www.earticle.net
한우 Germline에서 Octamer-Binding Transcription Factor 4(Oct-4)의 발현
김 현1 · 최재혁2 · 김성우1 · 양보석1 · 고응규1*
국립축산과학원 가축유전자원시험장1, 국립축산과학원 동물바이오공학과2
Expression of Oct-4 in Korean Native Cattle (Hanwoo) Germline
Hyun Kim1, Jae-Hyuk Choi2, Sung-woo Kim1, Bohsuk Yang1 and Yeoung-Gyu Ko1*
1Animal Genetic Resources Station, National Institute of Animal Science, R.D.A.,
2Animal Biotechnology Division, Korea national livestock institute, R.D.A.
ABSTRACT1)
The Oct-4 (octamer-4), a member of the POU family transcription factor, is expressed in early mouse embryogenesis and in pluripotent embryonic stem (ES) lines. Oct-4 expression is thought to remain confined to the germline after gastrulation in the embryo. Therefore, the study was designed to, study the location of Oct-4 protein in the ovaries, placenta and testis of Korean native cattle (Hanwoo). Expression of Oct-4 mRNA in the ovaries and placenta of bovine was confirmed by RT-PCR and immunohistochemical analysis. Oct-4 was expressed in granulosa, thecal cells irrespective of the shape and size of follicles and endometerium of Korean native cattle (Hanwoo). Expression of Oct-4 was profound in all the tissues of Korean native cattle (Hanwoo) suggestung their role in them. Oct-4 localization and expression could contribute to further developmental studies in Korean native cattle (Hanwoo).
(Key words: Oct-4, Germline, Ovary, Placenta, Testis)
Ι .
서론Oct-4(octamer-binding transcription factor 4)는 promoter 혹은 enhancer 영역 내에서 octamer를 인식하는 염기서열과 결합에 의해 표적유전자의 전사를 조절하는 전사인자 POU 패밀리의 한 멤버이다(Scholer 등, 1991; Pesce 등, 2001). 전 사인자로서 기능하는데, Pou5f1 유전자에 의해 암호화된 마 우스 등 설치류의 Oct-4 발달중인 마우스 배아에서 다분화 성 세포군의 구축과 유지 그리고 초기계통발달의 조절을 위 해 필수적인 역할을 한다고 보고되었다(Nichols 등, 1998;
Pesce 등, 2001). 할구 분할 단계, 배반포의 내부 세포괴 (inner cell mass, ICM), 착상 후 초기배아의 외피세포 그리 고 다분화능을 지닌 배아줄기세포 등에서 발현 된다고 보고 되었다(Yeom 등, 1991; Palmieri 등, 1994). 이와 같이 Oct-4 는 전사인자로써 배 발생기 전 과정에 걸쳐 생식세포(germ
cell)와 배아 줄기세포(embryonic stem, ES) 에서 발현된다 고 보고되었다(Fire 등, 1998; Niwa 등, 2001). 특히, 분화 과 정 중에서도 Oct-4는 초기 수정란(Scholer 등, 1990)에서 강 한 발현 양상을 보였으며, 내부 세포괴 그리고 영양막, 내배 엽, 외배엽으로 구분되는 배반포 단계에 있어서는 내부 세포 괴 에서 제한적으로 발현 한다고도 보고되었다(Pesce 등, 1998). 이는 내부 세포괴를 구성하고 있는 세포들은 잠재적 으로 여러 세포로 분화 할 수 있는 다분화성을 지닌 배아줄 기세포(embryonic stem, ES)인데 반하여, 영양막 그리고 내 배엽, 외배엽의 세포들은 수정란을 보호하고 그 기능을 유지 하는 조직으로 분화하도록 이미 운명이 결정되어 있는 차이 점이 있다.
