명: 바늘꽃(Chung, 1957, 1965; Song et al., 1974; Im, 1976; Do and Im, 1988; Kim et al., 1988; Im et al., 1998; Lee, 1996a,b; Lee, 1999; Lee, 2006).
다년초로서 높이 24-80 cm이고, 섬유형의 뿌리가 발달하 며 식물 전체에 털이 많다. 잎은 대생하며, 잎의 기부가 다 소 원줄기를 감싼다. 잎의 길이는 2.1-4.6 cm, 너비는 0.7-2.0 cm 이며, 난형 또는 난상 피침형이고, 엽두는 예두, 아둔두 또는 둔두이며, 엽저는 둔저, 원저 또는 아심장저이다. 엽연 에는 불규칙한 톱니가 있으며 한쪽면의 거치수는 8-13개이 다. 엽병은 거의 없거나 0.8-1.5 mm로 아주 짧다. 화서는 엽 액에서 꽃이 1개씩만 달리는 단정화서이다. 꽃은 7-9월에 피며, 분홍색 또는 자주색이고, 꽃받침과 꽃잎의 수는 각각 4 개이다. 꽃받침 길이는 3.4-5.7 mm이고, 너비는 0.7-1.5 mm 이다. 꽃잎 길이는 4.9-7.4 mm이며, 꽃잎의 정단이 2개로 얕 게 갈라진다. 수술은 8개이며, 긴 수술대 길이는 2.1-3.3 mm, 짧은 수술대 길이는 1.1-2.6 mm이고, 약의 길이는 0.7-1.1 mm, 너비는 0.3-0.9 mm이다. 자방은 하위이며, 주두는 곤봉 형이고, 주두 길이는 0.7-2.8 mm, 너비는 0.5-1.5 mm이며, 암 술대 길이는 1.5-3.7 mm이다. 삭과는 길이 2.6-6.7 cm이다. 종 자는 긴 난형이고 끝이 둥글며, 종자의 표면에는 유두상 돌 기가 밀생하고(Fig. 2: A-C), 기부에는 적갈색 털이 있다.
분포: 전국
분류학적 검토: 바늘꽃은 다른 종들과 다르게 근경에서 새로운 어린 줄기가 발달한다.
Hara(1942) 는 원줄기와 잎, 그리고 자방와 화경에 굽은 털과 선모가 있는 것을 바늘꽃의 신변종인 한라바늘꽃(E.
pyrricholophum var. curvatopilosum)으로 발표하였다. 그러 나 한라산 및 제주도 내륙에서 채집한 집단과 육지에서 채집한 바늘꽃 집단을 모두 관찰한 결과 원줄기와 잎에는 굽은 털이 존재하였으며 선모는 관찰되지 않았다. 또한 Chen et al.(1992)은 이미 var. curvatopilosum를 var.
pyrricholophum의 이명으로 기재하였으며, Iwatsuki et al.(2002) 또한 이를 따랐다. 따라서 한라바늘꽃은 원변종 인 E. pyrricholophum에 포함되어야 함이 타당한 듯하다.
제주도 한라산에 분포하는 바늘꽃에 대한 정확한 분류군 실체를 파악하기 위해 분자형질 등을 이용한 추가적인 분 류학적인 연구가 필요하다.
(3) Epilobium amurense Haussknecht, Oesterr. Bot.
Zeitschr. 29: 55 (1879) subsp. amurense (Fig. 5).
E. laetum Wallich ex Haussknecht, Monogr. Epil. 218.
Fig. 5. Habit of Epilobium amurense subsp. amurense.
(1884).
E. nepalense Haussknecht. Oesterr. Bot. Zeitschr. 29: 53 (1879).
E. tenue Komarov, Trudy Imp. S.-Peterburgsk. Bot. Sada 25: 95 (1905).
E. yabei H. Lèveillè, Bull. Soc. Agric. Sarthe 40: 72 (1905).
E. gansuense H. Lèveillè, Bull. Herb. Boiss., sèr. 2, 7: 590 (1907).
E. miyabe H. Lèveillè, Repert. Spec. Nov. Regni Veg. 5: 8 (1908).
E. ovale Takeda, J. Linn. Soc., Bot. 42: 466 (1914).
국명: 호바늘꽃(Chung, 1957, 1965; Im, 1976; Lee, 1996a,b; Lee, 1999; Lee, 2006), 큰바늘꽃(Song et al., 1974), 둥근잎바늘꽃(Do and Im, 1998, Im et al., 1998).
