Effects of Sodium Butyrate, a Histone Deacetylase Inhibitor, on TRAIL-mediated Apoptosis in Human Bladder Cancer Cells
Min-Ho Han
1and Yung Hyun Choi
2,3*
1Natural products Research Team, National Marine Biodiversity Institute of Korea, Seocheon-gun 325-902, Korea
2Department of Biochemistry, Dongeui University College of Korean Medicine, Busan 614-052, Korea
3Anti-Aging Research Center and Blue-Bio Industry RIC, Dongeui University, Busan 614-71, Korea Received October 26, 2015 /Revised November 12, 2015 /Accepted November 13, 2015
The tumour necrosis factor (TNF)-related apoptosis-inducing ligand (TRAIL) is considered a promising anticancer agent due to its unique ability to induce cancer cell death having only negligible effects on normal cells. However, many cancer cells tend to be resistant to TRAIL. In this study, we inves- tigated the effects and molecular mechanisms of sodium butyrate (SB), a histone deacetylase inhibitor, in sensitizing TRAIL-induced apoptosis in 5637 human bladder cancer cells. Our results indicated that co-treatment with SB and TRAIL significantly increased the apoptosis induction, compared with treat- ment with either agent alone. Co-treatment with SB and TRAIL effectively increased the cell-surface expression of death receptor (DR) 5, but not DR4, which was associated with the inhibition of cellular Fas-associated death domain (FADD)-like interleukin-1β-converting enzyme (FLICE) inhibitory protein (c-FLIP). Furthermore, the activation of caspases (caspase-3, -8 and -9) and degradation of poly(ADP- ribose) were markedly increased in 5637 cells co-treated with SB and TRAIL; however, the synergistic effect was perfectly attenuated by caspase inhibitors. We also found that combined treatment with SB and TRAIL effectively induced the expression of pro-apoptotic Bax, cytosolic cytochrome c and cleave Bid to truncated Bid (tBid), along with down-regulation of anti-apoptotic Bcl-xL expression. These re- sults collectively suggest that a combined regimen of SB plus TRAIL may offer an effective therapeutic strategy for safely and selectively treating TRAIL-resistant bladder cancer cells.
Key words :
Apoptosis, Caspase, DR5, Sodium butyrate, TRAIL ((TNF)-related apoptosis-inducing ligand)
*Corresponding author
*Tel : +82-51-850-7413, Fax : +82-51-853-4036
*E-mail : [email protected]
This is an Open-Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Life Science 2016 Vol. 26. No. 4. 431~438 DOI : http://dx.doi.org/10.5352/JLS.2016.26.4.431
서 론
항암제 개발의 주요한 전략 중 하나가 정상세포에는 영향을 주지 않으면서 암세포 특이적으로 증식 억제를 통한 체내에서 의 제거이다. 프로그램화된 세포사멸로 알려진 apoptosis가 개체의 발생과 증식 및 항상성 유지에 필수적인 과정이지만, 체내의 비정상적인 세포들을 제거하는 중요한 생리학적 과정 이기도 하다[13, 20]. 최근 apoptosis의 유도 기전에 대한 다양 한 분류가 시도되고 있으나, 일반적으로 세포막에 존재하는 death receptor (DR)와 해당 DR 특이적 death legend와의 결 합으로 개시되는 DR-mediated extrinsic apoptosis 경로와 mi- tochondria를 중심으로 apoptosis가 활성화되는 mitochon- dria-mediated intrinsic apoptosis 경로로 대별된다[4, 21].
한편 tumour necrosis factor (TNF) family에 속하는
TNF-related apoptosis-inducing ligand (TRAIL)는 정상세포 에는 유의적인 영향을 주지 않으면서 암세포 특이적인 apop- tosis 유도 능력으로 인하여 항암제로서의 적용에 매우 큰 장 점을 지닌다[1, 37]. TRAIL은 transmembrane type-I receptor 인 DR4 (TRAIL-R1) 및 DR5 (TRAIL-R2)와 결합으로 Fas-asso- ciated death domain protein (FADD)과 pro-caspase-8을 포함 하는 death-inducing signaling complex (DISC)를 형성하여 apoptosis 신호를 개시한다. 일단 DISC가 형성이 되고 나면, pro-caspase-8이 활성형으로 전환되면서 effector caspase인 caspase-3 및 -7이 연속적으로 활성화는 DR-mediated ex- trinsic apoptosis 경로인 caspase cascade가 형성된다[22, 38].
이 과정은 anti-apoptotic 유전자로 잘 알려진 경쟁적 저해제인
FADD-like apoptosis regulator (c-FLIP)에 의하여 조절된다
[3, 33]. 활성화된 caspase-8은 Bid를 truncated Bid (tBid) 형으
로 전환시켜 apoptosis 활성 신호를 미토콘드리아로 전달하여
cytochrome c와 같은 apoptosis 촉발인자들은 세포질로 유리
시켜 caspase-9가 활성화되는 mitochondria-mediated in-
trinsic apoptosis 경로를 증폭시킨다[4, 22]. 그러나 비록 TRAIL
이 암세포 선택적인 항암활성을 지니는 것은 확실하지만, 많
은 암세포에서 이미 TRAIL에 대한 저항성을 획득되어 있음이
알려지면서, TRAIL의 저항성을 극복할 수 있는 신물질의 발 굴에 큰 관심이 모아지고 있다[6, 11, 34]. 아울러 다양한 천연 물 또는 생체 신호 분자 조절제들이 TRAIL 저항성을 가지는 다양한 암세포의 사멸을 촉진할 수 있다는 결과들이 발표되고 있다[6, 12, 17-19, 22, 27, 30, 31, 35, 38]. 이러한 결과들은 TRAIL 저항성의 극복을 위한 새로운 항암 전략 수립의 근거 로 제시될 수 있다.