최근 보고에 의하면, Oct-4의 발현 분석 결과, 배아 줄기세 포에서 발현이 증가하면 원시 내배엽과 원시 중배엽으로 분 화를 하고, 일정한 수준의 발현양상을 나타낼 때는 자가 재
* Corresponding Author: Yeoung-Gyu Ko, Animal Genetic Resources Station, National Institute of Animal Science, R.D.A., Namwon 590-832, Korea. Tel: +82-63-620-3562, E-mail: [email protected]
[Provider:earticle] Download by IP 118.70.52.165 at Monday, December 20, 2021 8:04 PM
www.earticle.net
생과 다분화성을 계속 유지하며, 발현이 억제될 경우 영양 막 외배엽으로 분화한다는 사실도 밝혀졌다(Chambers 등, 2003). 또한 Cinzia 등은 배 발생과정에 중요한 역할을 하는 Fibroblast growth factor-4(FGF-4)와 같은 다른 여러 유전자 의 octameric 염기 서열에 결합하여 표적 유전자의 전사를 촉진 또는 억제시키는 기능을 한다고 보고 했다(Cinzia 등, 2000). 하지만, Oct-4의 발현이 내부 세포괴에서만 제한적으 로 이루어지는 구체적인 이유에 대해서는 아직 밝혀지지 않 았고, Oct-4 발현의 증가 또는 억제를 조절하는 유전자에 대 한 연구도 미흡한 실정이다. 마우스에서 Oct-4 발현은 다분 화능세포의 확인을 위한 중요한 표지인자로써 생각되어 왔 다. 그리고, 잠재적으로 다른 포유류에서도 비슷한 기능을 할 가능성이 생각된다. 왜냐하면 단백질 sequence와 이를 조절 하는 영역 그리고 발현 형태와 기능을 관리 조절 하는 기능이 마우스, 사람 그리고 소에서 비슷한 양상을 보였다 (van 등, 1999; Nordhoff 등, 2001). 특히, 단백질 발현 레벨 에 있어서는 90.6% 과 81.7% 상동성을 가진다고 보고되었 다 마우스 등과 같은 설치류와 높은 상동성을 지닌 소의 Oct-4 단백질은 명확하게 착상 전 단계에서 다분화능 배자 성 세포에만 제한되는 것은 아니라 배반포의 모든 세포에서 와 같이 초기분화 그리고 분화 후 단계에서 배반포의 ICM 과 TE양쪽에서 존재한다고도 보고되었다(Kirchhof 등, 2000). 지금까지의 연구 성과들을 면밀히 살펴보면, 마우스 등과 같은 설치류 이외의 소 Oct-4에 관한 연구는 대부분이 착상 전 단계의 배아에서 Oct-4 mRNA가 배반포 단계보다 는 낮은 수준이지만 발현 한다고 했다(Vigneault 등, 2004).
또한 trophectodermal differentiation 조절자와 관계가 있는 interferon-g와 같은 TE-특이적인 유전자의 억제자로서 소의 Oct-4가 작용할 가능성에 대해서도 보고되었다(Ezashi 등, 2001).
이에 반하여, 특히, 한우에서는 이러한 Oct-4에 대한 연구 가 활발하게 이루어지지 않고 있으며, 난포가 발육하는 단 계, 배란 직전 단계까지 발육한 난포, 착상에서 중요한 의미 를 가지는 자궁 그리고 정소와 같은 번식 기관에서 한우의 Oct-4 에 대한 기초 자료 확보는 한우 생산성의 증대 및 번 식 기초 연구 기반의 자료로서도 상당한 가치가 있을 것으 로 사료된다. 따라서, 본 연구는 Oct-4가 임신 한우의 난소, 태반 및 수소의 정소 등과 같은 번식생리 조직 등에서 어떠 한 발현 양상을 보이는지를 확인해 보기 위해서 RT-PCR과 면역조직화학염색 방법을 이용하여 그 발현 양상을 검토해 보기로 하였다.
Ⅱ. 재료 및 방법
1. 공시 동물
별도의 표기가 없는 한 본 실험에 사용된 시약은 Sigma 사(St. Louis, MO, USA) 제품을 사용하였고, 실험에 사용된 동물 관리 및 절차는 국립축산과학원 동물복지위원회 (Suwon, Korea)의 승인을 얻었다. 본 연구에는 임신 중이거 나 분만 직후의 한우가 공시되었고, 임신우의 분만은 제왕 절개 수술과 정상 분만개체로 구분하였으며, 이전의 시험방 법과 동일하게 제왕절개 수술은 일반적으로 평균 분만 예정 일에서 하루 전에 실시되었다.
