http://dx.doi.org/10.4014/kjmb.1207.07024 pISSN 1598-642X eISSN 2234-7305
카사바 당화액을 이용한 실험실용 및 산업용 효모의 에탄올 발효성능 비교
진영욱
1
†·김진우1
†·박용철2
*·서진호1
*1서울대학교 농생명공학부, 2국민대학교 발효융합과
Received : July 30, 2012 / Revised : August 2, 2012 / Accepted : August 22, 2012
Comparison of Ethanol Fermentation Properties between Laboratorial and Industrial Yeast Strains using Cassava Hydrolysate. Chin, Young-Wook1†, Jin-Woo Kim1†, Yong-Cheol Park2*, and Jin-Ho Seo1*.
1Department of Agricultural Biotechnology, Seoul National University, Seoul 151-742, Korea, 2Department of Advanced Fermentation Fusion Science and Technology, Kookmin University, Seoul 136-702, Korea − In order to investigate the ethanol fermentation properties of alcohol yeasts a laboratorial strain (CEN.PK2-1D) and two industrial alcohol yeasts (JHS100 and JHS200) of Saccharomyces cerevisiae were cultured in a pure YP medium with 300 g/L glucose and cassava hydrolysate. Spot assay and cell viability tests showed that both the JHS100 and JHS200 strains exhibited higher ethanol tolerance than the CEN.PK2-1D strain. The JHS100 strain demonstrated the highest cell growth, glucose consumption and ethanol production. In particular, an anaerobic batch fermentation of the JHS100 strain using cassava hydrolysate with 250 g/L glucose resulted in a 106.1 g/L ethanol concentration, 0.42 g/g ethanol yield and 3.15 g/L-hr ethanol productivity, which were 53%, 13%, 53% higher than the corresponding values for the CEN.PK2-1D strain. By changing the pure YP medium to cassava hydrolysate, 19% and 17% decreases in ethanol yield and productivity for the CEN.PK2- 1D strain were observed, whereas the cultures of the JHS100 and JHS200 stains showed similar ethanol pro- ductivities and only an 8% decrease in ethanol yield. Furthermore, the JHS100 and JHS200 stains produced lower levels of glycerol and acetate byproducts than the CEN.PK2-1D strain. Consequently, the outstanding ethanol fermentation performance of the industrial strains might be owing to rapid cell growth, high ethanol tolerance, low nitrogen requirements and the low formation of by-products.
Key words: Ethanol, Saccharomyces cerevisiae, laboratorial yeast, industrial yeast, cassava hydrolysate
서 론
세계적으로 화석연료의 고갈과 이산화탄소 배출에 의한 지구온난화로 인해 석유자원을 대체할 수 있는 바이오 연료 에 대한 관심이 급증하고 있다. 이 중, bioethanol은 생산성 과 생산수율이 높아 경제적으로 생산할 수 있고, 인화성이 좋을 뿐만 아니라 가솔린과 혼합하여 이용할 수 있기 때문 에 석유자원의 대체연료로 각광받고 있다[5-6, 8]. 이러한 분 위기 속에서 미국과 브라질은 전분계 바이오매스를 이용한 bioethanol의 생산기술을 가장 먼저 상용화하였다. 하지만 주 요식량자원인 옥수수와 사탕수수 등이 bioethanol 생산의 원 료로 이용되면서 가격이 급등하였고 개발도상국의 기아문제
와 맞물려 윤리적 갈등을 야기하였다[10]. 따라서 이를 대체 할 수 있는 바이오매스를 찾고자 연구가 활발히 진행되고 있 다. 카사바(Manihot esculenta)는 열대성 덩이뿌리 작물로 가 격이 매우 저렴하고 재배와 당화가 용이할 뿐만 아니라 식 량자원으로의 이용도가 낮기 때문에 여러 나라에서 bioetha- nol의 원료로 이용하기 위한 연구들이 시도되고 있다[4, 8, 11].