한편 histone의 lysine 잔기에 acetylation과 deacetylation 은 histone deacetylases (HDACs)와 histone acetyltransfer- ases (HATs)의 두 효소에 의하여 전형적으로 조절되며, chro- matin 구조의 변형과 유전자 조절의 개시에 기본적으로 관여 한다[2, 25]. 특히 HAT의 기능적 불활성 또는 HDAC 활성의 조절 장애에 따른 특정 유전자의 비정상적인 발현은 암의 개 시와 종양 세포의 증식에 직접적으로 원인이 되기 때문에 HDAC 저해제는 항암 약물로서의 개발 가능성이 높음을 알 수 있다[2, 29]. 또한 최근 보고에 의하면 HDAC 저해제들은 다양한 TRAIL 저항성 암세포에서 apoptosis 유도에 효과적으 로 작용할 수 있음이 알려져 TRAIL 저항성 극복을 위한 복합 약물로서의 활용 가능성이 높음을 알 수 있다[5, 9, 12, 36].
본 연구에서는 이러한 가능성에 대한 추가적인 자료의 제시를 위하여 방광암세포를 대상으로 대표적인 HDAC 저해제인 so- dium butyrate (SB)의 활용 가능성을 조사하였으며, 유의적인 결과를 얻었기에 이를 보고하고자 한다.
재료 및 방법
시약 및 항체
SB와 재조합 TRAIL은 Calbiochem (San Diego, CA, USA) 과 KOMA Biotech Inc. (Seoul, Republic of Korea)에서 구입하 였으며, dimethyl sulfoxide (DMSO, Sigma-Aldrich Chemical Co., St. Louis, MO, USA)에 녹인 후 적정 농도로 희석하여 사용하였다. Dulbecco’ modified Eagle’s medium (DMEM), fetal bovine serum (FBS), penicillin 및 streptomycin은 GIBCO- BRL (Gaithersburg, MD, USA)에서 구입하였다. 3-(4,5-di- methyl-2-thiazolyl)-2,5-diphnyl-2H-tetrazolium bromide (MTT) 와 propidium iodide (PI)는 Sigma-Aldrich Chemical Co.에서 구입하였으며, annexin V-fluorescein isothiocyanate (FITC)와 caspase 억제제(pan-caspase inhibitor, z-Val-Ala-Asp (VAD)- fluoromethyl ketone (fmk), caspase-9 inhibitor, z-Leu-Glu- His-Asp (LEHD)-fmk, caspase-8 inhibitor, z-Ile-Glu-Thr-Asp (IETD)-fmk 및 caspase-3 inhibitor, z-Asp-Glu-Val-Asp (DEVD)- fmk)는 Calbiochem에서 구입하였다. 5,5′,6,6′-tetrachloro-1, 1′,3,3′-tetraethylbenzimidazolylcarbocyanine iodide (JC- 1)와 caspase activity assay kit는 R&D Systems (Minneapolis, MN, USA) 제품을 사용하였으며, 본 연구에 사용된 일차 항체 는 Santa Cruz Biotechnology (Santa Cruz, CA, USA) 및
Chemicon (Temecula, CA, USA)에서 구입하였다. Peroxidase- labelled donkey anti-rabbit, sheep anti-mouse immun- oglobulin 및 enhanced chemiluminescence (ECL) kit는 Amersham (Arlington Heights, IL, USA) 제품을 사용하였으 며, 나머지 언급되지 않은 시약들은 Sigma-Aldrich Chemical Co.에서 구입하였다.
세포배양 및 MTT assay
5637 인체 방광암세포는 American Type Culture Collec- tion (Manassas, MD, USA)에서 구입하였으며 10% FBS, 100 U/ml penicillin 및 100 μg/ml streptomycin이 함유된 DMEM 배지를 이용하여 37°C, 5% CO
2조건하에서 배양하였다. 5637 세포의 증식에 미치는 SB과 TRAIL의 영향을 조사하기 위해서 는 미토콘드리아 효소에 의하여 MTT-formazan을 형성하는 원리에 기반을 둔 MTT assay를 이용하였다.
DNA flow cytometry 분석
Apoptosis의 정량적 분석을 위한 DNA flow cytometry 분 석을 위해 준비된 세포를 고정하고 염색하기 위하여 Cycle TEST PLUS DNA REAGENT Kit (Becton Dickinson, San Jose, CA, USA)를 이용하였으며, 염색 후 4
oC, 암실에서 30분 동안 반응을 시켰다. 반응이 끝난 세포를 35 mm mesh를 이용 하여 단일세포로 분리한 후 FACSCalibur (Becton Dickinson) 를 적용시켜 형광반응에 따른 Cellular DNA content 및 histo- gram을 CellQuest software 및 ModiFit LT 프로그램을 이용 하여 분석하였다.