다소 곧추 선 다년초로서 높이가 17-57 cm이고, 땅속 기 는 줄기를 형성하며, 줄기 기부에는 로제트형의 짧은 근생 엽이 존재한다. 가지가 갈라지지 않거나 또는 드물게 가지 가 갈라지기도 한다. 줄기에는 두 줄의 구부러진 털이 촘 촘히 나 있다. 잎은 길이 2.4-4.5 cm, 너비 0.7-1.8 cm이며, 난 형 또는 피침형이고, 엽두는 예두 또는 아둔두이고, 엽저 는 원저에서 넓은 설저이다. 엽연에는 날카로운 거치가 있 고 한쪽면 거치의 수는 10-25개이다. 엽병은 거의 없으나 있는 경우에는 길이 1.1-4.8 mm로 짧다. 화서는 직립하고, 엽액에 꽃이 1개씩만 달리는 단정화서이다. 꽃은 7-8월에 피며, 꽃잎은 흰색, 장밋빛 또는 자주색이다. 꽃받침은 길 이 2.9-5.6 mm이며, 너비 0.6-1.7 mm이고, 털이 밀생한다.
꽃잎의 길이는 3.1-5.4 mm이다. 암술대 길이는 0.9-2.9 mm 이며, 주두는 다소 두상이고 주두 길이는 1.1-2.2 mm, 너비 는 0.4-1.9 mm이다. 수술은 8개이며 긴 수술대 길이는1.3- 3.9 mm, 짧은 수술대 길이는 0.7-2.8 mm이며, 약의 길이는 0.5-1.1 mm, 너비는 0.2-0.5 mm이다. 삭과는 길이 4.4-7.1 cm 이다. 종자는 난형이고, 표면에는 유두상 돌기가 있다. 종 자에는 암갈색의 솜털이 붙어 있다(Fig. 2: G-I).
분포: 전국
분류학적 검토: 호바늘꽃은 돌바늘꽃과 형태적으로 비 슷하지만, 전체적으로 줄기에 두 줄로 배열된 구부러진 털이 촘촘히 나 있고, 줄기가 거의 가지 치지 않으며, 식물 의 크기가 작아 구분된다(Table 1). 또한 돌바늘꽃에 비하 여 다소 기울어져 자라고 줄기가 연하며, 뿌리 근처에 로 제트형의 어린잎이 모여 난다.
(4) Epilobium amurense Haussknecht subsp.
cephalostigma (Haussknecht) Chen et al., Syst. Bot. Monogr.
34: 127 (1992). (Fig. 6).
E. cephalostigma Haussknecht, Oesterr. Bot. Zeitschr. 29:
57 (1879).
E. calycinum Haussknecht, Monogr. Epil. 196 (1884).
E. nudicarpum Komarov, Trudy Imp. S.-Peterburgsk. Bot.
Sada 18: 432 (1901).
E. cephalostigma var. nudicarpum (Komarov) H. Hara, J.
Jap. Bot. 18: 234 (1942).
E. consimile var. japonicum Nakai, Bot. Mag. (Tokyo) 25:
148 (1911).
E. sugaharai Koidzumi, Acta Phytotax. Geobot. 5: 121 (1936).
국명: 돌바늘꽃(Chung, 1957, 1965; Song et al., 1974; Im, 1976; Do and Im, 1988; Kim et al., 1988; Im et al., 1998; Lee, 1996a,b; Lee, 1999; Lee, 2006).
곧추 선 다년초이며 높이는 31-98 cm이고, 근경이 발달 하고, 주로 윗부분에서 가지가 많이 갈라진다. 전체적으 로 줄기는 무모이나, 줄기와 엽병이 연결되는 마디부분과 악편에는 털이 드물게 분포한다. 잎은 길이 3.2-6.1 cm, 너 비 0.9-1.7 cm이며, 장타원상 피침형, 좁은 난형 또는 드물 게 좁은 마름모형이고, 엽두는 예두 또는 점첨두, 엽저는 설저이며, 엽연에는 치아상 톱니가 한쪽면에 16-35개 있
Fig. 6. Habit of Epilobium amurense subsp. cephalostigma.