한편, 원료수급 이외에도 또 하나의 중요한 요소인 생산 균주의 측면에서 ethanol 발효성능이 가장 뛰어나다고 알려 진 알코올효모 Saccharomyces cerevisiae를 이용하여 ethanol 의 수율과 생산성을 향상시키기 위해 방대한 양의 유전적 생 리학적 연구들이 진행되어 왔다. 실제로 이러한 연구들을 통 해 ethanol, 고온, 고농도의 당에 대한 내성 및 ethanol 생산 능력이 향상된 다수의 효모들이 개발되었으나 이 효모들은 세포를 재사용하는 산업적인 조건에서는 향상된 발효능력이 소실되었기 때문에 산업적으로 이용할 수 없었다[13]. 그에 따라 현재 산업적인 bioethanol의 생산에는 무작위 돌연변이 를 통하여 ethanol 발효성능을 향상시킨 S. cerevisiae 변이 주가 주로 이용되고 있는데, 아직까지 이러한 산업용 알코
*Corresponding author J.-H. S.
Tel: +82-2-880-4855, Fax: +82-2-873-5095 E-mail: [email protected]
Y.-C. P.
Tel: +82-2-910-5462, Fax: +82-2-910-5739 E-mail: [email protected]
†
Y.W. Chin and J.W. Kim contributed to this research equally.
올 효모에 대한 연구는 미미한 실정이다[2]. 따라서 본 연구 에서는 산업용 및 실험실용 효모의 생장속도, ethanol stress 에 대한 내성 및 생존능력을 정량적으로 비교하고, 카사바 당화액을 이용한 회분식 발효공정을 통해서 ethanol 생산에 관련된 특성을 비교 분석함으로써 산업적인 ethanol의 생산 에 도움을 주기 위한 기초연구를 수행하였다.
재료 및 방법 균주 및 배양조건
실험실용 효모인 S. cerevisiae CEN.PK2-1D와 (주)진로로부 터 제공받은 산업용 효모인 S. cerevisiae JHS100과 JHS200 을 이용하였다. JHS100과 JHS200은 internal transcribed spacer (ITS) region의 염기서열을 분석 하여 S. cerevisiae임을 확인하였다. 모든 균주는 70%(v/v) YPD배지(20 g/L peptone, 10 g/L yeast extract, 20 g/L glucose)와 30%(v/v) glycerol 으로 구성된 혼합용액에서 -70oC에 보관하였고, 5 mL YPD 배지가 담긴 시험관에서 20시간 동안 30oC와 250 rpm의 조 건에서 진탕 배양시킨 후 종균으로 이용하였다. 플라스크 발 효는 300 g/L glucose가 함유된 100 mL YP배지가 담긴 500 mL baffled flask 이용하여 30oC와 90 rpm의 조건에서 실시하였고, 이 때 종균은 초기 세포 농도(OD600)가 약 20 이 되도록 접종하였다. 발효기를 이용한 발효실험에는 (주) 창해에탄올로부터 제공받은 카사바 당화액만을 배지로 이용 하였고 질소원을 따로 추가하지 않았다. 종균 배양된 효모 를 200 mL YPD배지가 담긴 500 mL 플라스크에서 30시간 동안 30oC와 250 rpm의 조건으로 전배양을 한 후 초기 OD600가 약 20이 되도록 0.5 L의 카사바 당화액에 접종하였 다. 1 L 배양기가 장착된 발효기(Kobiotech KL-1l, Korea)를 이용하였고 30oC와 200 rpm의 조건과 공기를 주입하지 않 는 조건에서 발효하였으며 pH는 2 N NaOH 용액과 2 N HCl 용액을 이용하여 5.5로 유지하였다.
Ethanol 내성 실험
알코올 효모의 ethanol 내성을 비교하기 위하여 spot assay 와 세포 viability를 측정하였다. Spot assay를 위해서 세 효 모를 5 mL YPD배지에서 30oC, 250 rpm의 조건으로 초기대 수증식기에 도달할 때까지 배양한 후, 배양액의 균체량을 증 류수를 이용하여 동일하게 맞추었다. 배양액을 YPD 배지를 이용하여 10에서 105까지 10배 단위로 희석한 후 각각 0%
와 10%(v/v)의 ethanol이 함유된 YPD plate에 7 µL씩 떨어 뜨리고 colony가 형성될 때까지 30oC에서 정치배양하였다.