단백질의 분리 및 Western blot analysis
준비된 세포를 lysis buffer [25 mM Tris-Cl (pH 7.5), 250
mM NaCl, 5 mM ethylenediaminetetra acetic acid (EDTA),
1% nonidet P40 (NP-40), 1 mM phenymethylsulfonyl fluo-
ride 및 5 mM dithiothreitol (DTT)]를 이용하여 4
oC에서 1시
간 동안 반응시켜 용해시켰다. 동일 조건에서 배양된 세포의
세포질 cytochrome c 발현 검출을 위한 세포질 단백질은
Active Motif Co. (Carlsbad, CA, USA)에서 구입한 cytosolic
fractionation kit를 시용하여 분리하였다. Bio-Rad 단백질 정
량 시약(Bio-Rad, Hercules, CA, USA)을 이용하여 단백질 농
도를 정량 한 다음 Laemmli sample buffer (Bio-Rad)와 혼합
하여 균일한 농도의 단백질 sample을 만들었다. 이렇게 만들
어진 단백질 sample은 sodium dodecyl sulphate (SDS)-polya-
crylamide gel을 이용한 전기영동을 실시하고 nitrocellulose
membrane (Schleicher and Schuell, Keene, NH, USA)으로
electroblotting에 의해 전이시켰다. 단백질이 전이된 nitro-
cellulose membrane은 5% skim milk 용액을 이용하여 비특이
적인 단백질들에 대한 blocking을 실시하였다. 다시 1차 항체
를 처리하고, 적정 2차 항체를 사용하여 반응시켰다. 2차 항체
Fig. 1. Effects of SB on the inhibition of cell viability and induction of apoptosis in TRAIL-treated 5637 cells. Cells were seeded at 1×105 cells/ml, and were then treated with the indicated concentrations of TRAIL, SB, or TRAIL plus SB for 24 hr. (A) Cell viability was determined by an MTT assay. (B) The cells were stained with annexin V-FITC and PI, and the percentages of apoptotic cells (annexin V+ cells) were then analyzed using flow cytometric analysis. The data are expressed as the mean
± SD of three independent experiments. The significance was determined by the Student’s t-test (*p<0.05 vs. untreated control).
에 대한 반응이 끝난 후 충분히 세척하고 암실에서 ECL 용액 을 적용시킨 다음 X-ray film에 감광시켜 특정단백질의 발현 양을 분석하였다.
DRs의 세포 표면 부착 분석
DR4 및 DR5의 세포 표면 발현 여부를 조사하기 위하여 준비된 세포에 DR4 및 DR5 항체를 1시간 동안 반응시킨 후 phycoerythrin (PE)-conjugated goat anti-mouse IgG (Santa Cruz Biotechnology)를 1시간 처리하였다. 단일세포로 분리한 후 DRs의 발현 양상은 FACSCalibur를 이용하여 조사하였다[23].
Mitochondrial membrane potential (MMP, Δψm)의 분석
SB와 TRAIL 처리에 의한 apoptosis 유발에서 미토콘드리 아의 관련성 여부를 확인하기 위하여 준비된 세포를 모은 다 음 500 μl의 PBS에 부유시키고 10 μM의 JC-1 용액을 첨가하여 37
oC에서 20분 동안 반응시켰다. 반응이 끝난 후 원심분리를 이용하여 상층액을 제거한 다음 차가운 PBS를 첨가하고 FACSCalibur에 적용시켜 MMP (Δψm)의 변화 정도를 분석하 였다.
In vitro caspase의 활성 측정
준비된 세포들의 caspase 활성을 측정하기 위하여 150 μg의 단백질이 함유된 50 μl의 sample에 50 μl의 reaction buffer [40 mM HEPES (pH 7.4), 20% glycerol (v/v), 1 mM EDTA, 0.2% NP-40 및 10 mM DTT]를 혼합한 다음 5 μl caspases 기질[caspase-3, Asp-Glu-Val-Asp (DEVD)-p-nitroaniline (pNA);
caspase-8, Ile-Glu-Thr-Asp (IETD)-pNA; caspase-9, Leu-Glu-
His-Asp (LEHD)-pNA]을 첨가하여 37
oC, 암실에서 3시간 동 안 반응시켰다. 반응이 끝난 후 ELISA reader (Molecular Devices, Sunnyvale, CA, USA)를 이용하여 405 nm 파장에서 각 caspase 활성 정도에 따른 변화를 측정하였다.
통계분석
모든 실험결과는 평균 ± 표준편차로 표시하였고 SigmaPlot (Systat Software Inc., San Jose, CA, USA)을 이용하여 Student
t-test를 이용하여 통계적 유의성을 얻었다.결과 및 고찰
TRAIL에 의한 세포증식 억제 및 apoptosis 유발에 미치 는 SB의 영향
5637 인체 방광암세포에서 SB에 의한 TRAIL의 암세포 증 식 억제 감수성 증대 가능성을 조사하기 위하여 SB와 TRAIL 단독 및 복합 처리 24시간 후 MTT assay를 실시하였다. Fig.