다. 엽병은 거의 없거나 또는 있는 경우 길이는 0.8-2.3 mm 이다. 화서는 다소 직립하고, 윗부분의 엽액에 1개씩의 꽃 이 달리는 단정화서이다. 꽃은 7-8월에 피며, 꽃잎은 분홍 색, 장밋빛, 또는 자주색이며, 드물지만 흰색도 있다. 꽃받 침은 길이 3.1-5.8 mm, 너비 0.9-1.3 mm이고, 털이 조금 있 다. 꽃잎의 길이는 3.3-6.1 mm이다. 수술은 8개이며, 긴 수 술대 길이는 1.7-3.1 mm이고, 짧은 수술대 길이는 0.9-2.2 mm 이고, 약은 길이 0.5-1.0 mm, 너비는 0.3-0.8 mm이다. 암 술대의 길이는 1.2-2.6 mm이고, 주두는 다소 두상이며, 주 두의 길이는 0.9-2.2 mm, 너비는 0.7-1.3 mm이다. 삭과는 길이 4.2-6.7 cm이다. 종자는 긴 난형이며, 종자 표면에는 유두상의 작은 돌기가 있고, 종자에 붙어 있는 솜털은 옅 은 흰색이다(Fig. 2: D-F).
분포: 전국
분류학적 검토: 돌바늘꽃은 호바늘꽃과 형태적으로 비 슷하나 직립하며, 호바늘꽃보다 키가 크고 줄기도 굵고 강하다. 또한 호바늘꽃에 비하여 돌바늘꽃은 엽연이 날카 롭고 표면은 거칠며, 털은 주로 잎이 달린 마디 주변에 나 고, 전체적으로 털이 없다(Table 1).
한편 넓은잎바늘꽃(E. cephalostigma Hausskn. var.
nudicarpum (Komarov) Hara), 두메바늘꽃(E. angulatum Komarov), 명천바늘꽃(E. cylindrostigma Komarov), 흰털바 늘꽃(E. coreanum H. Lèveillè)은 Iwatsuki et al.(2002)와 Chen et al.(1992) 에 의해서 돌바늘꽃의 이명으로 처리되었다.
(5) Epilobium ciliatum Rafinesque, Med. Repos. . 5: 361 (1808). (Fig. 7).
E. glandulosum Lehmann, Pugill. 2: 14 (1830).
E. maximowiczii Haussknecht, Oesterr. Bot. Zeitschr. 29: 57 (1879).
E. glandulosum for. brevifolia Haussknecht, Monogr. Epilob.
273 (1884).
E. punctatum H. Lèveillè, Bull. Acad. Geogr. Bot. 11: 316 (1902).
E. kurilense Nakai, Bot. Mag. (Tokyo) 22: 83 (1908).
E. glandulosum var. kurilense (Nakai) H. Hara, J. Jap. Bot.
18: 238 (1942).
E. glandulosum var. asiaticum H. Hara, J. Jap. Bot. 18: 241 (1942).
E. glandulosum var. brevifolium (Hausskn.) Nakai in Bull.
Nat. Sci. Mus. Tokyo 31: 81 (1952).
국명: 줄바늘꽃(Lee, 1996a,b; Lee, 1999; Lee, 2006), 멍울 바늘꽃(Song et al., 1974), 몽울바늘꽃(Im, 1976; Im et al., 1998).
다년초로서 높이 38-74 cm이고, 근경을 가지고 있다. 원 줄기는 곧추 자라며 능선이 있고 위쪽부분에서 가지가 많
이 갈라지며, 줄기 밑부분에는 털이 없거나 또는 능선 위 에만 털이 있고 윗부분에는 털이 많다. 잎은 대생하며, 소 엽은 피침형 또는 좁은 난형이고, 예두이며, 원저 또는 아 심장저이다. 엽연에는 불규칙한 잔톱니가 있고, 한쪽면 거치의 수는 13-33개이다. 잎의 길이는 3.9-7.5 cm, 너비는 1.1-2.4 cm이며, 엽병의 길이는 1.8-4.0 mm이다. 꽃은 7-8월 에 피며, 분홍색에서부터 적자색이다. 화서는 윗부분의 엽액에 1개씩 꽃이 달리는 단정화서이다. 꽃받침잎은 길 이 2.4-3.5 mm, 너비 0.7-1.0 mm이다. 꽃잎의 길이는 3.5- 5 mm, 너비는 2-3 mm이다. 수술은 8개이며, 긴 수술대의 길이는 2.5-3.2 mm이고, 짧은 수술대의 길이는 1.5-2.5 mm 이며, 약의 길이는 0.5-0.6 mm, 너비는 0.25-0.35 mm이다.