또한 ethanol에 대한 cell viability 비교 실험을 위해서 동일 과정으로 배양한 효모를 15%(v/v) ethanol이 함유된 5 mL YPD 액체배지에 배양하면서 1.0, 3.0, 6.0, 12.5, 16.0, 22.0 시간에 샘플을 채취하여 세포가 약 2.5×102가 되도록 희석 한 후 YPD plate에 도말하고 30oC에서 생성된 colony의 개
수를 측정하였다.
균체농도 및 대사산물의 농도 측정
균체농도는 분광광도계(Amersham Pharmacia Ultrospec 2000, USA)를 이용하여 600 nm에서 측정하였다. Glucose 및 ethanol, acetate, glycerol을 포함하는 대사산물의 농도는 refractive index (RI) detector가 장착된 HPLC (Agilent 1200 series, USA)를 이용하여 분석하였고, 컬럼은 REZEX ROA- organic acid column (Phenomenex, USA)를 사용하였다. 이 동상으로 5 mM의 황산용액을 0.6 mL/min의 속도로 흘려주 었고, 컬럼 온도는 60oC로 설정하였다. HPLC 샘플은 배양 액 2 mL을 취하고 원심분리기를 이용하여 13,000 rpm으로 1분간 원심분리 시킨 후 상등액을 10배 희석하였고, 이를 필 터링하여 준비하였다.
결과 및 고찰 세포성장속도와 에탄올 내성의 비교
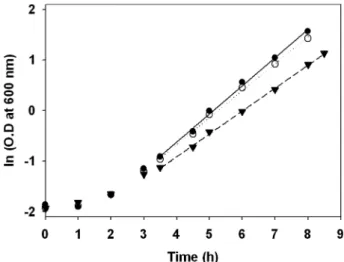
경제적인 ethanol의 생산을 위해서 빠른 세포성장속도와 고농도의 ethanol stress에 대한 내성은 중요한 요소이다[1, 12]. 먼저 ethanol stress가 없는 조건에서 실험실용 효모인 CEN.PK2-1D과 산업용 효모인 JHS100와 JHS200의 비성장 속도(specific growth rate)를 비교하기 위해 플라스크에서 호 기적 배양을 하며 약 1시간 간격으로 균체농도를 측정하였 다. 초기대수증식기인 3.5-8시간 구간에서 CEN.PK2-1D, JHS100, JHS200의 비성장속도를 측정해 본 결과, 각각 0.452/h, 0.551/h, 0.531/h로 JHS100과 JHS200의 성장속도 가 CEN.PK2-1D보다 각각 22%와 17% 더 빨랐다(Fig. 1).
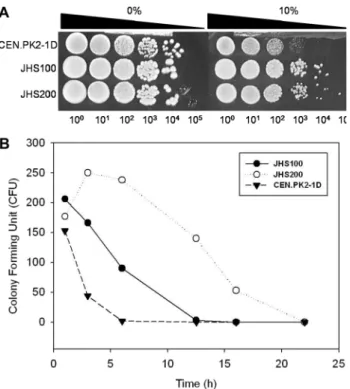
Ethanol stress에 대한 세 효모들의 내성을 plate assay를 통
Fig. 1. Time-course growth of S. cerevisiae CEN.PK2-1D, JHS100
and JHS200 strains in YP medium containing 20 g/L glucose
at 30
oC and 250 rpm. Symbols denote as follows; ▼ , CEN.PK2-
1D; ● , JHS100; ○ , JHS200.
해 비교해 보았다(Fig. 2A). 에탄올을 첨가하지 않은 YPD 배지에서는 세 균주 모두 동일한 colony 형성능력을 보였다.
10% 에탄올을 첨가한 경우, 실험실용 효모인 CEN.PK2-1D 는 급격하게 colony 형성능력이 떨어져 103으로 희석한 경 우, 작은 colony를 형성하였다. 한편 산업용 효모 JHS100과 JHS200은 104으로 희석한 경우까지 colony 형성능력을 보 였다. Fig. 2B와 같이 고농도 ethanol에서의 세포생존능력을 시험한 결과 중에서 배양 후 1시간의 세포생존능력과 비교 할 경우, CEN.PK2-1D 균주는 6시간 배양 후에 모든 세포 가 사멸하였으나 JHS100은 40%가 생존하였고 JHS200은 오히려 약 37% 증가하였다. JHS200은 12.5시간 이후에 모 두 사멸한 반면 JHS200은 22시간에 모두 사멸하였다. 세 균 주를 비교하였을 때 세포성장능력과 ethanol 내성, 효모균주 의 생존능력에서 모두 산업용균주가 실험실균주보다 우수하 였다. 특히 JHS100은 세포성장속도가, JHS200은 ethanol에 대한 내성이 우수한 것으로 판단되었다.