1A의 결과에 나타내었듯이, TRAIL 단독 처리에 의한 5637
세포의 증식억제가 SB와의 혼합처리에 의하여 유의적으로 증
대되었음을 알 수 있었다. 이런 증식 억제 감수성의 증대가
apoptosis 유발 촉진과 연관성이 있을 것으로 기대되어 동일
조건에서 배양된 5637 세포를 대상으로 flow cytometry 분석
을 실시한 결과, TRAIL 단독 처리군에 의하여 유발된 apopto-
sis가 SB 동시 처리에 의하여 매우 증가되었다(Fig. 1B). 이러한
현상은 TRAIL 저항성을 지니는 대장암세포와 백혈병세포, 유
방암세포, 폐암 및 전립선 암세포에서 관찰된 것과 유사한 현
상이었다[16, 24, 33, 36]. 특히 TRAIL에 대한 유사한 저항성을
가진 것으로 알려진 T24 방광암세포에서도 본 연구의 결과와
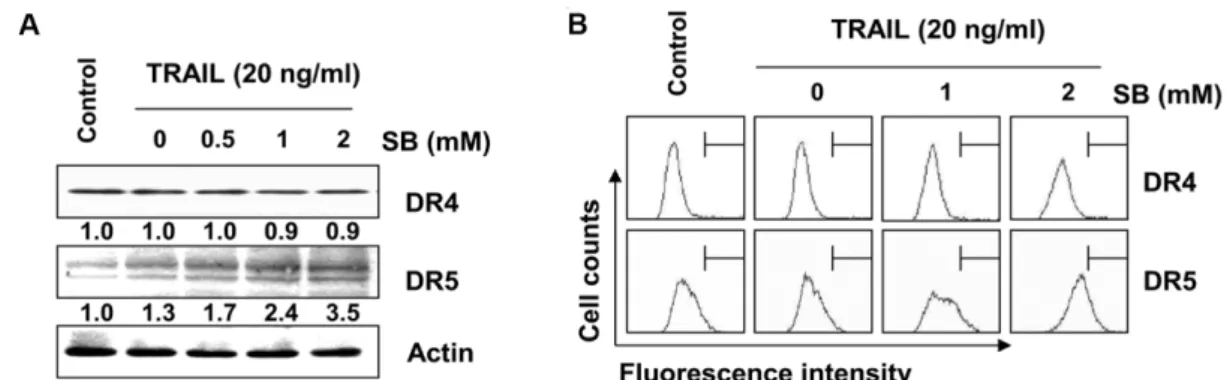
Fig. 2. Effects of SB and TRAIL on the expression of DRs in 5637 cells. The cell were co-treated with indicated concentrations of TRAIL and SB for 24 hr. (A) Cells were lysed and then equal amounts of cell lysates were separated on SDS-polyacrylamide gels and transferred to nitrocellulose membranes. Membranes were probed with anti-DR4 and anti-DR5 antibodies. An ECL detection system was used for visualization of proteins. Actin was used as an internal control. The numbers represent the average densitometric analyses as compared with actin in, at a minimum, two or three different experiments. (B) The cell surface expression of DR4 and DR5 of cells grown under the same conditions as in (A) was analyzed by flow cytometry after staining with anti-DR4 and anti-DR5 antibodies.
같은 효과가 보고된 바 있어[8], 방광암세포에서 SB가 TRAIL 에 의한 apoptosis 유도를 증진시킬 수 있음을 확인하였다.
DR 연관 apoptosis 조절인자들의 발현에 미치는 SB와 TRAIL의 영향
TRAIL에 의한 apoptosis 유발 신호의 개시는 일반적으로 extrinsic pathway를 경유하며, TRAIL은 다양한 DRs 중에서 DR4 또는 DR5와의 결합을 통하여 apoptosis 신호를 촉진한다 [21, 28]. 따라서 5637 세포에서 SB와 TRAIL의 복합처리에 의 한 apoptosis 신호에 미치는 DRs의 영향을 조사한 결과, TRAIL 처리군에서 DR5 단백질의 발현이 SB 농도 의존적으로 증가하 였을 뿐만 아니라(Fig. 2A), 세포 막 표면으로의 전이가 증가되 었음을 알 수 있었다(Fig. 2B). 그러나, DR4의 경우는 SB와 TRAIL 복합처리군에서 큰 변화가 관찰되지 않았다. 그리고 SB 및 TRAIL 복합처리에 의하여 Fas, Fas Ligend (FasL), TNF receptor-associated protein with a death domain (FADD)의 발현에는 큰 변화가 없었으나, cFLIP의 발현은 복합 처리 시간 의 경과에 따라 매우 감소되었다(Fig. 3). 이러한 결과는 Fas/
FasL 및 TNF signaling 경로는 SB 및 TRAIL 복합처리에 의하 여 큰 영향을 받지 않으며, cFLIP의 발현 감소에 따른 DR- mediated extrinsic apoptosis 경로가 활성화되었음을 의미한 다. 따라서 SB 및 TRAIL 복합처리는 DR5의 발현 증대를 통하 여 DISC의 형성을 촉진함으로서[3, 32] extrinsic apoptosis 경 로의 개시를 유발하였을 것으로 생각된다.
Inhibitor of apoptosis protein (IAP) family 단백질 및 caspase의 활성에 미치는 SB와 TRAIL의 영향
TRAIL과 연관된 extrinsic apoptosis 경로 활성과 직접 연관 이 있는 caspase는 caspase-8로서 caspase-8이 활성화되면 ef- fector caspase에 해당되는 caspase-3 또는 -7의 효소적 활성의
Fig. 3. Effects of SB and TRAIL on the levels of DR-related and IAP family proteins in 5637 cells. Cells were treated with 2 mM SB and 20 ng/ml TRAIL alone, or a combination of SB and TRAIL for 12 or 24 hr and harvested. Cells were lysed and then equal amounts of cell lysates were separated on SDS-polyacrylamide gels and transferred to nitrocellulose membranes. Membranes were probed with the indicated antibodies. An ECL detection system was used for visualization of proteins. Actin was used as an internal control. The numbers represent the aver- age densitometric analyses as compared with actin in, at a minimum, two or three different experiments.