2. 공시 재료의 처리
소의 태반분엽은 조직을 phosphate buffered saline(PBS, pH 7.2)에 두세 번 깨끗이 Washing 해주고, 이러한 궁부로 부터 fetal cotyledons 그리고 maternal caruncles로 구분하 여 분석 직전까지 -80℃에서 보관하였다. 면역조직화학 염색 을 위한 시료는 제왕 절개 수술 후 회수 되어진 직경 평균 5-6 ㎝ 태반을 면역 조직 화학 염색 실험을 위해 가로 1 ㎝, 세로 1 ㎝로 균등 분할하여 10% formalin solution(St.
Louis, Mo, USA)에 고정시켜 상온(23-25℃)에 보관하였다.
또한, 가로 1 ㎝, 세로 1 ㎝로 균등 분할하여 파라핀 블록을 제작 한 후 사용직전 까지 4℃에 보관하였다. 또한 공시 되어 진 난소 및 정소는 사육중인 한우의 시험 도태 시료를 회수하 여 위의 태반과 같은 방법으로 처리 보관하여 공시하였다.
3. RNA 추출 및 RT-PCR
Oct-4 발현의 관찰은 Qiagen RNasy mini kit(Qiagen, Velncia, USA)를 이용해 합성된 total RNA를 사용하였다.
RT-PCR의 실험은 500 ㎍의 total RNA와 hexamer 그리고 Avian Myeloblastosis Virus-Reverse Transcription(AMV-RT) 를 첨가하여 first cDNA를 합성하였다. PCR은 2 ㎕의 cDNA를 주형으로 하여, 0.25 ml of a-Taq(5 units/ml), 5 ml of 10xPCR buffer, 4 ml of dNTPs(2.5 mM), 3.75 ml of 10xEnhancer, PCR을 위한 멸균된 물 33 ml 그리고 Fig. 1과 같이 디자인된 1 ml의 Oct-4 primers(10 pmol)의 Mixture를 사용하였다. primer: forward 5`-GGTTCTCTTTGGAAA GGTGTTCA-3`reverse 5`-ACACTCGGACCACGTCTTTC-3`
를 이용하여 수행되었다. Oct-4 유전자의 증폭과정은 94℃
[Provider:earticle] Download by IP 118.70.52.165 at Monday, December 20, 2021 8:04 PM
www.earticle.net
에서 10분간 예비 변성(pre-denature) 시킨 후, 94℃ 에서 1 분간 변성(denature)하였고, 36℃에서 1분간 가열냉각 (annealing), 72℃ 1분간 확장(extension) 반응을 35회씩 반 복한 다음 72℃에서 10분 동안 확장(extension)시키는 과정 을 마지막으로 완료시켰다. 증폭되어진 생산물은 0.9% 아가 로스 겔에서 전기영동 후 ethidium bromide 염색을 통하여 시각화하였다.
Fig. 1. Ideogram of bovine Oct-4 cDNA for RT-PCR 4. 면역조직 염색법(immunohistochemistry)
10% formalin에 고정 되어진 태반(평균 임신기간 283±3 일)분만, 난소, 정소 조직을 5 ㎛의 두께로 자르기 위하여 파 라핀을 블록화 한 후, 조직 내 핵을 보기 위한 hematoxylin 염색과 세포질을 eosin으로 염색해 immunostaining 결과와 비교 하였다. 5 ㎛ 두께로 잘리진 조직은 gelatin 코팅이 된 슬라이드 글라스에 잘 펴서 붙인 후 슬라이드 글라스를 PBS 에 세척 후 0.1% triton(St. Louis, Mo, USA)/PBS(St. Louis, Mo, USA)에 30분 간 처리하여 세포막을 확장 한 후 5%
BSA(bovine serum albumin, SIGMA, USA)/0.1% triton(St.
Louis, Mo, USA)/PBS에 2시간 동안 처리하여 면역 조직 화 학 염색 시 잔상을 제거하기 위한 blocking을 실시하였다.
blocking 처리가 끝난 후 조직이 흡착된 슬라이드를 습윤 상자에 넣고 유성 DACO(St. Louis, Mo, USA) 펜으로 원을 조직 주위에 그려준 뒤, 그 원안에 0.1% triton/PBS로 1:100 비율로 희석된 mouse monoclonal IgG-Oct4(Santa Cruz, C-10, USA)를 처리하여 4℃에서 약 16시간 동안 배양 primary antibody 처리를 하였다. Primary antibody에 반응 하는 secondary antibody는 FITC(florescein isothiocyanate, Santa Cruz, USA)와 TRITC(texas red isothiocyanate, Santa Cruz, USA)로 형광 염색 하였으며, 모든 염색 과정의 처리 후 gel mount 용액으로 커버 글라스를 덮고 공 초점 현미경 (confocal microscope/Olympus, Tokyo, Japan) 으로 200배 또는 400배로 확대 관찰 하였다.