주두는 두상이고, 길이는 0.8-2 mm, 너비는 0.6-0.8 mm이 며, 암술대 길이는1.8-3.2 mm이다. 삭과는 길이 4.5-7 cm이 다. 종자는 좁은 타원형이며, 예두이고, 흰색 솜털이 존재 하며, 종자 표면에는 유두상의 작은 돌기가 이랑 모양으 로 존재한다(Fig. 3: J-L).
분포: 우리나라 북부지방 및 백두산
분류학적 검토: 줄바늘꽃으로 기록되어 온 E. glandulosum var. asiaticum Hara(Oh, 1983; Lee, 1996a,b ; Lee, 1999; Lee, 2006) 는 Chen et al.(1992)와 Iwatsuki et al.(2002)에 의해 E. ciliatum subsp. ciliatum 의 이명으로 처리되었다. Oh(1983)가 기록한 섬 바늘꽃(E. glandulosum var. kurilense (Nakai) Hara)은 E. ciliatum subsp. ciliatum 의 이명으로 처리되었다(Iwatsuki et al., 2002).
Fig. 7. Habit of Epilobium ciliatum.
Hoch and Raven(1977)은 E. ciliatum을 세 개의 아종(E.
ciliatum subsp. ciliatum, E. ciliatum subsp. glandulosum, E.
ciliatum subsp. watsonii)으로 나누었다. Chen et al.(1992)은 장백산 부근에서 채집한 식물이 전형적인 subsp.
glandulosum 보다는 꽃의 크기가 작고, subsp. ciliatum과 비 슷하다고 여겨 subsp. ciliatum으로 동정하였다. 이후에 Chen et al.(2007) 은 subsp. glandulosum은 북미에만 분포한 다고 기록하였다. 본 연구를 통해 확보된 장백산 집단의 개체도 subsp. ciliatum으로 동정되었다. 한반도 북쪽 지역 에 서식하는 줄바늘꽃은 바늘꽃속의 다른 분류군들보다 엽병이 긴 편이며, 엽연이 부드러운 예거치이고, 종자 표 면에 유두상의 작은 돌기가 이랑형으로 배열되어 속내 다 른 분류군들과 잘 구분된다.
(6) Epilobium platystigmatosum C. B. Robinson, Philipp.
J. Sci. 3: 210 (1908). (Fig. 8).
E. formosanum Masamune, Trans. Nat. Hist. Soc. Taiwan 29: 62 (1939).
E. cephalostigma var. linearifolium Hisauti, J. Jap. Bot. 14:
143 (1938).
E. sohayakiense Koidzumi, Acta Phytotax. Geobot. 8: 61 (1939).
국명: 가는민바늘꽃(오, 1958).
곧게 선 다년초이며, 근경이 발달하고, 높이는 15-70 cm 정도이다. 줄기는 매끄럽고 무모이다. 잎은 대생하며, 선 형 또는 좁은 피침형이며, 엽두는 예두 또는 둔두이고, 엽 저는 유저이며, 엽연에는 얕은 거치가 있고 한쪽면의 거 치수는 3-8개이다. 잎은 길이 1-4.5 cm, 너비 0.15-0.5 cm이 며, 엽맥이나 잎 가장자리를 제외하고는 털이 없다. 엽병 은 길이 1-4 mm이다. 화서는 윗부분의 엽액에 1개씩 꽃이 달리는 단정화서이다. 꽃은 8-9월에 피며, 흰색 또는 분홍 색이며, 드물게 적자색이다. 꽃받침은 길이 2.5-3.2 mm, 너 비는 1-1.2 mm이고, 장타원형-피침형이다. 꽃잎은 길이 3- 5 mm, 너비는 1.8-2.5 mm이다. 긴 수술대 길이는 1.6-2 mm, 짧은 수술대 길이는 0.6-1.2 mm이고, 약은 길이 0.4- 0.6 mm, 너비 0.25-0.35 mm이다. 주두는 두상이고, 길이는 0.7-1.4 mm, 너비는 0.5-1.3 mm이며, 암술대 길이는 1.8- 2 mm 이다. 삭과는 길이 2.3-5 cm이다.