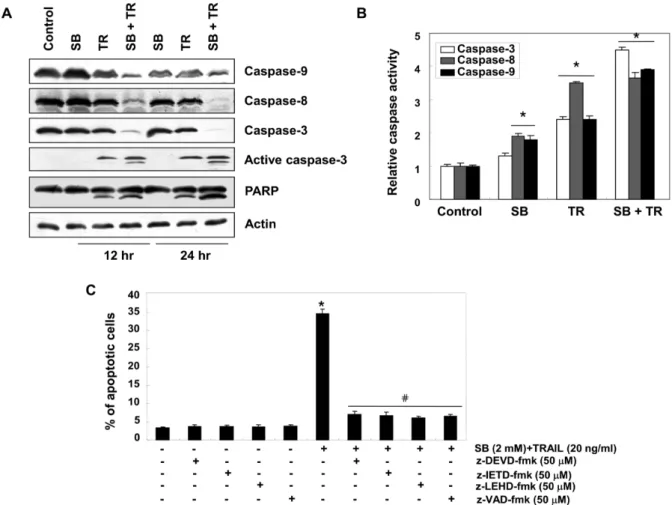
Fig. 4. Effects of SB and TRAIL on the activation of caspases in 5637 cells. Cells were treated with 2 mM SB and 20 ng/ml TRAIL alone, or a combination of SB and TRAIL for 12 or 24 hr. (A) Western blotting was performed using the indicated antibodies and anti-actin antibody to serve as control for the loading of protein level. (B) Cells were treated with 2 mM SB and 20 ng/ml TRAIL alone, or a combination of SB and TRAIL for 24 hr and caspase (-3, -8, and -9) activity was determined using caspase assay kits. Data are expressed as the mean ± SD of three independent experiments. The significance was determined using Student's t-test (*p<0.05 vs. untreated control). (C) Cells were incubated in the presence or absence of caspase inhibitors for 1 h before being exposed to 2 mM SB and 20 ng/ml TRAIL. After 24 hr of incubation, the percentages of apoptotic cells (annexin V+ cells) were then analyzed using flow cytometric analysis. The data are expressed as the mean
± SD of three independent experiments. The significance was determined by the Student’s t-test (*p<0.05 vs. untreated control;
#p<0.05 vs SB plus TRAIL-treated cells).
증대로 다양한 기질단백질들의 분해가 개시된다[21, 28]. 한편 caspase의 활성은 IAP family에 속하는 단백질들과의 직접적 인 결합을 통하여 그들의 apoptotic 활성을 억제할 수 있는 것으로 알려져 있다[14, 15]. 본 연구의 결과에 의하면 SB와 TRAIL이 함께 처리된 5637 세포에서 IAP family에 해당되는 단백질들(XIAP, cIAP-1 및 cIAP-2)의 발현이 매우 감소되었으 며(Fig. 3), extrinsic 및 intrinsic apoptosis 경로의 initiator cas- pase로 알려진 caspase-8 및 -9의 비활성형 단백질의 발현이 매우 감소되었다. 아울러 두 caspase의 하위 effector caspase 인 caspase-3의 비활성형 단백질의 감소와 활성형 단백질의 증가가 관찰되었다(Fig. 4A). 따라서 각 caspase의 활성화 정도 를 각 효소들의 기질을 이용한 정량적인 비교를 실시한 결과,
SB 및 TRAIL 단독 처리군에 비하여 복합 처리군에서 3가지
caspase 활성이 모두 유의적으로 증가되었다(Fig. 4B). 또한
DNA repair와 genomic stability에 관여하는 효소로서 활성화
된 caspase-3의 표적 단백질로 잘 알려져 있는 poly(ADP-ri-
bose) polymerase (PARP)의 단편화 유도 현상[26]이 TRAIL
단독 처리군에 비하여 SB와의 복합처리에 의하여 더욱 증가되
었다(Fig. 4A). 그러나 SB 및 TRAIL 복합 처리에 의한 5637
방광암세포의 apoptosis는 pan-caspase 저해제뿐 만 아니라,
각 caspase의 특이적 활성 저해제(z-DEVD-fmk, a caspase-3
inhibitor; z-IETD-fmk, a caspase-8 inhibitor; z-LEHD-fmk, a
caspase-9 inhibitor; z-VAD-fmk, a pan-caspase inhibitor) 처
리에 의하여 유의적으로 차단되었다(Fig. 4C). 이상의 결과는
Fig. 5. Effects of SB and TRAIL on the levels of Bcl-2 family proteins and values of MMP (Δ
ψ
m) in 5637 cells. (A) Cells were treated with 2 mM SB and 20 ng/ml TRAIL alone, or a combination of SB and TRAIL for 12 or 24 hr. Equal amounts of cell lysates or cytosolic proteins (for cytochrome c) were resolved by SDS-polyacrylamide gels, transferred to nitrocellulose membranes, and probed with the indicated antibodies. Actin was used as an internal control. The numbers represent the average densitometric analyses as compared with actin in, at a minimum, two or three different experiments. (B) Cells were treated with 2 mM SB and 20 ng/ml TRAIL alone, or a combination of SB and TRAIL for 24 hr. The values of MMP (Δψ
m) were evaluated by flow cytometric analysis. Data are expressed as the mean ± SD of three independent experiments.The significance was determined using Student's t-test (*p<0.05 vs. untreated control).
SB와 TRAIL의 복합 처리에 의한 extrinsic apoptosis 경로 활 성이 caspase cascade를 통하여 intrinsic apoptosis 경로를 증 폭시켰을 가능성이 있음을 의미하는 것으로 최종적인 apopto- sis 유도에 caspase 활성이 핵심적으로 관여하였음을 의미하 는 것이다.