Ⅲ. 결과 및 고찰
1. 한우의 태반, 난소 및 정소에서의 Oct-4 mRNA 발현
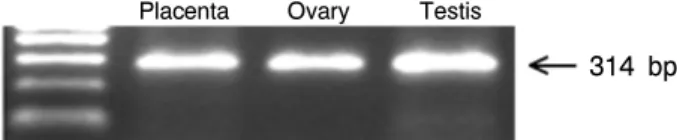
한우의 난소, 태반 및 정소 조직에서 추출된 total RNA을 이용하여 RT-PCR을 수행하였으며, RT-PCR에 의해 얻어진 증폭 생산물을 전기영동으로 확인한 결과 3가지 조직에서 모두 314 bp 크기의 절편이 증폭 되었다(Fig. 2). 이 결과로 부터, 한우의 번식생리 기관인 난소, 태반 및 정소에서 Oct-4가 발현됨을 확인 할 수 있었다.
Placenta Ovary Testis
314 bp
Fig. 2. Electrophoresis photograph was showed results of RT-PCR for Oct-4 in extracted total RNA from specific tissues (placenta, ovary and testis) of Hanwoo.
2. 한우의 난소와 태반에서의 Oct-4 단백질 발현 양상
한우 난소, 태반 조직 내 Oct-4 단백질의 발현 분포 양상 을 면역조직화학 염색법을 이용해 확인을 했다. 먼저, Oct-4 시그널이 조금 약하지만, 원시 난포에서 Oct-4가 발현하는 양상을 확인했다. 그리고 난포성숙이 진행 중인 2차 난포와 배란직전의 그라피안(graffian) 난포에서도 Oct-4 단백질이 발현하고 있음을 확인했다. 일반적으로, Oct-4의 기능을 밝 히기 위한 연구에서 흰쥐, 마우스 그리고 햄스터 등과 같은 설치류가 실험 재료로서 이용되고, 그 결과들이 보고되어 왔지만, 본 연구 결과로 부터, 난포의 발육 단계에 상관없이 발육중인 난포에서 Oct-4 단백질이 과립막 세포 그리고 내, 외협 막 세포 에서 발현하고 있음을 처음으로 한우에서 확 인을 했다는 것에는 큰 의미가 있을 것으로 생각된다. 또한 Oct-4의 발현 변화가 난포에서 난자의 발달에 미치는 영향 에 대한 근거가 될 것으로 사료된다. 한편, 태반에서는 주로 영양아 세포층에서 Oct-4 단백질이 강하게 발현되는 양상을 보이는 것으로부터 난포 발육, 배란, 수정 후 착상 그리고 분만시기의 태반에서도 발현하는 것이 관찰되었다. 특히, 수 정란분화 후 영양막 외배엽으로 발달한 부분은 Oct-4의 발 현 양이 적어짐에 따라 수정란 착상 후 태반의 영양아 세포 층(trophoblast)으로 발달하여 태반의 영양아 세포층으로서 의 역할로 모체와 태아간의 영양분에 대한 통로 및 저장 역
[Provider:earticle] Download by IP 118.70.52.165 at Monday, December 20, 2021 8:04 PM
www.earticle.net
할을 한다(Chambers 등, 2003). 난소, 태반 및 정소에서 Oct-4 mRNA의 발현으로 밝혀진 한우의 Oct-4에 대한 연구 는 단백질로서 Oct-4의 발현이 난소와 태반 정소에서 모두 이루어지고 있음을 말해주고 있다.