분포: 경북
분류학적 검토: 본 식물에 대하여 오(1958)는 돌바늘꽃 의 신변종(E. coreanum Leveille var. angustifolia Chung)으로 표본에 처음으로 기록하였다. 그러나 오(1958)는 본 식물 을 기록한 이후에 발표한 논문(Oh, 1983)에 본 식물을 기 록하지 않았으며, 그 뒤로 본 종은 우리나라 어느 문헌에 도 기록되지 않았다. 연구 결과 본 식물은 돌바늘꽃과는 외부형태학적으로 확연한 차이가 있었다. 즉, 돌바늘꽃의 잎은 장타원상 피침형이고, 잎의 거치수가 상당히 많지만 (16-35 개), 가는민바늘꽃의 잎은 선형이며, 잎의 거치수가 적고(3-8개), 잎의 정단 부위에만 거치가 존재한다. 줄기 또한 돌바늘꽃에 비해 가는민바늘꽃은 아주 가는 특징을 지닌다. 이러한 특징에 근거하여 본 식물은 Epilobium platystigmatosum C. B. Robinson 임이 밝혀졌다(chen et al., 1992, 2007; Iwatsuki et al., 2002). 본 종에 대한 추가적인 분 류학적 연구 및 우리 나라 분포지에 대한 상세한 조사가 필요하다고 생각된다.
(7) Epilobium palustre Linnaeus, Sp. Pl. 1: 348 (1753).
(Fig. 9).
E. palustre var. typicum C. B. Clarke in J. D. Hooker, Fl.
Brit. India 2: 585 (1879).
E. rhynchocarpum Boissier, Diagn. ser. 2, 2: 53 (1856).
E. palustre L. var. lavandulaefolium Lecoq et Lamotte ex Hausskn., Monogr. E. 133 (1884).
E. dahuricum auct. :Nakai in Bot. Mag. Tokyo 22: 74 & 83 (1908).
E. fischerianum Pavlov, Bjull. Moskovsk. Obsc. Isp. Prir.,
Fig. 8. Habit of Epilobium platystigmatosum.Otd. Biol. n. s., 38: 105 (1929).
E. palustre var. majus C. B. Clarke in J. D. Hooker, Fl. Brit.
India 2: 585 (1879).
E. palustre var. minimum C. B. Clarke in J. D. Hooker, Fl.
Brit. India 2: 586 (1879).
국명: 버들바늘꽃(Im, 1976; Do and Im, 1988; Lee, 1996a,b; Im et al., 1998; Lee, 1999; Lee, 2006), 좀버들바늘 꽃(Song et al., 1974; Im, 1976; Im et al., 1998).
다년초로서 높이 40-110 cm이고, 기부에 섬유상의 뿌리 가 발달한다. 줄기는 보통 곁가지가 없으나 때로는 곁가 지가 발달하기도 한다. 잎은 좁은 피침형이며, 예두이고 아원저이며, 엽연에는 거치가 없고, 잎의 길이는 1.2-4.5 cm, 너비는 1.8-4.5 mm이다. 엽병은 1-3 mm이거나 또는 없 다. 화서는 엽액에서 꽃이 1개씩만 달리는 단정화서이다.
꽃은 7-8월에 피며 연한홍색이고, 소화경에는 굽은 털이 밀생한다. 꽃받침은 길이 2.5-4.5 mm, 너비는 0.8-1.2 mm이 다. 꽃잎은 길이 5-9 mm 이고, 폭 2-3 mm이다. 수술은 8개 이며, 수술대 길이는 긴 것은 2-2.8 mm이고, 짧은 것은 1.2- 1.5 mm이며, 약의 길이는 0.43-0.74 mm, 너비는 0.16-0.51
mm이다. 주두는 곤봉형이며 길이가 0.3-1.2 mm, 너비가 0.3-1.0 mm 이고, 암술대는 길이 1.5-2.3 mm이다. 삭과는 길 이 3-9cm이다. 종자는 긴 타원형이고, 정단이 뾰족하며, 종자의 표면에는 그물형의 돌기가 있고, 갈색 솜털이 붙 어 있다(Fig. 3: D-F).