SB와 TRAIL 동시 처리에 의한 apoptosis 유도에서 Bcl- 2 family 단백질의 역할
Bcl-2 family는 apoptosis 경로 유도를 촉진하거나 억제하는 정반대의 역할을 하는 다양한 단백질로 구성되어 있다. 미토 콘드리아는 apoptosis의 intrinsic pathway에서 가장 중요한 역할을 하는 기관으로 Bcl-2 family에 속하는 인자들의 발현 변화에 의하여 미토콘드리아 막의 전위가 조절된다고 알려져 있다[7]. Bcl-2 family는 apoptosis를 억제하는 anti-apoptotic 인자(Bcl-2 및 Bcl-xL 등)와 apoptosis를 유발하는 pro-apop- totic 인자(Bax 및 Bad 등)로 구성되어 있으며 이들은 서로 dimer의 형태로 결합을 할 수 있다[10]. 하지만 세포 내 스트레 스나 세포 독성에 의해 mitochondria의 막 전위가 변화되면, 이 균형은 깨어지고 mitochondria에서 cytochrome c와 같은 apoptosis 유발 인자들이 세포질로 방출된다. 방출된 cyto- chrome c는 apoptotic protease activating factor (Apaf)-1 과 caspase-9에 결합함으로써 apoptosome을 형성하여 caspase-9 를 활성화시키며, 결과적으로 caspase-3을 활성화시킴으로써 apoptosis를 유발시키는 것으로 알려져 있다[7]. 본 연구의 결
과에서, SB와 TRAIL의 복합 처리는 caspase-8 뿐만 아니라 caspase-9의 활성도 증대시켰기에(Fig. 4), 활성화된 caspase-9 가 미토콘드리아의 기능 손상과 연계된 intrinsic apoptosis 경 로를 활성화시켰을 것으로 기대되어, Bcl-2 family에 속하는 다양한 단백질들의 발현에 미치는 SB와 TRAIL의 영향을 조사 하였다. Fig. 5A에 나타낸 결과에 의하면, SB와 TRAIL의 복합 처리는 SB 및 TRAIL의 단독 처리군 비하여 Bax의 발현을 증 가시켰으며, Bcl-2의 발현에는 큰 변화가 없었으나 Bcl-xL의 발현은 감소시켰다. 특히 활성화된 caspase-8에 의한 trun- cated Bid (tBid)로 전환되어 intrinsic apoptosis 경로를 증폭시 키는 것으로 알려진 Bid 단백질[4, 21]의 tBid로의 단편화가 동반되었다. 또한 세포질에서의 cytochrome c의 발현이 SB 및 TRAIL의 단독 처리군에 비하여 복합 처리군에서 처리 시 간 의존적으로 증가되었다(Fig. 5A). 따라서 SB 및 TRAIL의 복합처리에 의한 intrinsic apoptosis 경로 활성의 부가적인 자 료를 제시하기 위하여 JC-1 fluorescent cationic dye를 이용한 MMP (Δψm) 값의 변화를 조사한 결과, 제시된 결과(Fig. 5B) 에서 알 수 있듯이 SB와 TRAIL의 단독 처리군에 비하여 복합 처리군에서 MMP (Δψm)의 소실이 현저하게 증대되었다.
이상의 결과는 SB와 TRAIL의 복합 처리에 의하여 DR5의
발현 증대에 따른 extrinsic apoptosis 경로의 개시가 tBid의
형성으로 intrinsic apoptosis 경로를 증폭시킴으로서 SB 및
TRAIL의 단독 처리군에 비하여 apoptosis 유발 효율을 증대
시켰음을 의미한다. 아울러 두 경로의 증폭은 Bid 의존적으로
일어나고 있음을 보여주는 결과이다. 이상의 결과에서 HDAC 저해제인 SB는 TRAIL 저항성을 획득한 암세포의 apoptosis 유도를 위한 촉진제로서 효율적으로 사용될 수 있음을 보여 주는 것이다.
감사의 글
이 논문은 2015년도 정부(미래창조과학부)의 재원으로 한 국연구재단의 지원을 받아 수행된 기초연구사업임(2015R1A 2A2A01004633).
References
1. Abdulghani, J. and El-Deiry, W. S. 2010. TRAIL receptor signaling and therapeutics. Expert. Opin. Ther. Targets 14, 1091-1108.
2. Barneda-Zahonero, B. and Parra, M. 2012. Histone deacety- lases and cancer. Mol. Oncol. 6, 579-589.
3. Benayoun, B., Baghdiguian, S., Lajmanovich, A., Bartoli, M., Daniele, N., Gicquel, E., Bourg, N., Raynaud, F., Pasquier, M. A., Suel, L., Lochmuller, H., Lefranc, G. and Richard, I. 2008. NF-kappaB-dependent expression of the anti- apoptotic factor c-FLIP is regulated by calpain 3, the protein involved in limb-girdle muscular dystrophy type 2A.
FASEB J. 22, 1521-1529.
4. Billen, L. P., Shamas-Din, A. and Andrews, D. W. 2008. Bid:
a Bax-like BH3 protein. Oncogene 27, S93-104.
5. Carew, J. S., Giles, F. J. and Nawrocki, S. T. 2008. Histone deacetylase inhibitors: mechanisms of cell death and prom- ise in combination cancer therapy. Cancer Lett. 269, 7-17.
6. Crowder, R. N. and El-Deiry, W. S. 2012. Caspase-8 regu- lation of TRAIL-mediated cell death. Exp. Oncol. 34, 160-164.
7. Donovan, M. and Cotter, T. G. 2004. Control of mitochon- drial integrity by Bcl-2 family members and caspase-in- dependent cell death. Biochim. Biophys. Acta 1644, 133-147.