순수배지를 이용한 회분식 배양
세 균주의 기본적인 발효특성을 규명하기 위하여 순수배 지인 YP배지에 300 g/L glucose를 첨가한 배지를 이용하여 플라스크 수준에서 CEN.PK2-1D과 JHS100, JHS200의 회 분식 배양을 수행하였다(Fig. 3). 세 균주 JHS100, JHS200,
CEN.PK2-1D의 순서로 기질인 glucose를 빠르게 소모하였 고, 약 45시간 이후 JHS100만이 glucose를 모두 소모하였다.
Glucose의 빠른 소모에 따라서 JHS100 균주는 CEN.PK2-1D 균주와 비교하여 세포농도는 9%, ethanol 생산성(productivity) 은 23% 가량 높았다. 다만 ethanol 생산수율은 세 균주 모 두 0.46 g/g을 나타내었다. 이와 같은 결과를 분석하면, 세 효모의 비성장속도의 차이에 따라 발효속도가 일치하는 경
Fig. 2. Spot assay (A) and cell viability test (B) of S. cerevisiae CEN.PK2-1D, JHS100 and JHS200 strains. For spot assay, cell suspensions were diluted with YP medium by 10 times and spot- ted onto the solid YPD with or without 10% ethanol. After the cells were incubated in YPD medium with 15% ethanol, the col- ony forming unit (CFU) in the broth was determined.
Fig. 3. Batch fermentations of S. cerevisiae CEN.PK2-1D (A),
JHS100 (B) and JHS200 (C) strains using a baffled flask con-
taining YP medium with 300 g/L glucose at 30
oC, 90 rpm and
pH 5.5. Symbols denote as follows; ■ , glucose concentration; ● ,
O. D at 600 nm; ★ , ethanol concentration; ▲ , glycerol concentra-
tion; ▼ , acetate concentration.
향을 보이기 때문에 세포의 성장속도가 ethanol 생산수율 보 다 ethanol 생산성과 밀접한 연관이 있음을 보여주었다. 최 종적으로 300 g/L glucose를 함유한 YP배지를 이용한 경우, 가장 우수한 탄소대사능력을 보인 JHS100 균주는 회분식 배 양을 통해 136.6 g/L ethanol을 최종적으로 생산하였고, 3.05 g/L-hr의 생산성을 보였다(Table 1).
카사바 당화액을 이용한 회분식 배양
카사바 전분은 산 또는 효소를 이용하여 가수분해가 가능 한 unbranched amylose (약 20%)와 branched amylopectin (약 80%)으로 이루어져 있기 때문에 가수분해 시 대부분 미 생물이 이용할 수 있는 당으로 전환된다고 알려져 있다[3, 8]. 발효실험을 진행하기에 앞서 (주)창해에탄올로부터 제공 받은 카사바 당화액을 HPLC를 이용하여 분석하였다. 그 결 과 카사바 당화액에는 총 262.7 g/L의 당이 함유되어 있었고, 그 중에서 glucose가 약 93.6%를 차지하였으며 나머지는 이 당류(malose) 5.8%와 삼당류(malotriose) 0.6%로 이루어져 있었다. 카사바 당화액을 이용하고 질소원을 추가하지 않은 배지를 이용하고 실험실용 효모인 CEN.PK2-1D과 산업용 효모인 JHS100, JHS200를 사용하여 1 L 발효기 규모의 회 분식 혐기성 발효를 수행하였다(Fig. 4). YP 순수배지를 이 용한 것과 마찬가지로 JHS100 균주는 빠른 속도로 카사바 당화액에 함유된 250 g/L glucose를 약 30시간에 모두 소모 한 반면, JHS200 균주는 약 20 g/L glucose를, CEN.PK2-1D 균주는 약 70 g/L glucose를 소모하지 못하였다. 세포의 성 장도 glucose의 소모와 비례하여 JHS100 균주는 CEN.PK2- 1D 균주와 비교하여 1.7배의 높은 최종균체농도를 보였다.