분포: 전국
분류학적 검토: 버들바늘꽃은 잎에 거치가 없는 점에서 회령바늘꽃과 외형상 비슷하다. 그러나 버들바늘꽃은 잎 이 좁은 피침형(길이 1.2-4.5 cm, 너비 1.8-4.5 mm)인데 비 해, 회령바늘꽃은 잎의 너비가 버들바늘꽃보다 넓으며, 장타원상 피침형이거나 다소 좁은 장타원형(길이 2-7 cm, 너비 0.3-1.7 cm)으로 차이가 있다. 또한 버들바늘꽃의 종 자 표면은 그물형이고(Fig. 3: D-F), 회령바늘꽃의 종자 표 면은 유두상 같은 작은 돌기가 밀생하므로(Fig. 3: A-C) 잘 구분된다.
(8) Epilobium fastigiatoramosum Nakai, Bot. Mag. Tokyo 33: 9 (1919). (Fig. 10).
E. palustre var. mandjuricum Haussknecht, Monogr. Epilob.
134 (1884).
E. baicalense Popov, Bot. Mater. Gerb. Bot. Inst. Komarova
Fig. 9. Habit of Epilobium palustre. Fig. 10. Habit of Epilobium fastigiatoramosum.
Akad. Nauk SSSR 18: 6 (1957).
국명: 회령바늘꽃(Lee, 1996a,b; Lee, 1999; Lee, 2006), 가 지바늘꽃(Song et al., 1974; Im, 1976, Im et al., 1998), 버들 바늘꽃(Chung, 1957, 1965; Song et al., 1974).
다년초이며, 높이는 20-80 cm이고, 줄기는 가지가 많이 갈라지거나 그렇지 않으며, 줄기 윗부분에는 굽은 털이 있다. 잎은 대생하고, 장타원상 피침형이거나 또는 좁은 장타원형이고, 엽두는 예두 또는 둔두이며, 엽저는 아원 저이고 거치가 없다. 잎은 길이 2-7 cm, 너비 0.3-1.7 cm로 서 잎 앞면에 굽은 털이 산재하고, 뒷면에는 표면 뿐만 아 니라 주맥과 가장자리에도 굽은 털이 존재한다. 엽병은 거의 없거나 0.8-2 mm로서 매우 짧다. 화서는 엽액에서 꽃 이 1개씩만 달리는 단정화서이다. 꽃은 7-8월에 피며, 하 얀색 또는 연한 홍색이다. 꽃받침은 길이 2.5-3.3 mm, 너비 1-1.2 mm 이고, 꽃잎은 길이 3-4 mm, 너비 1.8-2.5 mm이다.
수술은 8개이며 긴 수술대의 길이는 1.5-2 mm, 짧은 수술 대의 길이는 1-1.5 mm이고, 약의 길이는 0.4-0.6 mm, 너비 는 0.3-0.4 mm이다. 주두는 두상이고 길이 0.9-1.4 mm, 너 비 0.5-0.9 mm이며, 암술대 길이는 1.5-2.5 mm이다. 삭과는 길이 1.7-7 cm이다. 종자는 좁은 난형이고 끝이 다소 둥글 며, 표면에는 유두상의 작은 돌기가 밀생하고, 황갈색의 솜털이 붙어 있다(Fig. 3: A-C).
분포: 우리나라 북부 지방과 백두산 주변 지역
분류학적 검토: 한반도의 회령바늘꽃 분포지역은 북부 지방으로서 전국에 분포하는 버들바늘꽃과 차이가 있다.
고 찰
한국산 바늘꽃족에 대한 외부형태와 종자의 미세구조 형질을 바탕으로 한 분류학적 연구를 수행하였다. 외부형 태학적 연구결과 잎의 외형, 엽연의 거치 여부, 식물체에 서 털의 형태 및 분포, 그리고 식물체의 크기 및 습성 등이 바늘꽃족 분류에 유용한 형질로 밝혀졌다. Chen et al.(1992)에 의해서 바늘꽃족의 분류학적 유용성이 입증된 종자의 표면무늬 또한 일부 한국산 분류군들의 식별에 유 용한 형질이었다. 즉, 바늘꽃속의 줄바늘꽃(이랑형)과 버 들바늘꽃(그물형)의 표면무늬는 바늘꽃속 나머지 분류군 들의 표면무늬(유두상 돌기형)와 잘 구분되었다.
