Vol. 24, No. 1, 21-32, April 2013 Original Article
Received on April 10, 2013. Revised on April 19, 2013. Accepted on April 21, 2013 Correspondence to: Jae-Hun Cheong
Department of Molecular Biology, College of Natural Sciences, Pusan National University, Busandaehak-ro 63beon-gil, Geumjeong-gu, Busan 609-735, Korea
Tel: 82-51-510-2277, Fax: 82-51-513-9258, E-mail: [email protected] Co-correspondence to: Hyung-Hoi Kim
Department of Laboratory Medicine, School of Medicine, Pusan National University, 179 Gudeok-ro, Seo-gu, Busan 602-739, Korea Tel: 82-51-240-7419, Fax: 82-51-247-6560, E-mail: [email protected]
단핵구에서 TNF-α Gene의 전사 활성을 증가시키는 HIF-1과 NF-κB의 기능적 상호작용
박민정
1ㆍ이선민
2ㆍ옥순정
2ㆍ김혜림
2ㆍ김형회
2ㆍ정재훈
1
부산대학교 자연과학대학 분자생물학과1, 부산대학교 의과대학 진단검사의학교실2
Functional Interaction of HIF-1 and NF-κB Increasing the Transcriptional Activation of TNF-α Gene in Monocytes
Min-Jeong Park
1, Sun-Min Lee
2, Soon-Jung Ok
2, Hye-Rim Kim
2, Hyung-Hoi Kim
2, Jae-Hun Cheong
1
Department of Molecular Biology, College of Natural Sciences1, Department of Laboratory Medicine, School of Medicine2, Pusan National
University, Busan, Korea
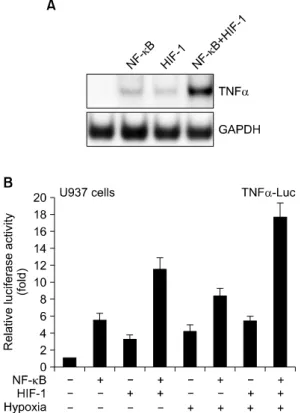
Background: Tumor necrosis factor alpha (TNF-α) is a pleiotropic cytokine fulfilling a broad variety of immunoregulatory functions. Monocytes and macrophages play a pivotal role in inflammation and immune regulation. NF-κB and HIF-1 are known to increase expression of the TNF-α gene in a separate way.
Methods: Human monocytic leukemia, U937 cells, were transfected using the standard electroporation method for intracellular expression of NF-κB and HIF-1. We performed analysis using the mammalian two-hybrid assay and co-immunoprecipitation assay for detection of protein interaction of both proteins. In addition, chromatin immuno- precipitation analysis was performed for examination of NF-κB and HIF-1 binding on the TNF-α gene promoter.
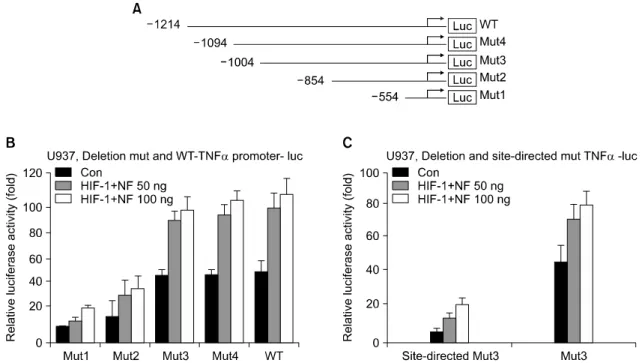
Results: Here we show that NF-κB and HIF-1 cooperatively induced an increase in expression of the TNF-α gene dependent on promoter activity by the direct protein interaction of these two transcription factors. Hypoxia signaling induced marked enhancement of the transactivation of TNF-α promoter by HIF-1 and NF-κB. A tandem NF-κB/HIF-1 binding site was identified within the TNF-α promoter, which acted as a strong enhancer element.
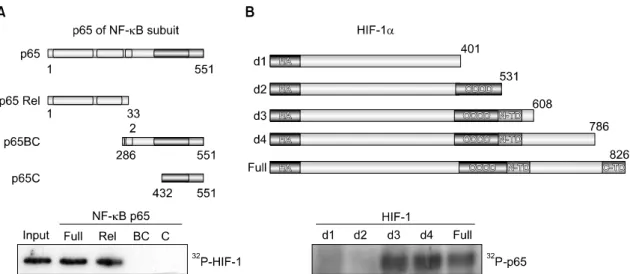
Physical association of the Rel domain of NF-κB and the N-TD domain of HIF-1 was required. Hypoxia treatment also resulted in a significant increase in the protein interaction of NF-κB and HIF-1 in vivo. Both transcription factors were recruited on the chromatin TNF-α promoter dependent on hypoxia stimuli.
Conclusion: The results of this study indicate that a variety of extracellular signals for activation of TNF-α gene expression might converge on the transcriptional regulation through the NF-κB/HIF-1 signaling pathway. (Korean J Blood Transfus 2013;24:21-32)
Key words: HIF-1, NF-κB, Hypoxia, TNF-α, Transactivation