8. Earel, J. K. Jr., VanOosten, R. L. and Griffith, T. S. 2006.
Histone deacetylase inhibitors modulate the sensitivity of tumor necrosis factor-related apoptosis-inducing ligand-re- sistant bladder tumor cells. Cancer Res. 66, 499-507.
9. Fulda, S. 2012. Histone deacetylase (HDAC) inhibitors and regulation of TRAIL-induced apoptosis. Exp. Cell Res. 318, 1208-1212.
10. Ghobrial, I. M., Witzig, T. E. and Adjei, A. A. 2005.
Targeting apoptosis pathways in cancer therapy. CA Cancer J. Clin. 55, 178-194.
11. Giménez-Bonafé, P., Tortosa, A. and Pérez-Tomás, R. 2009.
Overcoming drug resistance by enhancing apoptosis of tu- mor cells. Curr. Cancer Drug Targets 9, 320-340.
12. Han, M. H., Park, C., Kwon, T. K., Kim, G. Y., Kim, W.
J., Hong, S. H., Yoo, Y. H. and Choi, Y. H. 2015. The Histone Deacetylase Inhibitor Trichostatin A Sensitizes Human Renal Carcinoma Cells to TRAIL-Induced Apoptosis through Down-Regulation of c-FLIPL. Biomol. Ther. (Seoul) 23, 31-38.
13. Han, S. I., Kim, Y. S. and Kim, T. H. 2008. Role of apoptotic and necrotic cell death under physiologic conditions. BMB Rep. 41, 1-10.
14. Holcik, M., Gibson, H. and Korneluk, R. G. 2001. XIAP:
apoptotic brake and promising therapeutic targe. Apoptosis 6, 253-261.
15. Hussein, M. R., Haemel, A. K. and Wood, G. S. 2003.
Apoptosis and melanoma: molecular mechanisms. J. Pathol.
199, 275-288.
16. Inoue, H., Shiraki, K., Ohmori, S., Sakai, T., Deguchi, M., Yamanaka, T., Okano, H. and Nakano, T. 2002. Histone de- acetylase inhibitors sensitize human colonic adenocarcino- ma cell lines to TNF-related apoptosis inducing ligand- mediated apoptosis. Int. J. Mol. Med. 9, 521-525.
17. Jin, C. Y., Park, C., Cheong, J., Choi, B. T., Lee, T. H., Lee, J. D., Lee, W. H., Kim, G. Y., Ryu, C. H. and Choi, Y. H.
2007. Genistein sensitizes TRAIL-resistant human gastric ad- enocarcinoma AGS cells through activation of caspase-3.
Cancer Lett. 257, 56-64.
18. Jin, C. Y., Park, C., Hong, S. H., Han, M. H., Jeong, J. W., Xu, H., Liu, H., Kim, G. Y., Kim, W. J., Yoo, Y. H. and Choi, Y. H. 2013. Synergistic induction of TRAIL-mediated apop- tosis by anisomycin in human hepatoma cells via the BH3-only protein Bid and c-Jun/AP-1 signaling pathway.
Biomed. Pharmacother. 67, 321-328.
19. Jin, C. Y., Park, C., Hwang, H. J., Kim, G. Y., Choi, B. T., Kim, W. J. and Choi, Y. H. 2011. Naringenin up-regulates the expression of death receptor 5 and enhances TRAIL-in- duced apoptosis in human lung cancer A549 cells. Mol.
Nutr. Food Res. 55, 300-309.
20. Jin, Z. and El-Deiry, W. S. 2005. Overview of cell death sig- naling pathways. Cancer Biol. Ther. 4, 139-163.
21. Kantari, C. and Walczak, H. 2011. Caspase-8 and bid: caught in the act between death receptors and mitochondria.
Biochim. Biophys. Acta 1813, 558-563.
22. Kelly, M. M., Hoel, B. D. and Voelkel-Johnson, C. 2002.
Doxorubicin pretreatment sensitizes prostate cancer cell lines to TRAIL induced apoptosis which correlates with the loss of c-FLIP expression. Cancer Biol. Ther. 1, 520-527.
23. Kim, E. Y., Ryu, J. H. and Kim, A. K. 2013. CAPE promotes TRAIL-induced apoptosis through the upregulation of TRAIL receptors via activation of p38 and suppression of JNK in SK-Hep1 hepatocellular carcinoma cells. Int. J. Oncol.
43, 1291-1300.
24. Kim, Y. H., Park, J. W., Lee, J. Y. and Kwon, T. K. 2004.
Sodium butyrate sensitizes TRAIL-mediated apoptosis by induction of transcription from the DR5 gene promoter through Sp1 sites in colon cancer cells. Carcinogenesis 25, 1813-1820.
25. Kouraklis, G. and Theocharis, S. 2002. Histone deacetylase inhibitors and anticancer therapy. Curr. Med. Chem. Antican- cer Agents 2, 477-484.
26. Lazebnik, Y. A., Kaufmann, S. H., Desnoyers, S., Poirier, G.
G. and Earnshaw, W. C. 1994. Cleavage of poly(ADP-ribose) polymerase by a proteinase with properties like ICE. Nature 371, 346-347.