Ethanol 생산에 있어서도, glucose 소모에 따라서 ethanol 생 산변수가 달라졌는데, JHS100 균주는 CEN.PK2-1D 균주와 비교하여 1.5배의 최종 ethanol 농도와 생산성 증가를 보였 다. YPD 순수배지를 이용한 회분식배양결과(Fig. 3)와 달리, ethanol 수율 또한 JHS100 균주는 CEN.PK2-1D 균주보다 13% 증가한 수치를 보였다. 최종적으로 카사바 당화액만을 배지로 이용하고 산업용 효모 JHS100를 균주로 이용한 혐 기성 회분식 배양을 통하여, 최종 ethanol 농도 106.1 g/L,
생산수율 0.423 g/g, 생산성 3.15 g/L-hr를 얻었다.
배지에 따른 실험실용 및 산업용 효모의 ethanol 발효성능 비교분석
순수배지인 YP배지는 다양한 유기질소원과 무기질이 포 함되어 있는 반면, 카사바 당화액은 질소원 등의 함유량이 제한적이다. 이러한 배지의 환경에서 실험실용 및 산업용 효 모의 ethanol 발효성능은 Fig. 3과 Fig. 4에서 보는 것과 같 이 다른 경향을 보였다. 혐기성 발효에서 얻은 최종발효변 수를 Table 1에 정리하였고, 이 결과를 이용하여 배지에 따 라서 본 연구에 사용한 효모의 특성을 비교하고자 한다. 실 험실용 효모인 CEN.PK2-1D는 세포성장 및 에탄올 생산관 련 발효변수에 대해서 카사바 당화액을 이용한 결과가 YP 배지를 이용한 결과보다 상당히 낮았는데, 예로 ethanol 생 산수율은 19%, 생산성은 17%가 감소하였다. 산업용 효모인 JHS100과 JHS200은 ethanol 생산수율은 8%가 감소한 반면, 생산성은 유사한 값을 보였다. 즉, 세 효모 모두 카사바 당 화액을 이용하여 발효를 할 경우 영양이 풍부한 순수 YP배 지에서 배양하였을 때에 비하여 ethanol 생산수율이 감소하 는 경향을 보였고, 특히 CEN.PK2-1D은 ethanol 생산성이 큰 폭으로 감소하여 실험실용 효모가 산업용 효모에 비하여 배지조건, 특히 질소원에 더 많은 영향을 받는 것을 확인할 수 있었다.
Ethanol 발효 시 세포의 성장속도 및 ethanol에 대한 내성 과 더불어 중요하게 여겨지는 또 다른 요인은 부산물인 glycerol과 acetate이다. 두 가지 모두 세포내의 redox balance 를 맞추기 위해 생성되는 부산물로서 glycerol은 ethanol 다 음으로 탄소를 많이 소모할 뿐만 아니라 공정상의 어려움을 야기하고, acetate는 세포성장을 저해하는 등 ethanol 생산에 부정적인 영향을 미치는 것으로 알려져 있다[5, 7, 9]. 따라 서 산업용 효모의 우수한 ethanol 발효성능에 대해 분석해 보고자 세 효모에서 생성된 부산물을 비교해 보았다(Table 1). Acetate는 두 배지 모두 CEN.PK2-1D에서 가장 많이 생 성되었고 그 수율 또한 가장 높았다. 다음으로 glycerol의 생 성 측면에서는 세 효모의 ethanol 수율이 비슷했던 YPD배
Table 1. Summarized results of anaerobic batch fermentations of S. cerevisiae CEN.PK2-1D, JHS100 and JHS200 strains in YPD medium and cassava hydrolysate.
Media Strain Maximum
O.D.