본 연구를 통해서 처음으로 한우의 생식, 번식 기관인 난 소, 태반 및 정소에서 Oct-4 mRNA가 존재하고 있음을 확 인했고, 더 나아가 단백질 형태로서도 발육중인 난포의 과 립막세포 뿐만 아니라 협막세포 그리고 태반의 영양아 세포 에서도 강하게 발현하는 것을 확인했다. 특히, Oct-4가 그 기능이 조직의 분화 및 발달을 조절하는 인자의 하나로만 밝혀져 있지만(Yeom 등, 1996), 분만 후 태반에서 발현되는 양상을 기초로 하여 Oct-4 단백질에 결합하는 단백질 규명 과 그 기능의 해석을 통하여 다른 전사 조절인자로서의 역 할에 대한 연구를 할 수 있을 것이라 사료되며, 흰쥐, 마우 스 그리고 햄스터 등과 같은 설치류 등에서 Oct-4는 전사인 자로서 수정 후 배반포기 단계에서부터 증가하기 시작하고, 발생 초기 단계(blastocyst formation)에서의 배아 줄기 세포 분화를 조절하는 전능성으로 분화하는 조절 인자로서 작용 한다고 보고되고 있다(Boiani 등, 2002; Ryan 등, 1997). 더 욱이, Oct-4의 발현은 흰쥐, 마우스 그리고 햄스터 에서도 보고된 바와 같이 임신 기간 중 특정한 시기에 발현됨으로 써(up and down regulation) 한우에서도 이와 비슷한 발현 양상 보이고 있는 것으로 유추할 수 있고, 유전 정보의 재편 성에 전사인자로써 Oct-4가 중추적인 역할을 하는 것으로 미루어 볼 때 향후, 유전자 형질전환 동물이나 복제 동물의
연구에서도 중요한 열쇠가 될 것이라 사료된다. 더 나아가 지금까지 미흡했던 한우에 있어 Oct-4에 대한 연구가 활발 하게 이루어 질 수 있는 하나의 동기부여를 함과 동시에 난 포가 발육하는 단계, 배란 직전 단계까지 발육한 난포, 착상 에서 중요한 의미를 가지는 자궁, 태반 그리고 정소와 같은 번식 기관에서 한우의 Oct-4 에 대한 기초 자료 확보는 한 우 생산성의 증대 및 전반적인 번식 기초 연구 기반의 자료 로서도 상당한 가치가 있을 것으로 사료된다.
Ⅳ. 요약
본 연구는 신체 조직 발달에 관계성이 깊은 전사 인자 POU(Pit-Oct-Unc) 중의 하나인 Oct-4가 수정란의 단계 이후 한우의 태반, 난소 및 정소에서 어떤 양상으로 발현하는지 에 대한 기초자료 확보를 위한 연구를 RT-PCR과 면역 조직 화학염색 실험 방법을 이용하여 수행하였고 mRNA의 발현 여부와 이에 따른 조직에서의 Oct-4 단백질의 발현 부위 (localization)를 확인 하였는데, 그 결과를 요약하면 다음과 같다. 한우의 태반, 난소 및 정소 조직에서 추출된 total RNA을 이용하여 RT-PCR을 수행하였으며, RT-PCR에 의해 얻어진 증폭 산물을 전기영동으로 확인 한 결과. 한우의 3가 지 조직에서 모두 314 bp 크기의 mRNA가 증폭되었으며, 이러한 결과로 볼 때, 한우의 태반, 난소 및 정소에서 모두 Oct-4 mRNA가 발현됨을 알 수 있었다. 난소에서의 Oct-4
Fig. 3. Immunohistochemical analyses of primordial, secondary, graffian ovarian follicle and placenta of the bovine.
A (a), B (b), C (c) and D(d) show primordial, secondary, graffian ovarian follicle and placenta, respectively. Cross-section through the primordial, secondary, graafian ovarian follicle and placenta (a, b, c and d). Note that ovarian follicle and placenta which are positive for Oct-4 (arrowheads in A (a), B (b), C (c) and D (d), respectively. Scale bars=400 mm in B (b), C (c), D (d), and 200 mm in A(a). (d) Cross-section through the bovine placenta (hematoxylin and eosin stain).
[Provider:earticle] Download by IP 118.70.52.165 at Monday, December 20, 2021 8:04 PM
www.earticle.net
의 발현 양상은 원시난포 단계에서는 난포의 모든 부분에서 관찰되었고, 2차 난포의 단계에서는 과립막 세포와 과립막 에서 발현되었고, 배란 직후의 난포에서는 난포의 협막에서 발현되는 것이 관찰되었다. 이는 난포의 성숙에 따라 Oct-4 의 다른 양상의 발현이 있는 것으로 사료된다. 또, 태반에서 는 수정란의 영양막 외배엽에서 유래 되어진 영양아 세포층 에서 전반적으로 발현되는 것이 관찰되었다. 본 연구의 결 과에서와 같이 태반에서의 Oct-4 발현은 태아의 성장과 발 육에 관여하는 유전자에 결합하여 그 활성을 보이는 것으로 생각되어진다.