초록:인체 방광암세포에서 histone deacetylase 억제제인 sodium butyrate이 TRAIL에 의한 apoptosis 유도에 미치는 영향
한민호
1․최영현
2,3*
(1국립해양생물자원관 천연물연구팀, 2동의대학교 한의과대학 생화학교실, 3동의대학교 항노화연구소)
TRAIL은 정상세포에서는 세포독성을 나타내지 않는 반면, 암세포에서는 사멸을 유도하므로 항암제로 각광받 고 있지만 많은 암세포에서 TRAIL에 저항성을 가지고 있는 것으로 알려져 있으므로 이를 극복해야하는 큰 어려 움이 남아있다. 본 연구에서는 TRAIL에 저항성을 가지는 인간 방광암 세포주인 5637 세포를 이용하여 histone deacetylase 억제제인 sodium butyrate (SB)와 TRAIL을 혼합처리하였을 경우 유발되는 세포사멸 효과와 이와 관 련된 분자생물학적 메카니즘을 연구하였다. 세포독성이 없는 조건의 TRAIL과 SB를 혼합처리 하였을 경우 SB 단 독처리군 보다 세포사멸이 현저하게 증가하는 것으로 확인되었다. TRAIL과 SB의 혼합처리는 caspases (caspase- 3, -8 and -9)의 활성화 및 PARP의 단편화를 유발하였다. 하지만 caspase 억제제에 의하여 TRAIL과 SB의 혼합처 리에 의하여 유발되는 apoptosis가 현저하게 억제되는 것으로 나타났다. 또한 TRAIL과 SB의 혼합처리는 세포표 면에 존재하는 DR5의 발현 증가 및 c-FLIP의 발현 감소를 유발하였으며, pro-apoptotic protein인 Bax와 세포질 cytochrome c의 발현 증가 및 anti- apoptotic protein인 Bcl-xL의 발현감소와 함께 tBid의 형성을 유발하였다. 이 는 SB와 TRAIL의 혼합처리가 안전하고 선택적으로 TRAIL에 저항성을 가지는 방광암 세포에서 치료하는데 효과 적인 전략임을 제시하는 결과이다.
27. Lee, H. H., Jeong, J. W., Lee, J. H., Kim, G. Y., Cheong, J., Jeong, Y. K., Yoo, Y. H. and Choi, Y. H. 2013. Cordycepin increases sensitivity of Hep3B human hepatocellular carci- noma cells to TRAIL-mediated apoptosis by inactivating the JNK signaling pathway. Oncol. Rep. 30, 1257-1264.
28. Mérino, D., Lalaoui, N., Morizot, A., Solary, E. and Micheau, O. 2007. TRAIL in cancer therapy: present and future challenges. Expert. Opin. Ther. Targets 11, 1299-1314.
29. Monneret, C. 2005. Histone deacetylase inhibitors. Eur. J.
Med. Chem. 40, 1-13.
30. Mühlethaler-Mottet, A., Flahaut, M., Bourloud, K. B., Auderset, K., Meier, R., Joseph, J. M. and Gross, N. 2006.
Histone deacetylase inhibitors strongly sensitise neuro- blastoma cells to TRAIL-induced apoptosis by a caspases- dependent increase of the pro- to anti-apoptotic proteins ratio. BMC Cancer 6, 214.
31. Park, C., Jin, C. Y., Hwang, H. J., Kim, G. Y., Jung, J. H., Kim, W. J., Yoo, Y. H. and Choi, Y. H. 2012. J7, a methyl jasmonate derivative, enhances TRAIL-mediated apoptosis through up-regulation of reactive oxygen species generation in human hepatoma HepG2 cells. Toxicol. In Vitro 26, 86-93.
32. Rae, C., Langa, S., Tucker, S. J. and MacEwan, D. J. 2007.
Elevated NF-kappaB responses and FLIP levels in leukemic but not normal lymphocytes: reduction by salicylate allows TNF-induced apoptosis. Proc. Natl. Acad. Sci. USA 104, 12790-12795.
33. Rosato, R. R., Almenara, J. A., Dai, Y. and Grant, S. 2003.
Simultaneous activation of the intrinsic and extrinsic path- ways by histone deacetylase (HDAC) inhibitors and tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) synergistically induces mitochondrial damage and apopto- sis in human leukemia cells. Mol. Cancer Ther. 2, 1273-1284.
34. Safa, A. R. and Pollok, K. E. 2011. Targeting the Anti- Apoptotic Protein c-FLIP for Cancer Therapy. Cancers (Basel) 3, 1639-1671.
35. Seo, O. W., Kim, J. H., Lee, K. S., Lee, K. S., Kim, J. H., Won, M. H., Ha, K. S., Kwon, Y. G. and Kim, Y. M. 2012.
Kurarinone promotes TRAIL-induced apoptosis by inhibit- ing NF-κB-dependent cFLIP expression in HeLa cells. Exp.
Mol. Med. 44, 653-664.
36. Sonnemann, J., Gänge, J., Kumar, K. S., Müller, C., Bader, P. and Beck, J. F. 2005. Histone deacetylase inhibitors inter- act synergistically with tumor necrosis factor-related apop- tosis-inducing ligand (TRAIL) to induce apoptosis in carci- noma cell lines. Invest. New Drugs 23, 99-109.
37. Srivastava, R. K. 2001. TRAIL/Apo-2L: mechanisms and clinical applications in cancer. Neoplasia 3, 535-546.
38. Teraishi, F., Kagawa, S., Watanabe, T., Tango, Y., Kawashima, T., Umeoka, T., Nisizaki, M., Tanaka, N. and Fujiwara, T.
2005. ZD1839 (Gefitinib, 'Iressa'), an epidermal growth fac- tor receptor-tyrosine kinase inhibitor, enhances the anti-can- cer effects of TRAIL in human esophageal squamous cell carcinoma. FEBS Lett. 579, 4069-4075.