분홍바늘꽃속(Chamerion)의 분홍바늘꽃(Chamerion an- gustifolium) 은 잎이 나선상 호생하고, 수술대 8개의 길이가 비슷하며, 꽃잎 끝이 갈라지지 않는 특징으로 바늘꽃속 (Epilobium) 식물들과 잘 구분되었다. 한국산 바늘꽃속의 경우 큰바늘꽃(E. hirsutum), 바늘꽃(E. pyrricholophum), 호 바늘꽃(E. amurense subsp. amurense), 돌바늘꽃(E. amurense subsp. cephalostigma), 회령바늘꽃(E. fastigiatoramosum), 버 들바늘꽃(E. palustre), 줄바늘꽃(E. ciliatum subsp. ciliatum),
가는민바늘꽃(E. platystigmatosum)의 8분류군으로 정리되 었다. 그러나 가는민바늘꽃(E. platystigmatosum)은 현재 우 리나라 존재여부가 불확실하다. 가는민바늘꽃을 발견한 오(1958)는 본인이 발표한 논문에 본 종을 기재하지 않았 으며(Oh, 1983), 현재까지 우리나라 어느 문헌에도 본 종이 기록된 적 없다. 따라서 가는민바늘꽃의 한반도 존재여부 를 파악하기 위하여 오(1958)에 의해서 발견된 경북지역의 비슬산을 중심으로 한 추가적인 연구가 절실히 요구된다.
호바늘꽃(E. amurense subsp. amurense)은 대부분 고도가 높은 산에 분포하며 식물의 크기가 대체로 작고, 돌바늘 꽃(E. amurense subsp. cephalostigma)에 비해 전체적으로 털 이 많다. 그러나 이러한 특징 외에는 두 분류군은 형태적 으로 매우 유사하다. 돌바늘꽃보다 고도가 높은 곳에서 서식하는 호바늘꽃은 고산 생태계의 강한 바람과 낮은 온 도에 적응하기 위해서 개체의 크기를 줄이고, 식물체에 많은 털을 지니게 되었을 가능성이 있다. 그러나 이를 뒷 받침하기에는 형태적인 형질만으로는 한계가 있으므로, 이들 두 종의 종간 유연관계를 명확히 밝히기 위한 유전 자 분석 등을 이용한 추가적인 연구가 필요하다.
큰바늘꽃(E. hirsutum)은 꽃과 식물체의 크기(1 m 이상) 가 상당히 크고 전체에 강한 털이 존재하며, 잎에 날카로 운 거치가 발달하고, 특히 주두가 4갈래지는 특징을 지닌 다. 이는 한반도의 다른 바늘꽃속 종들에서는 볼 수 없는 큰바늘꽃 만이 가지는 유일한 특징이다. 특히 이러한 주 두 형태는 분홍바늘꽃(Chamerion angustifolium)의 주두와 유사하다. 큰바늘꽃 식물체의 크기(1 m이상) 및 서식지 특 성 또한 분홍바늘꽃(식물 높이 0.7-1.2 m)과 중복되어 두 종의 연관성을 뒷받침해주고 있다. 한반도 식생분포 특성 (Chang et al., 2011)에 의하면 큰바늘꽃은 울릉도 및 만주 구계에 속하고 분홍바늘꽃 또한 만주구계에 포함되고 있 다. 따라서 바늘꽃족에서 큰바늘꽃의 계통학적 중요성을 파악하기 위한 분자형질 등을 이용한 추가적인 연구가 필 요하다고 판단된다.
사 사
본 논문은 한반도 고유 식물자원 검색기술 개발 및 한반 도 식물지 발간(과제번호 052-081-070) 사업의 연구비지 원에 의하여 수행되었습니다. 귀중한 표본을 대여해 주신 CNNU, KU, KWNU, SNU의 표본관 관계자 분들께 깊이 감사드리며, 논문 심사과정 중 많은 조언과 교정에 도움 을 주신 익명의 심사자분들께도 감사드립니다.
인용문헌
Baum, D. A., K. J. Sytsma and P. C. Hoch. 1994. The phylogeny of Epilobium (Onagraceae) based on nuclear ribosomal DNA sequences. Systematic Botany 19: 363-388.