사사
본 연구는 2011년도 농촌진흥청 국립축산과학원 박사 후 연수과정 지원 사업에 의해 이루어진 것임.
Ⅴ. 인용문헌
1. Alexander, B., Catherine, E., Ovitt and Scholer, H. R., 1998. Oct-4: more than just POU powerful maker of the mammalian germline. Apmis. 106:114-126.
2. Andrea, R. P., Erika, M., Margit, R., Gerhard B. and Markus, H. 2003. Oct-4 expressing cells in human amniotic fluid: a new source for stem cell research.
Hum. Reprod. 18(7):1489-1493.
3. Attila, R., Alexey T., Sholer, H. R., and Matthias W.
2002. Differntial activity by DNA-induced quartnary structures of POU transcription factor. J. Phamacol.
64:979-984.
4. Boiani, M. and Scholer, H. R., 2002. Oct-4 distribution and level in mouse clones: consequences for pluripotency. Genes Dev. 16:1209-1219.
5. Chapman, D. L. and Wolgemuth, D. J., 1994.
Expression of proliferating cell nuclear antigen in the mouse germ line and surrounding somatic cells suggests both proliferation-dependent and- independent modes of function. J. Dev. Bio. 38:491-497.
6. Chambers, I., Colby, D., Robertson, M., Nichols, J., Lee, S., Tweedie, S. and Smith. 2003. A. Functional expression cloning of Nanog, a pluripotency sustaining
factor in embryonic stem cells. Cell. 113(5):643-655.
7. Cinzia B., Massimo F., Scholer, H. R. and Maurizio P.
2000. Phage diaplay screening revels an asociation between germline-specific transcrition factor Oct-4 and multiple cellular proteins. J. Mol. Biol. 304:529-540.
8. Eijk, M. J. T., Rooijen, M. A., Modina, S., Scesi, L., Folkers, G., Tol, H. T. A., Bevers, M. M., Fisher, S. R., Lewin, H. A. and Gandolfi, F. 1999. Molecular cloning, genetic mapping and developmental expression of bovine POU5f1. Biol. Reprod. 60:1093-1103.
9. Elbashir, S. M., Harborth, J., Lendeckel, W., Yalcin, A., Weber, K. and Tuschl, T., 2001. Duplexes of 21-nucleotide RNAs mediate RNA interference in cultured mammalian cells. Nature. 411:494-498.
10. Fire, A., Xu, S., Montgomery, S. A., Kostas, S. E., Driver and Mello, C. C., 1998. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature. 391:806-811.
11. Guang, J., P., Zeng, Y., CHANGI, Scholer, H. R. and Duanqing, P. 2002. Stem cell pluripotency and transcription factor Oct-4. Cell. 12(5-6):321-329.
12. Herr, W. and Cleary, M. A. 1995. The POU domain:
versatility intranscriptional regulation by a flexible two-in-one DNA-binding domain. Genes Dev. 9:1679-1693.
13. Kirchhof N, Carnwath J. W, Lemme E, Anastassiadis K, Scholer H, Niemann H. Expression pattern of Oct-4 in preimplantation embryos of different species. Bio.
Reprod. 2000. 63:1698-1705.
14. Laemmil, U. K., 1970. Cleavage of structural protein during the assembly of the head of bacteriophage T4.
Nature. 277:680-685.
15. Lyon, M. and Searle, A. G. 1989. Genetic Variants and Strains of the Laboratory mouse. 2nd. ed. Oxford University.
16. Lee, S. B. and Elbsteban, M. 1994. The Interferon- induced Double-Stranded RNA-Activated Protein Kinase Induces Apoptosis. J. Virol. 199:491-496.
17. Niwa, H. 2001. Molecular mecanism to maintain stem cell renewal of ES cells. Cell Struct. Funct. 26:137-148.
(투고일: 2011.10.06. 수정일: 2011.11.24. 판정일: 2011.11.25.)
[Provider:earticle] Download by IP 118.70.52.165 at Monday, December 20, 2021 8:04 PM