Ethanol Glycerol Acetate
Concentration (g/L)
Yield (g/g)
Productivity (g/L-hr)
Concentration (g/L)
Yield (g/g)
Concentration (g/L)
Yield (g/g) YP
+ 300 g/L glucose
CEN.PK2-1D 33.4 110.7 0.462 2.47 8.7 0.036 2.8 0.012
JHS100 36.4 136.6 0.461 3.05 10.9 0.037 2.3 0.008
JHS200 27.6 129.8 0.463 2.90 10.2 0.036 1.4 0.005
Cassava hydrolysate (250 g/L glucose)
CEN.PK2-1D 31.1 69.5 0.373 2.06 17.0 0.091 4.0 0.022
JHS100 52.1 106.2 0.423 3.15 14.4 0.057 1.3 0.005
JHS200 40.5 98.6 0.424 2.92 12.0 0.048 0.5 0.002
양에서는 수율이 비슷한 수준이었지만, 카사바 당화액에서 는 세 효모 모두 glycerol 수율이 증가하였으며 특히, YPD 배양에 비해 ethanol 수율이 가장 크게 감소했던 CEN.PK2- 1D의 경우 약 2.5배 증가하여 그 증가폭이 가장 컸다. 다시 말해 총질소 성분의 양이 제한적인 카사바 당화액에서는 CEN.PK2-1D보다 JHS100과 JHS200에서 부산물인 glycerol 과 acetate가 더 적게 생성된다고 할 수 있다.
본 연구에서 실시한 ethanol 발효실험의 결과 실험실용 효 모에 비하여 산업용 효모의 ethanol 발효성능이 우수하였고, 이러한 발효성능의 차이는 카사바 당화액에서 더 확연하게 나타남을 확인할 수 있었다. 이러한 현상은 산업용 효모인 JHS100과 JHS200이 실험실용 효모인 CEN.PK2-1D에 비하 여 세포성장속도가 빠르고 ethanol에 대한 내성과 생존능력 이 뛰어날 뿐만 아니라, 질소원에 대한 낮은 요구성과 부산 물인 glycerol과 acetate의 낮은 생산성 때문이라 예상한다.
따라서 무작위 돌연변이에 의해 제작된 산업용 효모인 JHS100과 JHS200은 이러한 특성에 관련된 대사경로에 변 화가 생겼을 것으로 추정되며, 차후에 이러한 결과를 바탕 으로 JHS100과 JHS200의 유전자 및 단백질 수준에서의 분 석을 진행할 것이다.
요 약
실험실용 효모 CEN.PK2-1D와 산업용 효모 JHS100와 JHS200의 ethanol 발효성능을 순수배지와 카사바 당화액에 서 비교하고 발효특성을 규명하기 위해 세포성장속도와 ethanol 내성, 부산물 glycerol과 acetate의 생성에 대해 비교 하였다. JHS100과 JHS200은 CEN.PK2-1D과 비교하여 세 포성장이 빨랐으며 ethanol에 대한 내성이 높았다. 순수YP 배지에 300 g/L glucose를 탄소원으로 이용한 회분식 배양 에서 세 효모 모두 0.46 g/g의 ethanol 생산수율을 나타냈으 며 생산성은 세포성장속도가 가장 빨랐던 JHS100이 3.05 g/
L-hr로 가장 높아서 최종적으로 JHS100 균주는 136.6 g/L의 ethanol을 생산하였다. 카사바 당화액에 질소원을 추가하지 않은 배지를 이용한 혐기성 회분식 배양에서 산업용 효모 JHS100은 106.1 g/L ethanol을 최종적으로 생산하였고, 0.42 g/g 생산수율과 3.15 g/L-hr 생산성을 보였다. 특히, 카사바 당화액과 순수 YP배지를 이용한 발효의 ethanol 생산변수를 비교할 경우, 실험실용 효모 CEN.PK2-1D는 생산수율과 생 산성이 각각 19%와 17% 감소한 반면, 산업용 효모 JHS100 과 JHS200은 생산수율이 8% 감소하였고 유사한 생산성을 보였다. 또한, ethanol 생산과정의 최대 부산물인 glycerol과 acetate의 생산에 대해서 JHS100과 JHS200이 CEN.PK2-1D 에 비하여 크게 낮았다. 따라서 산업용 효모인 JHS100과 JHS200의 뛰어난 ethanol 발효성능은 빠른 세포성장과 높은 ethanol 내성, 낮은 질소원 요구성, 부산물인 glycerol과 acetate의 낮은 생산성 등에 기인하는 것으로 예상한다.