Chang, C. S., H. Kim and K.-S. Chang. 2011. Illustrated Encyclo- pedia of Fauna and Flora of Korea. Vol. 43. Woody Plants.
Designpost Press, Paju. (in Korean)
Chen, C. J., P. C. Hoch and P. H. Raven. 1992. Systematics of Epi- lobium (Onagraceae) in China. Systematic Botany Mono- graphs. 34: 1-209.
Chen, C. J., P. C. Hoch and P. H. Raven. 2007. Epilobium L. In:
Flora of China, Vol.13. Wu, S., P. H. Raven and D. Y. Hong (eds.), Science Press and Missouri Botanical Garden Press, Beijing and St. Louis. Pp. 400-422.
Chung, T. H. 1957. Korean Flora, Part 2, Herbaceous Plants. Shin- jisa, Seoul. Pp. 431-439. (in Korean)
Chung, T. H. 1965. Illustrated Encyclopedia of Fauna and Flora of Korea vol. 5. Tracheophyta. Hyangmunsa, Seoul. Pp. 834-845.
(in Korean)
Do, B. S. and R. J. Im. 1988. Korean Illustrations of Plants. Soc.
Sci. Publ. Co., Pyungyang. Pp. 430-437. (in Korea)
Hara, H. 1942. The Journal of Japanese Botany. Tsumura Labora- tory. Meguro Tokyo. 18: 229-249.
Hoch, P. C. and P. H. Raven. 1977. New combinations in Epilo- bium (Onagraceae). Annals of the Missouri Botanical Garden.
Missouri Botanical Garden Press. 64: 136-136.
Im, R. J. 1976. Flora Coreana 4. Sci. Publ. Co., Pyungyang. Pp.
411-427. (in Korean)
Im, R. J., H. S. Kim, J. S. Gkwak, Y. C. La, Y. J. Li, K. P. Li and K. S. Han. 1998. Flora Coreana (5). The Science and Tech- nology Publishing House, Pyongyang. Pp. 78-95. (in Korean) Iwatsuki, K., D. E., Boufford and H. Ohba. 2002. Onagraceae In:
Flora of Japan, Vol. •±c. Iwatsuki K., D. E. Boufford, and H.
Ohba (eds.), Kodansha, Tokyo. Pp. 224-246.
Kim, H. S., S. J. Lee, H. S. Park and M. K. Kim. 1988. Colored Illustrations of Plants. Sci. Encyclogedia. Jonghap Publ. Co., Pyungyang. Pp. 410-415. (in Korea)
Lee, T. B. 1999. Illustrated Flora of Korea. Hyangmunsa, Seoul.
Pp. 566-570. (in Korean)
Lee, W. T. 1996a. Standard Illustrations of Korean Plants. Acad- emy Publ. Co., Seoul. Pp. 243-246. (in Korean)
Lee, W. T. 1996b. Lineamenta Florae Koreae. Academy Publ. Co., Seoul. Pp. 752-761.
Lee, W.S. 2004. Plant Taxonomy, Wooseong press. Seoul. Pp.
304-420. (in Korean)
Lee, Y. N. 2006. New Flora of Korea, Vol. I. Kyohaksa, Seoul. Pp.
787-793. (in Korean)
Levin, R. A., W. L. Wagner, P. C. Hoch, M. Nepokroeff, J. C.
Pires, E. A. Zimmer and K. J. Sytsma. 2003. Family-level relationships of onagraceae based on chloroplast rbcL and ndhF data, American Journal of Botany 90: 107-115.
Levin, R.A., W. L. Wagner, P. C. Hoch, W. J. Hahn, A. Rodriguez, D. A. Baum, L. Katinas, E. A. Zimmer and K. J. Sytsma.
2004. Paraphyly in Tribe Onagreae: Insights into Phyloge- netic Relationships of Onagraceae Based on Nuclear and Chloroplast Sequence Data. Systematic Botany 29: 147-164.
Nakai, T. 1909. Flora Koreana I, J. Coll. Sci. Imp. Univ. Tokyo.
Pp. 239-247.
Oh, S. Y. 1983. Enmerative and Phytogeographical Studies of Family Onagraceae in Korea. Research review of Kyungpook National University 35: 291-362.
Song, J. T., M. K. Park and Y. C. Kim. 1974. Korean Resources Flora. Gukjaemunhwasa, Seoul. Pp. 506-513. (in Korea)