A CKNOWLEDGEMENTS
We specially thank to JinRo Co. and Dr. Gi-Wook Choi at Changhae Ethanol Co. for providing two industrial strains and cassava hydrolysate, respectively. This work was funded by a grant from Marine Biotechnology Program funded by
Fig. 4. Batch fermentations of S. cerevisiae CEN.PK2-1D (A),
JHS100 (B) and JHS200 (C) strains using a bioreactor contain-
ing cassava hydrolysate only at 30
oC, 200 rpm and pH 5.5
without aeration. Symbols denote as follows; ■ , glucose concen-
tration; ● , O. D at 600 nm; ★ , ethanol concentration; ▲ , glycerol
concentration; ▼ , acetate concentration.
Ministry of Land, Transport and Maritime Affairs of Korean Government and a research development project no. FP0900- 2009-02 in Forest Science Technology supported by Korea Forest Service, Korea.
R EFERENCES
1. Bai, F. W., L. J. Chen, Z. Zhang, W. A. Anderson, and M.
Moo-Young. 2004. Continuous ethanol production and evaluation of yeast cell lysis and viability loss under very high gravity medium conditions. J. Biotechnol. 110: 287- 293.
2. Cheng, J.-S., B. Qiao, and Y.-J. Yuan. 2008. Comparative proteome analysis of robust Saccharomyces cerevisiae insights into industrial continuous and batch fermentation.
Appl. Microbiol. Biotechnol. 81: 327-338.
3. Ejiofor, A. O., Y. Chisti, and M. Moo-Young. 1996. Culture of Saccharomyces cerevisiae on hydrolyzed waste cassava starch for production of baking-quality yeast. Enzyme Microb. Tech. 18: 519-525.
4. Jansson, C., A. Westerbergh, J. Zhang, X. Hu, and C. Sun.
2009. Cassava, a potential biofuel crop in (the) People’s Republic of China. Appl. Energy 86: S95-S99.
5. Kim, J.-W., Y.-W. Chin, Y.-C. Park, and J.-H. Seo. 2012.
Effects of deletion of glycerol-3-phosphate dehydrogenase and glutamate dehydrogenase genes on glycerol and ethanol metabolism in recombinant Saccharomyces cerevisiae. Bio- process Biosys. Eng. 35: 49-54.
6. Lee, S.-H., T. Kodaki, Y.-C. Park, and J.-H. Seo. 2012.
Effects of NADH-preferring xylose reductase expression on
ethanol production from xylose in xylose-metabolizing recombinant Saccharomyces cerevisiae. J. Biotechnol. 158:
184-191.
7. Maiorella, B., H. W. Blanch, and C. R. Wilke. 1983. By- product inhibition effects on ethanolic fermentation by Saccharomyces cerevisiae. Biotechnol. Bioeng. 25: 103-121.
8. Moon, S.-K., S. W. Kim, and G.-W. Choi. 2012. Simultaneous saccharification and continuous fermentation of sludge- containing mash for bioethanol production by Saccharomyces cerevisiae CHFY0321. J. Biotechnol. 157: 584-589.
9. Pons, M.-N., A. Rajab, and J.-M. Engasser. 1986. Influence of acetate on growth kinetics and production control of Saccharomyces cerevisiae on glucose and ethanol. Appl.
Microbiol. Biotechnol. 24: 193-198.
10. Solomon, B. D., J. R. Barnes, and K. E. Halvorsen. 2007.
Grain and cellulosic ethanol: History, economics, and energy policy. Biomass Bioenerg. 31: 416-425.
11. Sriroth, K., K. Piyachomkwan, S. Wanlapatit, and S.
Nivitchan-zyong. 2010. The promise of a technology revolu- tion in cassava bioethanol: From Thai practice to the world practice. Fuel 89: 1333-1338.
12. Thatipamala, R., S. Rohani, and G. A. Hill. 1992. Effects of high product and substrate inhibitions on the kinetics and biomass and product yields during ethanol batch fermenta- tion. Biotechnol. Bioeng. 40: 289-297.
13. Zaldivar, J. Z., A. B. Borges, B. J. Johansson, H. S. Smits, S. V.-B. Villas-Bôas, J. N. Nielsen, and L. O. Olsson. 2002.
Fermentation performance and intracellular metabolite patterns in laboratory and industrial xylose-fermenting Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 59: 436-442.